

Individual Genomic Loci and mRNA Levels of Immune Biomarkers Associated with Pneumonia Susceptibility in Baladi Goats


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Research Subjects
2.3. DNA Isolation and Amplification
2.4. Sequencing DNA and Detecting Polymorphism
2.5. RNA Isolation and Immune Gene Quantitation
2.6. Statistical Analysis
3. Results
3.1. Clinical Findings
3.2. Polymorphisms of Immune Markers
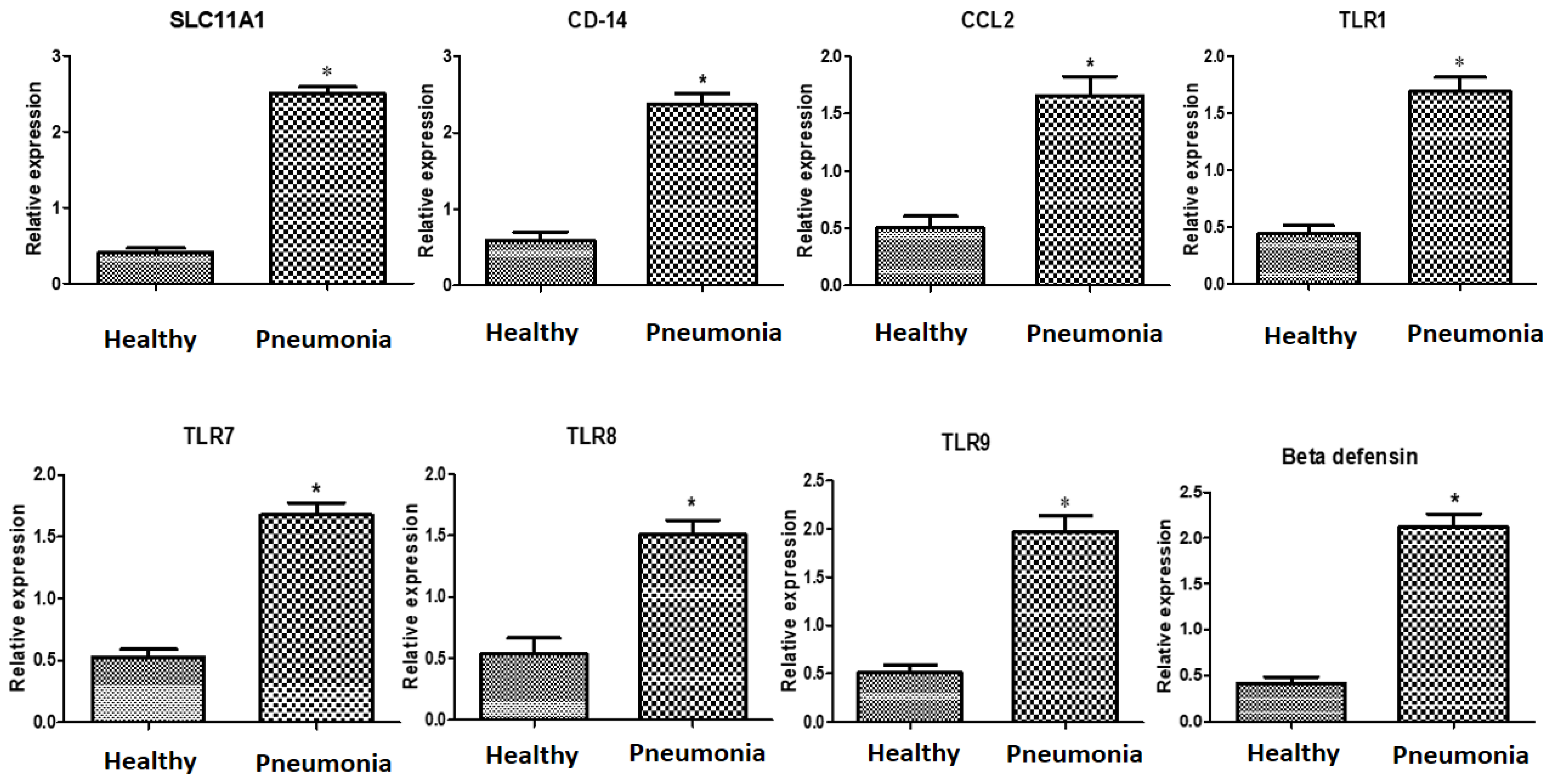
3.3. Immune Markers’ Patterns of Gene Expressions
4. Discussion
4.1. Individual Genomic Loci of Immune Markers Associated with Pneumonia Susceptibility
4.2. Transcript Levels of Immune Markers Associated with Pneumonia Susceptibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skapetas, B.; Bampidis, V. Goat production in the world: Present situation and trends. Livest. Res. Rural. Dev. 2016, 28, 200. [Google Scholar]
- Gall, C. Goat Breeds Around the World; CTA; Margraf/FAO: Weikersheim, Germany, 1996; p. 186. [Google Scholar]
- Zeder, M.A.; Hesse, B. The initial domestication of goats (Capra hircus) in the Zagros Mountains 10,000 years ago. Science 2000, 287, 2254–2257. [Google Scholar] [CrossRef] [PubMed]
- MacHugh, D.E.; Bradley, D.G. Livestock genetic origins: Goats buck the trend. Proc. Natl. Acad. Sci. USA 2001, 98, 5382–5384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, M. Review of modern strategies to enhance livestock genetic performance: From molecular markers to next-generation sequencing technologies in goats. J. Food Agric. Environ. 2014, 12, 752–761. [Google Scholar]
- Haftu, A.B.B.; Gebrehiwot, T. Study on prevalence of gastrointestinal nematodes and coccidian parasites affecting cattle in West Arsi zone, Ormia Regional State, Ethiopia. J. Biol. Agric. Healthc. 2014, 4, 32–38. [Google Scholar]
- Ahmed, J.; Ararsa, D.; Dareje, R.; Dinaol, B.; Roba, J. Gastrointestinal nematode parasites of small ruminants and anthelmintics efficacy test in sheep of Haramaya District, Eastern Ethiopia. Anim. Vet. Sci. 2017, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Crespo, H.; Bertolotti, L.; Proffiti, M.; Cascio, P.; Cerruti, F.; Acutis, P.L.; de Andrés, D.; Reina, R.; Rosati, S. Low proviral small ruminant lentivirus load as biomarker of natural restriction in goats. Vet. Microbiol. 2016, 192, 152–162. [Google Scholar] [CrossRef]
- Ayling, R.D.; Nicholas, R.A.J. Mycoplasma Respiratory Infections. In Diseases of Sheep, 4th ed.; Aitken, I.D., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 231–235. [Google Scholar]
- Bottinelli, M.; Schnee, C.; Lepri, E.; Stefanetti, V.; Filippini, G.; Gobbi, M.; Martina Sebastianelli, M.; Antenucci, P.; Rampacci, E.; Coletti, M.; et al. Investigation on mycoplasma populations in pneumonic dairy lamb lungs using a DNA microarray assay. Small Rumin. Res. 2017, 147, 96–100. [Google Scholar] [CrossRef]
- Medina, I.; Carbonell, J.; Pulido, L.; Madeira, S.; Goetz, S.; Conesa, A.; Tárraga, J.; Pascual-Montano, A.; Nogales-Cadenas, R.; Santoyo-Lopez, J.; et al. Babelomics: An integrative platform for the analysis of transcriptomics, proteomics and genomic data with advanced functional profiling. Nucleic Acids Res. 2010, 38, W210–W213. [Google Scholar] [CrossRef]
- Legastelois, I.; Cottin, V.; Mornex, J.F.; Cordier, G. Alveolar macrophages from sheep naturally infected by visna-maedi virus contribute to IL-8 production in the lung. Vet. Immunol. Immunopathol. 1997, 59, 131–139. [Google Scholar] [CrossRef]
- Song, Q.Y.; Zhang, Y.J.; Liu, Y.Q.; Li, T.Q. Antimicrobial activities of eighteen antimicrobials against Mycoplasma ovipneumoniae and Mycoplasma mycoides isolates in vitro. Pro. Vet. Med. 2011, 32, 14–18. [Google Scholar]
- Abendaño, N.; Tyukalova, L.; Barandika, J.F.; Balseiro, A.; Sevilla, I.A.; Garrido, J.M.; Juste, R.A.; Alonso-Hearn, M. Mycobacterium avium subsp. paratuberculosis isolates induce in vitro granuloma formation and show successful survival phenotype, common anti-inflammatory and antiapoptotic responses within ovine macrophages regardless of genotype or host of origin. PLoS ONE 2014, 9, e104238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Salman, D.; Marghani, B.H.; Gadalla, H.E.; Sayed-Ahmed, M.Z. Immunomodulatory Effect of Vitamin C on Proinflammatory Cytokines Production in Ossimi Lambs (Ovis aries) with Pneumonic Pasteurellosis. Animals 2021, 11, 3374. [Google Scholar] [CrossRef] [PubMed]
- Fernández, S.; Galapero, J.; Rey, J.; Pérez, C.J.; Ramos, A. Cytokines Study in Lungs Infected with Pasteurellaceae and Mycoplasma spp. from Fattening Lambs. J. Med. Microbiol. Immunol. Res. 2018, 2, 1. [Google Scholar]
- Stear, M.J.; Bishop, S.C.; Mallard, B.A.; Raadsma, H.W. The sustainability, feasibility and desirability of breeding livestock for disease resistance. Res. Vet. Sci. 2001, 71, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bishop, S.C.; Morris, C.A. Genetics of disease resistance in sheep and goats. Small Rumin. Res. 2007, 70, 48–59. [Google Scholar] [CrossRef]
- Berry, D.P.; Bermingham, M.L.; Good, M.; More, S.J. Genetics of animal health and disease in cattle. Ir. Vet. J. 2011, 64, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.N.; Mousel, M.R.; Reynolds, J.O.; Lewis, G.S.; Herrmann-Hoesing, L.M. Common promoter deletion is associated with 3.9-fold differential transcription of ovine CCR5 and reduced proviral level of ovine progressive pneumonia virus. Anim. Genet. 2009, 40, 583–589. [Google Scholar] [CrossRef]
- Woodall, C.J.; Maclaren, L.J.; Watt, N.J. Differential levels of mRNAs for cytokines, the interleukin-2 receptor and class II DR/DQ genes in ovine interstitial pneumonia induced by Maedi Visna virus infection. Vet. Pathol. 1997, 34, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Ibeagha-Awemu, E.M.; Kgwatalala, P.; Ibeagha, A.E.; Zhao, X. A critical analysis of disease-associated DNA polymorphisms in the genes of cattle, goat, sheep, and pig. Mamm. Genome 2008, 19, 226–245. [Google Scholar] [CrossRef] [Green Version]
- Cortes, O.; Cañon, J.; Gama, L.T. Applications of Microsatellites and Single Nucleotide Polymorphisms for the Genetic Characterization of Cattle and Small Ruminants: An Overview. Ruminants 2022, 2, 456–470. [Google Scholar] [CrossRef]
- Marwaha, S.; Knowles, J.W.; Ashley, E.A. A guide for the diagnosis of rare and undiagnosed disease: Beyond the exome. Genome Med. 2022, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, X.; Li, Q.; Wan, K.; Gao, R.; Han, G.; Li, C.; Xu, M.; Jia, B.; Shen, X. MHC-DRB1 exon 2 polymorphism and its association with mycoplasma ovipneumonia resistance or susceptibility genotypes in sheep. J. Genet. 2020, 99, 22. [Google Scholar] [CrossRef] [PubMed]
- Leymaster, K.A.; Chitko-McKown, C.G.; Clawson, M.L.; Harhay, G.P.; Heaton, M.P. Effects of TMEM154 haplotypes 1 and 3 on susceptibility to ovine progressive pneumonia virus following natural exposure in sheep. J. Anim Sci. 2013, 91, 5114–5121. [Google Scholar] [CrossRef]
- Pugh, D.G.; Baird, A.N. Sheep & Goat Medicine-E-Book; Elsevier/Saunders: Maryland Heights, Mo, USA, 2012. [Google Scholar]
- Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W.; Constable, P.D. Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 10th ed.; Elsevier/Saunders: London, UK, 2006; pp. 966–994. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=1679700 (accessed on 2 January 2023).
- Boom, R.; Sol, C.J.; Salimans, M.M.; Jansen, C.L.; Wertheim-van Dillen, P.M.; Noordaa, J.V.D. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [Green Version]
- Boesenberg-Smith, K.A.; Pessarakli, M.M.; Wolk, D.M. Assessment of DNA Yield and Purity: An Overlooked Detail of PCR Troubleshooting. Clin. Microbiol. Newsl. 2012, 34, 1–6. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−∆∆CT. Methods. 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Triantaphyllopoulos, K.A.; Ikonomopoulos, I.; Bannister, A.J. Epigenetics and inheritance of phenotype variation in livestock. Epigenetics Chromatin. 2016, 9, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, S.C.; Woolliams, J.A. Genomics and disease resistance studies in livestock. Livest. Sci. 2014, 166, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zonaed Siddiki, A.M.A.M.; Miah, G.; Islam, M.S.; Kumkum, M.; Rumi, M.H.; Baten, A.; Hossain, M.A. Goat Genomic Resources: The search for genes associated with its economic traits. Int. J. Genom. 2020, 18, 5940205. [Google Scholar] [CrossRef]
- Kirkpatrick, B.W.; Cooke, M.E.; Frie, M.; Sporer, K.R.B.; Lett, B.; Wells, S.J.; Coussens, P.M. Genome-wide association analysis for susceptibility to infection by Mycobacterium avium ssp. paratuberculosis in US Holsteins. J. Dairy Sci. 2022, 105, 4301–4313. [Google Scholar] [CrossRef]
- Cartegni, L.; Chew, S.L.; Krainer, A.R. Listening to silence and understanding nonsense: Exonic mutations that affect splicing. Nat. Rev. Genet. 2002, 3, 285–298. [Google Scholar] [CrossRef]
- Michel, O.; LeVan, T.D.; Stern, D.; Dentener, M.; Thorn, J.; Gnat, D.; Beijer, M.L.; Cochaux, P.; Holt, P.G.; Martinez, F.D.; et al. Systemic responsiveness to lipopolysaccharide and polymorphisms in the toll-like receptor 4 gene in human beings. J. Allergy Clin. Immunol. 2003, 112, 923–929. [Google Scholar] [CrossRef]
- Smirnova, I.; Poltorak, A.; Chan, E.K.; McBride, C.; Beutler, B. Phylogenetic variation and polymorphism at the toll-like receptor 4 locus (TLR-4). Genome Biol. 2000, 1, RESEARCH002. [Google Scholar] [CrossRef] [Green Version]
- Yaman, Y.; Bay, V.; Aymaz, R.; Keleş, M.; Öner, Y.; Teferedegn, E.Y.; Ün, C. A novel 2 bp deletion variant in Ovine-DRB1 gene is associated with increased Visna/maedi susceptibility in Turkish sheep. Sci. Rep. 2021, 11, 14435. [Google Scholar] [CrossRef]
- Salamaikina, S.; Karnaushkina, M.; Korchagin, V.; Litvinova, M.; Mironov, K.; Akimkin, V. TLRs Gene Polymorphisms Associated with Pneumonia before and during COVID-19 Pandemic. Diagnostics 2023, 13, 121. [Google Scholar] [CrossRef]
- Jansen, R.C.; Nap, J.P. Genetical genomics: The added value from segregation. Trends Genet. 2001, 17, 388–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairfax, B.P.; Knight, J.C. Genetics of gene expression in immunity to infection. Curr. Opin. Immunol. 2014, 30, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cloney, R. Complex traits: Integrating gene variation and expression to understand complex traits. Nat. Rev. Genet. 2016, 17, 194. [Google Scholar] [CrossRef] [PubMed]
- Abendaño, N.; Esparza-Baquer, A.; Bernales, I.; Reina, R.; de Andrés, D.; Jugo, B.M. Gene Expression Profiling Reveals New Pathways and Genes Associated with Visna/Maedi Viral Disease. Animals 2021, 11, 1785. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Ganguly, N.; Singh, R.; Deb, S.M.; Kumar, S.; Sharma, A.; Mitra, A. DNA polymorphism in SLC11A1 gene and its association with brucellosis resistance in Indian zebu (Bos indicus) and crossbred (Bos indicus×Bos taurus) Cattle. Asian-Aust. J. Anim. Sci. 2011, 24, 898–904. [Google Scholar] [CrossRef]
- Wright, S.D.; Ramos, R.A.; Tobias, P.S.; Ulevitch, R.J.; Mathison, J.C. CD14, a receptor for complexes of lipopolysaccharide (LPS) and LPS binding protein. Science 1990, 249, 1431–1433. [Google Scholar] [CrossRef] [PubMed]
- Hailman, E.; Lichenstein, H.S.; Wurfel, M.M.; Miller, D.S.; Johnson, D.A.; Kelley, M.; Busse, L.A.; Zukowski, M.M.; Wright, S.D. Lipopolysaccharide (LPS)-binding protein accelerates the binding of LPS to CD14. J. Exp. Med. 1994, 179, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T. Expression of potential lymphocyte trafficking mediator molecules in the mammary gland. Vet. Res. 2003, 34, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Proudfoot, A.I. Chemokine receptors: Multifaceted therapeutic targets. Nat. Rev. Immunol. 2002, 2, 106–115. [Google Scholar] [CrossRef]
- Botos, I.; Segal, D.M.; Davies, D.R. The structural biology of Toll-like receptors. Structure 2011, 13, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Into, T.; Yasuda, M.; Okusawa, T.; Hamahira, S.; Kuroki, Y.; Eto, A.; Nisizawa, T.; Morita, M.; Shibata, K. Involvement of leucine residues at positions 107, 112, and 115 in a leucine-rich repeat motif of human Toll-like receptor 2 in the recognition of diacylated lipoproteins and lipopeptides and Staphylococcus aureus Peptidoglycans. J. Immunol. 2003, 171, 3675–3683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.N.; Taylor, K.H.; Abbey, C.A.; Gill, C.A.; Womack, J.E. Haplotype variation in bovine Toll-like receptor 4 and computational prediction of a positively selected ligand-binding domain. Proc. Natl. Acad. Sci. USA 2003, 100, 10364–10369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caverly, J.M.; Diamond, G.; Gallup, J.M.; Brogden, K.A.; Dixon, R.A.; Ackermann, M.R. Coordinated Expression of Tracheal Antimicrobial Peptide and Inflammatory-Response Elements in the Lungs of Neonatal Calves with Acute Bacterial Pneumonia. Infect. Immun. 2003, 71, 2950–2955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.J.; McQuillan, C.; Finlay, E.K.; O’Farrelly, C.; Fair, S.; Meade, K.G. Comparative genomic identification and validation of β-defensin genes in the Ovis aries genome. BMC Genom. 2017, 18, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Nguyen, T.; Liu, L.; Shamova, O.; Brogden, K.; Lehrer, R.I. Differential expression of caprine β-defensins in digestive and respiratory tissues. Infect Immun. 1999, 67, 6221–6224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttner, K.M.; Lambeth, M.R.; Burkin, H.R.; Burkin, D.J.; Broad, T.E. Localization and genomic organization of sheep antimicrobial peptide genes. Gene 1998, 206, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wang, Y.; Zhang, Y.; Yang, M.; Lv, J.; Liu, J.; Zhang, Y. TALE nickase-mediated SP110 knockin endows cattle with increased resistance to tuberculosis. Proc. Natl. Acad. Sci. USA 2015, 112, E1530–E1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, D.B.; Nakajima, A.; Gulick, T.; Chiche, J.D.; Orth, D.; de La Monte, S.M.; Bloch, K.D. Sp110 localizes to the PML-Sp100 nuclear body and may function as a nuclear hormone receptor transcriptional coactivator. Mol Cell Biol. 2000, 20, 6138–6146. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Larrañaga, O.; Garrido, J.M.; Iriondo, M.; Manzano, C.; Molina, E.; Montes, I.; Vazquez, P.; Koets, A.P.; Rutten, V.P.; Juste, R.A.; et al. SP110 as a novel susceptibility gene for Mycobacterium avium subspecies paratuberculosis infection in cattle. J. Dairy Sci. 2010, 93, 5950–5958. [Google Scholar] [CrossRef] [Green Version]
- Chabas, D.; Baranzini, S.E.; Mitchell, D.; Bernard, C.C.; Rittling, S.R.; Denhardt, D.T.; Sobel, R.A.; Lock, C.; Karpuj, M.; Pedotti, R.; et al. The influence of the proinflammatory cytokine, osteopontin, on autoimmune demyelinating disease. Science 2001, 294, 1731–1735. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a means to cope with environmental insults: Regulation of inflammation, tissue remodeling, and cell survival. J. Clin. Investig. 2001, 107, 1055–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balakrishnan, A.; Marathe, S.A.; Joglekar, M.; Chakravortty, D. Bactericidal/permeability increasing protein: A multifaceted protein with functions beyond LPS neutralization. Innate Immun. 2013, 19, 339–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levashina, E.A.; Moita, L.F.; Blandin, S.; Vriend, G.; Lagueux, M.; Kafatos, F.C. Conserved role of a complement-like protein in phagocytosis revealed by dsRNA knockout in cultured cells of the mosquito, Anopheles gambiae. Cell 2001, 104, 709–718. [Google Scholar] [CrossRef]
- Banks, R.E.; Evans, S.W.; Van Leuven, F.; Alexander, D.; McMahon, M.J.; Whicher, J.T. Measurement of the ‘fast’ or complexed form of alpha 2-macroglobulin in biological fluids using a sandwich enzyme immunoassay. J. Immunol. Methods 1990, 126, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Bonacci, G.R.; Caceres, L.C.; Sanchez, M.C.; Chiabrando, G.A. Activated alpha(2)-macroglobulin induces cell proliferation and mitogen-activated protein kinase activation by LRP-1 in the J774 macrophage-derived cell line. Arch. Biochem. Biophys. 2007, 460, 100–106. [Google Scholar] [CrossRef]
- Borth, W. Alpha 2-macroglobulin, a multifunctional binding protein with targeting characteristics. FASEB J. 1992, 6, 3345–3353. [Google Scholar] [CrossRef]
- Sommerschild, H.T.; Kirkebøen, K.A. Adenosine and cardioprotection during ischaemia and reperfusion—An overview. Acta Anaesthesiol. Scand. 2000, 44, 1038–1055. [Google Scholar] [CrossRef]
- He, H.R.; Li, Y.J.; He, G.H.; Qiang, H.; Zhai, Y.J.; Ma, M.; Wang, Y.J.; Wang, Y.; Zheng, X.W.; Dong, Y.L.; et al. The Polymorphism in ADORA3 Decreases Transcriptional Activity and Influences the Chronic Heart Failure Risk in the Chinese. Biomed. Res. Int. 2018, 31, 4969385. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Liu, T.; Tu, W.; Li, W.; Dong, G.; Xu, C.; Qin, B.; Liu, K.; Yang, J.; et al. CARD15 Gene Polymorphisms Are Associated with Tuberculosis Susceptibility in Chinese Holstein Cows. PLoS ONE 2015, 10, e0135085. [Google Scholar] [CrossRef]
- Li, C.; Ma, L.; Chen, X. Interferon regulatory factor 3-CL, an isoform of IRF3, antagonizes activity of IRF3. Cell. Mol. Immunol. 2011, 8, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.E.; Noyce, R.S.; Mossman, K.L. Innate cellular response to virus particle entry requires IRF3 but not virus replication. J. Virol. 2004, 78, 1706–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, C.L.; Hsu, H.; Chen, C.; Palmer, M.; McGill, J.; Waters, W.R.; Telfer, J.C. The role of bovine γδ T cells and their WC1 co-receptor in response to bacterial pathogens and promoting vaccine efficacy: A model for cattle and humans. Vet. Immunol. Immunopathol. 2014, 159, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Butty, A.M.; Chud, T.C.S.; Cardoso, D.F.; Lopes, L.S.F.; Miglior, F.; Schenkel, F.S.; Cánovas, A.; Häfliger, I.M.; Drögemüller, C.; Stothard, P.; et al. Genome-wide association study between copy number variants and hoof health traits in Holstein dairy cattle. J. Dairy Sci. 2021, 104, 8050–8061. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.X.; Park, Y.K.; Bae, M.; Lee, J.Y. The Potential Role of an Endotoxin Tolerance-Like Mechanism for the Anti-Inflammatory Effect of Spirulina platensis Organic Extract in Macrophages. J. Med. Food. 2017, 20, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D. Pathogen-dependent induction of cytokines and other soluble inflammatory mediators during intramammary infection of dairy cows. J. Anim Sci. 2009, 87, 10–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslan, O.; Goksu, A.G.; Apaydin, N. The evaluation of oxidative stress in lambs with Pestivirus infection. J. Hellenic Vet. Med. Soc. 2017, 68, 299–306. [Google Scholar] [CrossRef]
- Wei, W.; Feng, W.; Xin, G.; Niu, T.; Yan, X. Enhanced effect of κ-carrageenan on TNBS-induced inflammation in mice. Int. Immunopharmacol. 2016, 39, 218–228. [Google Scholar] [CrossRef]
- Blattman, A.W.; Hulme, D.J.; Kinghom, B.P.; Woolaston, R.R.; Gray, G.D.; Beh, K.J. A search for associations between major histocompatibility complex restriction fragment length polymorphism bands and resistances to Haemonchus contours infection in sheep. Anim. Genet. 1993, 24, 277–282. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; McManus, D.P. Concepts in immunology and diagnosis of hydatid disease. Clin. Microbiol. Rev. 2003, 16, 18–36. [Google Scholar] [CrossRef] [Green Version]
- Torcal, M.; Lozano, L.; Larrad, J.C. Immune response and in vivo production of cytokines in patients with liver hydatidosis. Clin. Exp. Immunol. 1996, 106, 317–322. [Google Scholar] [CrossRef]
- Moroz, A.; Czopowicz, M.; Sobczak-Filipiak, M.; Dolka, I.; Rzewuska, M.; Kizerwetter-Swida, M.; Chrobak-Chmiel, D.; Mickiewicz, M.; Witkowski, L.; Szalu´s-Jordanow, O.; et al. The Prevalence of Histopathological Features of Pneumonia in Goats with Symptomatic Caprine Arthritis-Encephalitis. Pathogens 2022, 11, 629. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | SNPs | Healthy n = 60 | Pneumonic n = 60 | Total n = 120 | Mutation Type | Number and Kind of Amino Acid | Chi Value | p Value |
|---|---|---|---|---|---|---|---|---|
| SLC11A1 | A27T | - | 32 | 32/120 | Synonymous | 27 S | 67.25 | <0.0001 |
| C416T | 25 | - | 25/120 | Non-synonymous | 139 T to I | 52.64 | <0.0001 | |
| CD14 | G457A | 36 | - | 36/120 | Non-synonymous | 153 A to T | 75.66 | <0.0001 |
| A491C | 19 | - | 19/120 | Non-synonymous | 164 Q to F | 39.93 | <0.0001 | |
| CCL2 | A262G | 27 | - | 27/120 | Non-synonymous | 88 R to G | 56.74 | <0.0001 |
| T479C | - | 31 | 31/120 | Non-synonymous | 160 L to S | 65.15 | <0.0001 | |
| TLR1 | A189T | - | 26 | 26/120 | Synonymous | 63 A | 54.64 | <0.0001 |
| G285C | 23 | - | 23/120 | Non-synonymous | 95 Q to H | 48.34 | <0.0001 | |
| G308A | 17 | - | 17/120 | Non-synonymous | 103 S to N | 35.73 | <0.0001 | |
| G360C | 38 | - | 38/120 | Non-synonymous | 102 E to D | 79.86 | <0.0001 | |
| G394T | - | 42 | 42/120 | Non-synonymous | 132 V to F | 88.27 | <0.0001 | |
| TLR7 | C347T | 36 | - | 36/120 | Non-synonymous | 116 A to V | 75.66 | <0.0001 |
| TLR8 | G709A | 46 | - | 46/120 | Non-synonymous | 237 G to S | 96.67 | <0.0001 |
| TLR9 | A174G | - | 28 | 28/120 | Synonymous | 58 G | 58.84 | <0.0001 |
| β defensin | C30T | - | 18 | 18/120 | Synonymous | 10 R | 37.83 | <0.0001 |
| T105A | - | 49 | 49/120 | Non-synonymous | 35 D to E | 102.98 | <0.0001 | |
| T197C | 33 | - | 33/120 | Non-synonymous | 66 V to A | 69.35 | <0.0001 | |
| C225T | - | 24 | 24/120 | Synonymous | 75 T | 50.44 | <0.0001 | |
| SP110 | A62C | 27 | - | 27/120 | Non-synonymous | 21 N to T | 56.74 | <0.0001 |
| C184T | 51 | - | 51/120 | Non-synonymous | 62 R to C | 107.18 | <0.0001 | |
| SPP1 | C857T | - | 36 | 36/120 | Non-synonymous | 286 T to I | 75.66 | <0.0001 |
| BP1 | A75G | 16 | - | 16/120 | Synonymous | 25 T | 33.63 | <0.0001 |
| G257A | 37 | - | 37/120 | Non-synonymous | 86 R to Q | 77.76 | <0.0001 | |
| A456G | 25 | - | 25/120 | Synonymous | 152 S | 52.54 | <0.0001 | |
| A2M | C36G | 28 | - | 28/120 | Synonymous | 12 S | 58.84 | <0.0001 |
| ADORA3 | C387A | - | 39 | 39/120 | Synonymous | 129 I | 81.96 | <0.0001 |
| CARD15 | A73G | - | 24 | 24/120 | Non-synonymous | 25 S to G | 50.44 | <0.0001 |
| IRF3 | T133G | 34 | - | 34/120 | Non-synonymous | 45 F to V | 71.45 | <0.0001 |
| C273G | 23 | - | 23/120 | Synonymous | 91 S | 48.34 | <0.0001 | |
| SCART1 | C42T | - | 47 | 47/120 | Synonymous | 14 A | 98.77 | <0.0001 |
| A126G | 19 | - | 19/120 | Synonymous | 42 S | 39.93 | <0.0001 | |
| A147G | - | 53 | 53/120 | Synonymous | 49 V | 111.38 | <0.0001 | |
| T380C | - | 31 | 31/120 | Non-synonymous | 127 V to A | 65.15 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ateya, A.; Al-Sharif, M.; Abdo, M.; Fericean, L.; Essa, B. Individual Genomic Loci and mRNA Levels of Immune Biomarkers Associated with Pneumonia Susceptibility in Baladi Goats. Vet. Sci. 2023, 10, 185. https://doi.org/10.3390/vetsci10030185
Ateya A, Al-Sharif M, Abdo M, Fericean L, Essa B. Individual Genomic Loci and mRNA Levels of Immune Biomarkers Associated with Pneumonia Susceptibility in Baladi Goats. Veterinary Sciences. 2023; 10(3):185. https://doi.org/10.3390/vetsci10030185
Chicago/Turabian StyleAteya, Ahmed, Mona Al-Sharif, Mohamed Abdo, Liana Fericean, and Bothaina Essa. 2023. "Individual Genomic Loci and mRNA Levels of Immune Biomarkers Associated with Pneumonia Susceptibility in Baladi Goats" Veterinary Sciences 10, no. 3: 185. https://doi.org/10.3390/vetsci10030185