
Impacts of Solid-State Fermented Barley with Fibrolytic Exogenous Enzymes on Feed Utilization, and Antioxidant Status of Broiler Chickens

 , , , ,
, , , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Two-Stage Solid Fermentation of Barley with Exogenous Enzymatic Treatment
2.3. Nutritive Value Evaluation of Fermented Barley
2.3.1. Chemical Analysis and Microbial Assessment
2.3.2. Analysis of Reduced Sugar
2.3.3. Phosphorus Analysis
2.3.4. Analysis of Phytase Enzyme Activity
2.4. Animals and Experimental Design
2.5. Growth Performance and Nutrient Metabolizability Trial Monitoring
2.6. Sampling
2.7. Digesta Viscosity and Duodenal pH
2.8. Digestive Enzymes Estimation
2.9. Serum Biochemical Investigations
2.10. Measurement of Redox State in Jejunum and Breast Muscle
2.11. Quantification of Intestinal Barrier Function and Nutrient Transporter-Associated Genes via RT-qPCR
2.12. Histopathological Examination of Intestine
2.13. Statistical Analysis
3. Results
3.1. Chemical Analysis of Unfermented Barley Grains and Fermented and Enzymatically Treated Barley Grains
3.2. Growth Performance
3.3. Estimating Nutrient Metabolizability and Activities of Pancreatic Digestive Enzymes
3.4. Estimating Immune and Biochemical-Related Parameters
3.5. Antioxidant Functions of FBEs on Intestinal Tissues and Breast Muscle
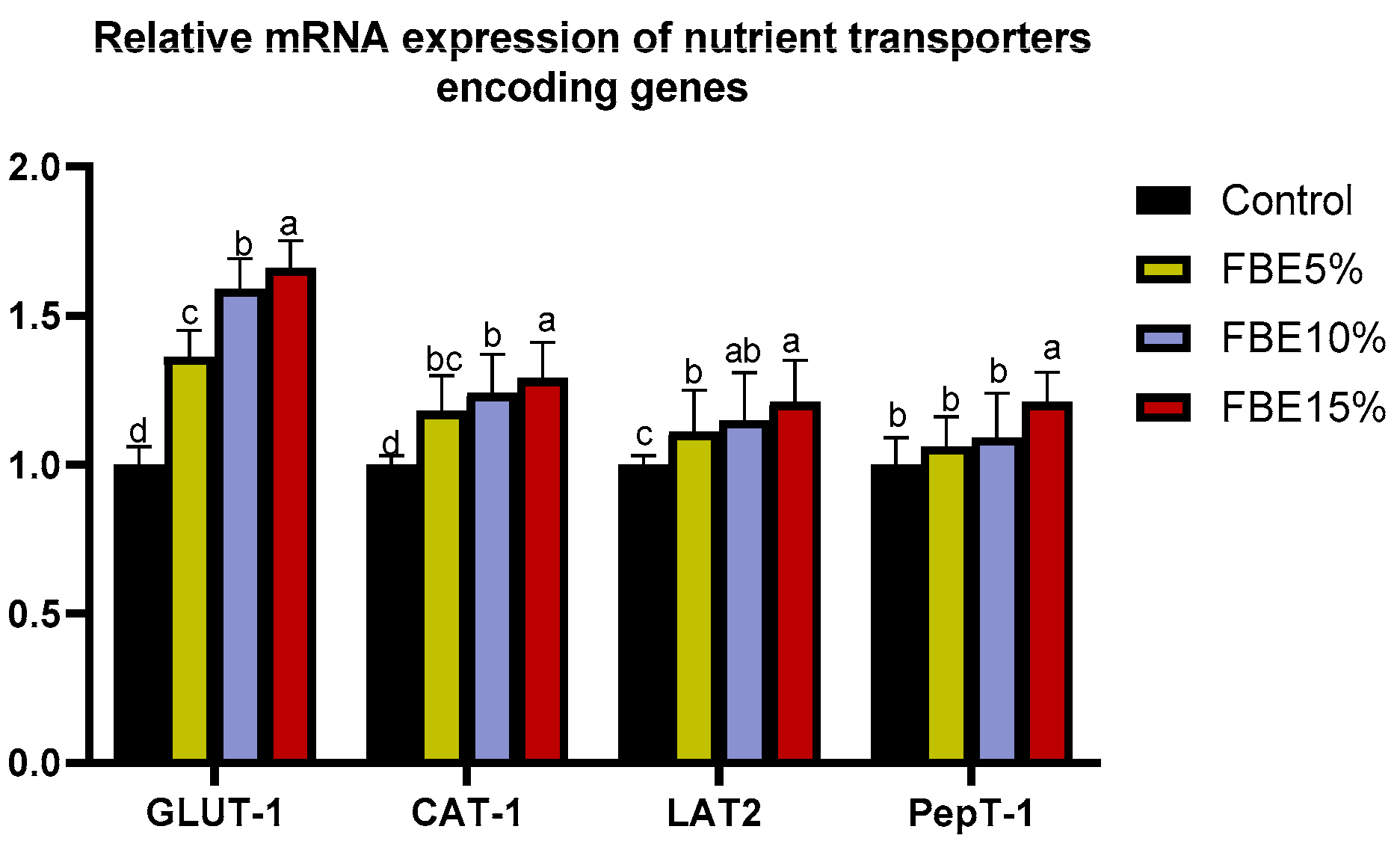
3.6. Intestinal Nutrient Transporter-Related Genes Expression
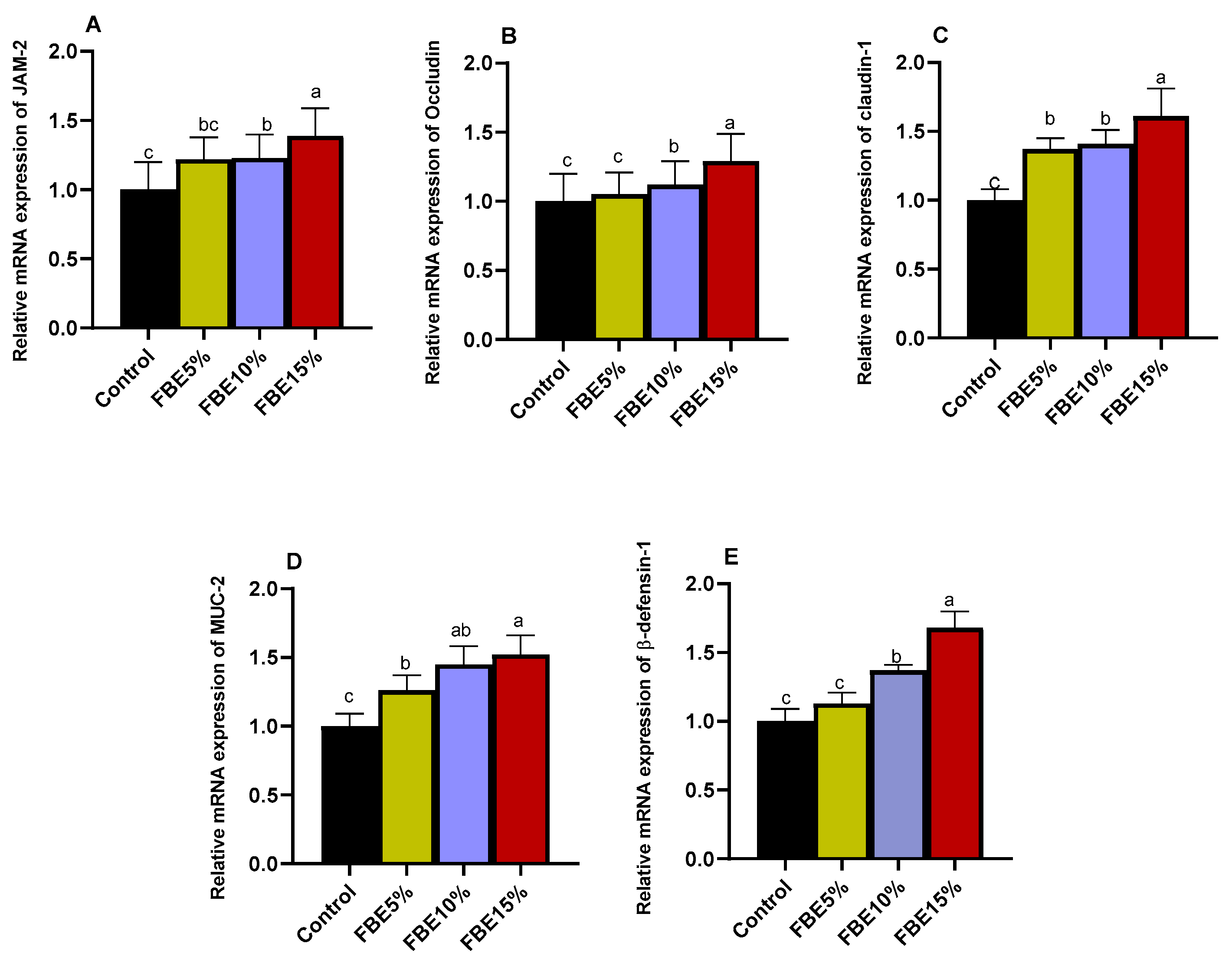
3.7. Intestinal Tight Junction-Related Genes Expression
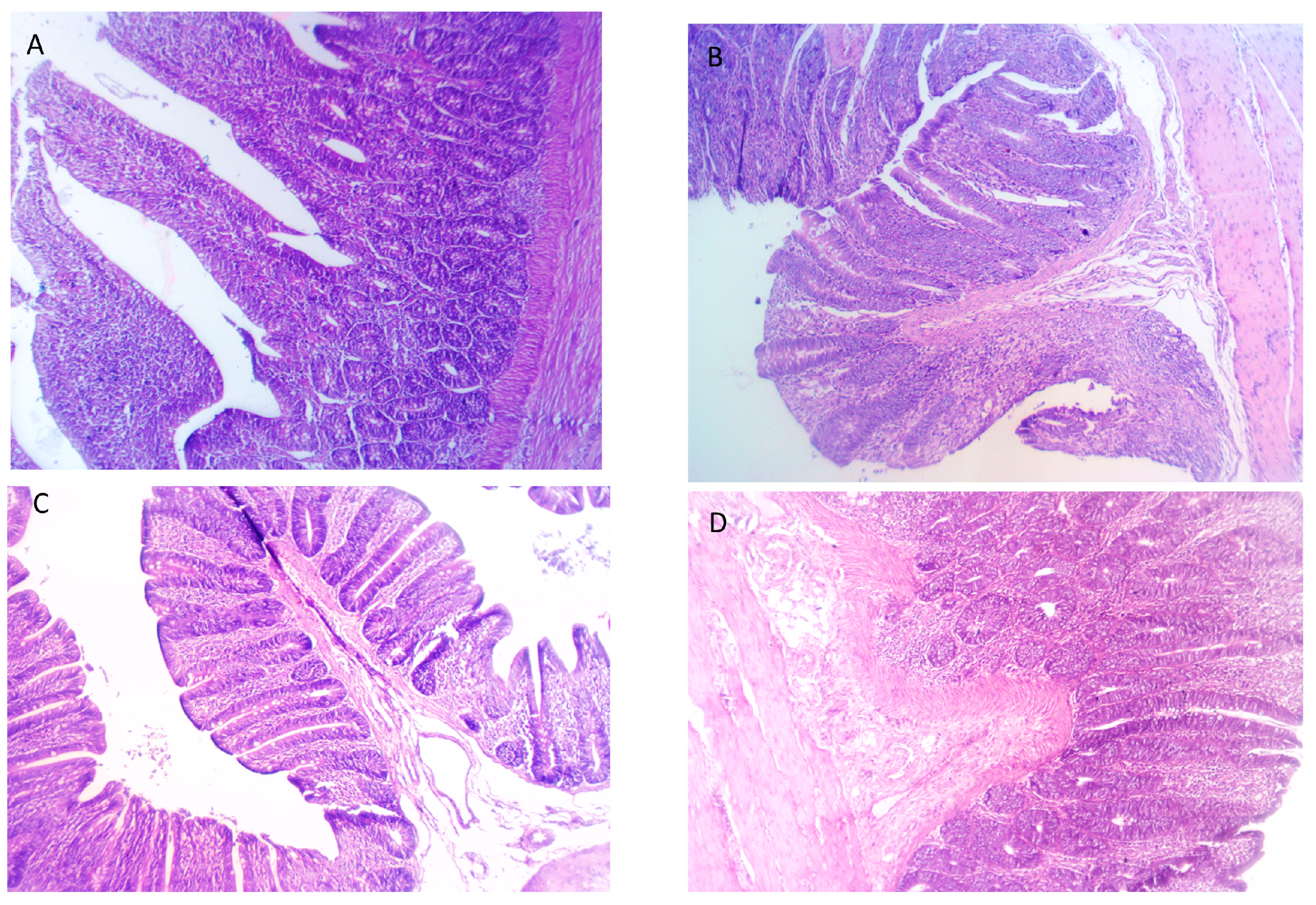
3.8. Evaluating Intestinal Histomorphology in Response to Feeding on FBEs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thirumalaisamy, G.; Muralidharan, J.; Senthilkumar, S.; Hema Sayee, R.; Priyadharsini, M. Cost-effective feeding of poultry. Int. J. Sci. Environ. Technol. 2016, 5, 3997–4005. [Google Scholar]
- Perera, W.; Abdollahi, M.; Ravindran, V.; Zaefarian, F.; Wester, T.; Ravindran, G. Nutritional evaluation of two barley cultivars, without and with carbohydrase supplementation, for broilers: Metabolisable energy and standardised amino acid digestibility. Br. Poult. Sci. 2019, 60, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Messia, M.C.; Candigliota, T.; De Arcangelis, E.; Marconi, E. Arabinoxylans and [beta]-glucans assessment in cereals. Ital. J. Food Sci. 2017, 29, 112–123. [Google Scholar]
- Kim, C.; Kang, H. Effects of fermented barley or wheat as feed supplement on growth performance, gut health and meat quality of broilers. Eur. Poult. Sci. 2016, 80, 1–11. [Google Scholar]
- Al-Khalaifah, H.S.; Shahin, S.; Omar, A.E.; Mohammed, H.A.; Mahmoud, H.; Ibrahim, D. Effects of graded levels of microbial fermented or enzymatically treated dried brewer’s grains on growth, digestive and nutrient transporter genes expression and cost effectiveness in broiler chickens. BMC Vet. Res. 2020, 16, 424. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Lázaro, R.; Latorre, M.; Gracia, M.; Mateos, G. Influence of enzyme supplementation and heat processing of barley on digestive traits and productive performance of broilers. Poult. Sci. 2008, 87, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wareth, A.A.; Mohamed, E.M.; Hassan, H.A.; Eldeek, A.A.; Lohakare, J. Effect of substituting hydroponic barley forage with or without enzymes on performance of growing rabbits. Sci. Rep. 2023, 13, 943. [Google Scholar] [CrossRef]
- Sharma, R.; Garg, P.; Kumar, P.; Bhatia, S.K.; Kulshrestha, S. Microbial fermentation and its role in quality improvement of fermented foods. Fermentation 2020, 6, 106. [Google Scholar] [CrossRef]
- Sugiharto, S.; Yudiarti, T.; Isroli, I. Performances and haematological profile of broilers fed fermented dried cassava (Manihot esculenta Crantz). Trop. Anim. Health Prod. 2016, 48, 1337–1341. [Google Scholar] [CrossRef]
- Abdel-Raheem, S.M.; Mohammed, E.S.Y.; Mahmoud, R.E.; El Gamal, M.F.; Nada, H.S.; El-Ghareeb, W.R.; Marzok, M.; Meligy, A.M.; Abdulmohsen, M.; Ismail, H. Double-fermented soybean meal totally replaces soybean meal in broiler rations with favorable impact on performance, digestibility, amino acids transporters and meat nutritional value. Animals 2023, 13, 1030. [Google Scholar] [CrossRef]
- Ibrahim, D.; Moustafa, A.; Shahin, S.; Sherief, W.; Farag, M.; Nassan, M. Impact of fermented or enzymatically fermented dried olive pomace on growth, expression of digestive enzymes and glucose transporters genes, oxidative stability of frozen meat and economic efficiency of broiler chickens. Front. Vet. Sci. 2021, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Stanbury, P.F.; Whitaker, A.; Hall, S.J. Principles of Fermentation Technology; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Omar, A.E.; Al-Khalaifah, H.S.; Ismail, T.A.; Abd El-Aziz, R.M.; El-Mandrawy, S.A.; Shalaby, S.I.; Ibrahim, D. Performance, serum biochemical and immunological parameters, and digestive enzyme and intestinal barrier-related gene expression of broiler chickens fed fermented fava bean by-products as a substitute for conventional feed. Front. Vet. Sci. 2021, 8, 696841. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhang, B.; Li, J.; Zhu, L. Effects of fermented feed on growth performance, immune response, and antioxidant capacity in laying hen chicks and the underlying molecular mechanism involving nuclear factor-κB. Poult. Sci. 2020, 99, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Agyekum, A.; Beaulieu, A.; Pieper, R.; Van Kessel, A.G. Fermentation of barley and wheat with lactic acid bacteria and exogenous enzyme on nutrient composition, microbial count, and fermentative characteristics. Can. J. Anim. Sci. 2020, 101, 106–117. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International; Association of Official Analytical Chemists: Rockville, MD, USA, 2023. [Google Scholar]
- Van Soest, P.v.; Robertson, J.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Kirkpatrick, D.S.; Bishop, S.H. Simplified wet ash procedure for total phosphorus analysis of organophosphonates in biological samples. Anal. Chem. 1971, 43, 1707–1709. [Google Scholar] [CrossRef]
- Gao, Y.; Shang, C.; Maroof, M.S.; Biyashev, R.; Grabau, E.; Kwanyuen, P.; Burton, J.; Buss, G. A modified colorimetric method for phytic acid analysis in soybean. Crop Sci. 2007, 47, 1797–1803. [Google Scholar] [CrossRef]
- ISO 30024:2009; Animal Feeding Stuffs—Determination of Phytase Activity. International Organization for Standardization: Geneva, Switzerland, 2009.
- Aviagen. Ross 308: Broiler’s Management and Nutrition Specification; Aviagen: Huntsville, AL, USA, 2018. [Google Scholar]
- Janssen, W. European Table of Energy Values for Poultry Feedstuffs; Spelderholt Center for Poultry Research and Information Services: Beekbergen, The Netherlands, 1989. [Google Scholar]
- Souza, O.; Adams, C.; Rodrigues, B.; Krause, A.; Bonamigo, R.; Zavarize, K.; Stefanello, C. The impact of Bacillus subtilis PB6 and chromium propionate on the performance, egg quality and nutrient metabolizability of layer breeders. Animals 2021, 11, 3084. [Google Scholar] [CrossRef]
- Zhang, C.; Hao, E.; Chen, X.; Huang, C.; Liu, G.; Chen, H.; Wang, D.; Shi, L.; Xuan, F.; Chang, D. Dietary Fiber Level Improve Growth Performance, Nutrient Digestibility, Immune and Intestinal Morphology of Broilers from Day 22 to 42. Animals 2023, 13, 1227. [Google Scholar] [CrossRef]
- Perera, W.; Abdollahi, M.; Zaefarian, F.; Wester, T.; Ravindran, G.; Ravindran, V. Influence of inclusion level of barley in wheat-based diets and supplementation of carbohydrase on growth performance, nutrient utilisation and gut morphometry in broiler starters. Br. Poult. Sci. 2019, 60, 736–748. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Puente, E.; Sánchez-Martín, M.A.; de Andrés, J.; Rodríguez-Izquierdo, L.; Méndez, L.; Palomero, J. Expression and functional analysis of the hydrogen peroxide biosensors HyPer and HyPer2 in C2C12 myoblasts/myotubes and single skeletal muscle fibres. Sci. Rep. 2020, 10, 871. [Google Scholar] [CrossRef] [PubMed]
- Pranczk, J.; Jacewicz, D.; Wyrzykowski, D.; Chmurzynski, L. Analytical methods for determination of reactive oxygen species. Curr. Pharm. Anal. 2014, 10, 293–304. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Aptekmann, K.; Artoni, S.B.; Stefanini, M.; Orsi, M. Morphometric analysis of the intestine of domestic quails (Coturnix coturnix japonica) treated with different levels of dietary calcium. Anat. Histol. Embryol. 2001, 30, 277–280. [Google Scholar] [CrossRef]
- Sakamoto, K.; Hirose, H.; Onizuka, A.; Hayashi, M.; Futamura, N.; Kawamura, Y.; Ezaki, T. Quantitative study of changes in intestinal morphology and mucus gel on total parenteral nutrition in rats. J. Surg. Res. 2000, 94, 99–106. [Google Scholar] [CrossRef]
- Abdollahi, M.; Ravindran, V. Alternative feed ingredients for poultry diets: Challenges and prospects. In Proceedings of the Australian Poultry Science Symposium, Sydney, Australia, 17–20 February 2019; pp. 102–108. [Google Scholar]
- Jacob, J.; Pescatore, A. Using barley in poultry diets—A review. J. Appl. Poult. Res. 2012, 21, 915–940. [Google Scholar] [CrossRef]
- Amerah, A. Interactions between wheat characteristics and feed enzyme supplementation in broiler diets. Anim. Feed Sci. Technol. 2015, 199, 1–9. [Google Scholar] [CrossRef]
- Shi, C.; Zhang, Y.; Lu, Z.; Wang, Y. Solid-state fermentation of corn-soybean meal mixed feed with Bacillus subtilis and Enterococcus faecium for degrading antinutritional factors and enhancing nutritional value. J. Anim. Sci. Biotechnol. 2017, 8, 50. [Google Scholar] [CrossRef]
- Cheng, Y.-H.; Hsiao, F.S.-H.; Wen, C.-M.; Wu, C.-Y.; Dybus, A.; Yu, Y.-H. Mixed fermentation of soybean meal by protease and probiotics and its effects on the growth performance and immune response in broilers. J. Appl. Anim. Res. 2019, 47, 339–348. [Google Scholar] [CrossRef]
- Azeez, S.O.; Chinma, C.E.; Bassey, S.O.; Eze, U.R.; Makinde, A.F.; Sakariyah, A.A.; Okubanjo, S.S.; Danbaba, N.; Adebo, O.A. Impact of germination alone or in combination with solid-state fermentation on the physicochemical, antioxidant, in vitro digestibility, functional and thermal properties of brown finger millet flours. LWT 2022, 154, 112734. [Google Scholar] [CrossRef]
- Stoffel, F.; Santana, W.D.O.; Fontana, R.C.; Gregolon, J.G.N.; Kist, T.B.L.; De Siqueira, F.G.; Mendonça, S.; Camassola, M. Chemical features and bioactivity of grain flours colonized by macrofungi as a strategy for nutritional enrichment. Food Chem. 2019, 297, 124988. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Lu, H.; Shu, Q.; Chen, Q.; Wang, J. The Influence of Different Pretreatment Methods of Highland Barley by Solid-State Fermentation with Agaricus sinodeliciosus var. Chaidam ZJU-TP-08 on Its Nutrient Content, Functional Properties and Physicochemical Characteristics. J. Fungi 2022, 8, 940. [Google Scholar] [CrossRef] [PubMed]
- Dallagnol, A.M.; Pescuma, M.; De Valdez, G.F.; Rollán, G. Fermentation of quinoa and wheat slurries by Lactobacillus plantarum CRL 778: Proteolytic activity. Appl. Microbiol. Biotechnol. 2013, 97, 3129–3140. [Google Scholar] [CrossRef]
- Blake, T.; Blake, V.C.; Bowman, J.G.; Abdel-Haleem, H. Barley feed uses and quality improvement. In Barley: Production, Improvement, and Uses; Ministry of Education Ethiopia: Addis Ababa, Ethiopia, 2011; pp. 522–531. [Google Scholar]
- Bedford, M.R. The effect of enzymes on digestion. J. Appl. Poult. Res. 1996, 5, 370–378. [Google Scholar] [CrossRef]
- Yeh, R.H.; Hsieh, C.W.; Chen, K.L. Screening lactic acid bacteria to manufacture two-stage fermented feed and pelleting to investigate the feeding effect on broilers. Poult. Sci. 2018, 97, 236–246. [Google Scholar] [CrossRef]
- Teng, D.; Gao, M.; Yang, Y.; Liu, B.; Tian, Z.; Wang, J. Bio-modification of soybean meal with Bacillus subtilis or Aspergillus oryzae. Biocatal. Agric. Biotechnol. 2012, 1, 32–38. [Google Scholar] [CrossRef]
- Xiao, X.; Li, J.; Xiong, H.; Tui, W.; Zhu, Y.; Zhang, J. Effect of extrusion or fermentation on physicochemical and digestive properties of barley powder. Front. Nutr. 2022, 8, 794355. [Google Scholar] [CrossRef]
- Yasar, S.; Desen, E.D. Analytical transferability of a universal method to determine in vitro activities of different phytase products in feed and their in vivo efficacies in Japanese quails (Coturnix coturnix japonica). Bull. UASVM Anim. Sci. Biotechnol. 2014, 71, 36–50. [Google Scholar]
- Yasar, S.; Tosun, R. Predicting chemical, enzymatic and nutritional properties of fermented barley (Hordeum vulgare L.) by second derivate spectra analysis from attenuated total reflectance—Fourier transform infrared data and its nutritional value in Japanese quails. Arch. Anim. Nutr. 2018, 72, 407–423. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, J.; Wang, Y.; Lv, J.; Li, J.; Guo, L.; Min, Y. Fermented corn–soybean meal mixed feed modulates intestinal morphology, barrier functions and cecal microbiota in laying hens. Animals 2021, 11, 3059. [Google Scholar] [CrossRef] [PubMed]
- Varsha, K.K.; Priya, S.; Devendra, L.; Nampoothiri, K.M. Control of spoilage fungi by protective lactic acid bacteria displaying probiotic properties. Appl. Biochem. Biotechnol. 2014, 172, 3402–3413. [Google Scholar] [CrossRef] [PubMed]
- Canibe, N.; Jensen, B.B. Fermented liquid feed—Microbial and nutritional aspects and impact on enteric diseases in pigs. Anim. Feed Sci. Technol. 2012, 173, 17–40. [Google Scholar] [CrossRef]
- Li, L.; Chou, X.; Yang, H. Optimization of fermentation conditions for broiler feed and its application effect. Feed Res. 2019, 491, 19–22. [Google Scholar]
- Skrede, G.; Herstad, O.; Sahlstrøm, S.; Holck, A.; Slinde, E.; Skrede, A. Effects of lactic acid fermentation on wheat and barley carbohydrate composition and production performance in the chicken. Anim. Feed Sci. Technol. 2003, 105, 135–148. [Google Scholar] [CrossRef]
- Gong, L.; Wang, B.; Mei, X.; Xu, H.; Qin, Y.; Li, W.; Zhou, Y. Effects of three probiotic Bacillus on growth performance, digestive enzyme activities, antioxidative capacity, serum immunity, and biochemical parameters in broilers. Anim. Sci. J. 2018, 89, 1561–1571. [Google Scholar] [CrossRef]
- Yaghobfar, A.; Kalantar, M. Effect of non-starch polysaccharide (NSP) of wheat and barley supplemented with exogenous enzyme blend on growth performance, gut microbial, pancreatic enzyme activities, expression of glucose transporter (SGLT1) and mucin producer (MUC2) genes of broiler chickens. Braz. J. Poult. Sci. 2017, 19, 629–638. [Google Scholar]
- Horvatovic, M.; Glamocic, D.; Zikic, D.; Hadnadjev, T. Performance and some intestinal functions of broilers fed diets with different inclusion levels of sunflower meal and supplemented or not with enzymes. Braz. J. Poult. Sci. 2015, 17, 25–30. [Google Scholar] [CrossRef]
- Perera, W.; Abdollahi, M.; Zaefarian, F.; Wester, T.; Ravindran, V. The effect of graded inclusions of waxy starch hull-less barley and a multi-component exogenous carbohydrase on the growth performance, nutrient digestibility and intestinal morphometry of broiler chickens. Br. Poult. Sci. 2020, 61, 442–453. [Google Scholar] [CrossRef]
- Elbaz, A.M.; El-Sheikh, S.E.; Abdel-Maksoud, A. Growth performance, nutrient digestibility, antioxidant state, ileal histomorphometry, and cecal ecology of broilers fed on fermented canola meal with and without exogenous enzymes. Trop. Anim. Health Prod. 2023, 55, 46. [Google Scholar] [CrossRef]
- Liu, S.; Du, M.; Tu, Y.; You, W.; Chen, W.; Liu, G.; Li, J.; Wang, Y.; Lu, Z.; Wang, T. Fermented mixed feed alters growth performance, carcass traits, meat quality and muscle fatty acid and amino acid profiles in finishing pigs. Anim. Nutr. 2023, 12, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Jiang, Z.; Wang, C.; Zhang, Y.; Gong, T.; Wang, F.; Jin, M.; Wang, Y.; Lu, Z. Co-fermented defatted rice bran alters gut microbiota and improves growth performance, antioxidant capacity, immune status and intestinal permeability of finishing pigs. Anim. Nutr. 2022, 11, 413–424. [Google Scholar] [CrossRef]
- Hui, Y.; Tamez-Hidalgo, P.; Cieplak, T.; Satessa, G.D.; Kot, W.; Kjærulff, S.; Nielsen, M.O.; Nielsen, D.S.; Krych, L. Supplementation of a lacto-fermented rapeseed-seaweed blend promotes gut microbial-and gut immune-modulation in weaner piglets. J. Anim. Sci. Biotechnol. 2021, 12, 85. [Google Scholar] [CrossRef]
- Lv, J.; Guo, L.; Chen, B.; Hao, K.; Ma, H.; Liu, Y.; Min, Y. Effects of different probiotic fermented feeds on production performance and intestinal health of laying hens. Poult. Sci. 2022, 101, 101570. [Google Scholar] [CrossRef]
- Lawal, T.; Iyayi, E.; Adeniyi, B.; Adaramoye, O. Biodegradation of palm kernel cake with multienzyme complexes from fungi and its feeding value for broilers. Int. J. Poult. Sci. 2010, 9, 695–701. [Google Scholar] [CrossRef]
- Khalil, M.; Abdollahi, M.; Zaefarian, F.; Chrystal, P.; Ravindran, V. Apparent metabolizable energy of cereal grains for broiler chickens is influenced by age. Poult. Sci. 2021, 100, 101288. [Google Scholar] [CrossRef] [PubMed]
- McNab, J.; Smithard, R. Barley β-glucan: An antinutritional factor in poultry feeding. Nutr. Res. Rev. 1992, 5, 45–60. [Google Scholar] [CrossRef]
- Peng, W.; Talpur, M.Z.; Zeng, Y.; Xie, P.; Li, J.; Wang, S.; Wang, L.; Zhu, X.; Gao, P.; Jiang, Q. Influence of fermented feed additive on gut morphology, immune status, and microbiota in broilers. BMC Vet. Res. 2022, 18, 218. [Google Scholar] [CrossRef]
- Drażbo, A.; Ognik, K.; Zaworska, A.; Ferenc, K.; Jankowski, J. The effect of raw and fermented rapeseed cake on the metabolic parameters, immune status, and intestinal morphology of turkeys. Poult. Sci. 2018, 97, 3910–3920. [Google Scholar] [CrossRef]
- Peng, W.; Talpur, M.Z.; Zeng, Y.; Xie, P.; Li, J.; Wang, S.; Wang, L.; Zhu, X.; Gao, P.; Jiang, Q. Effect of fermented feed additive on the broilers’ histology/morphology of the GIT, immune status, and microbiota. Res. Sq. 2021, preprint. [Google Scholar] [CrossRef]
- El-Sanhoury, M.; Ahmed, A. Broiler performance, enzymes activity and histological observations affected by multi enzymes complex (ZADO®). Egypt. J. Nutr. Feed. 2017, 20, 309–320. [Google Scholar] [CrossRef]
- El-Ghamry, A.; Al-Harthi, M.; Attia, Y. Possibility to improve rice polishing utilisation in broiler diets by enzymes or dietary formulation based on digestible amino acids. Arch. Geflugelk. 2005, 69, 49–56. [Google Scholar]
- Bedford, M.R. The evolution and application of enzymes in the animal feed industry: The role of data interpretation. Br. Poult. Sci. 2018, 59, 486–493. [Google Scholar] [CrossRef] [PubMed]
- González-Ortiz, G.; Kozłowski, K.; Drażbo, A.; Bedford, M.R. Response of turkeys fed wheat-barley-rye based diets to xylanase supplementation. Anim. Feed Sci. Technol. 2017, 229, 117–123. [Google Scholar] [CrossRef]
- Kanhar, A.R.; Hussain, S.F.; Shar, A.H.; Ujjan, J.A. Isolation of A Novel Bacterial Strain Planococcus Plakortidi from Ross Broiler Chicken Gut (CECUM) with Combination of Non-Starch Polysaccride Degrading Pectinase Enzyme. Pak. J. Med. Health Sci. 2022, 16, 962. [Google Scholar] [CrossRef]
- Perera, W.N.U.; Abdollahi, M.R.; Zaefarian, F.; Wester, T.J.; Ravindran, V. Barley, an undervalued cereal for poultry diets: Limitations and opportunities. Animals 2022, 12, 2525. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M. The role of exogenous enzymes in promoting growth and improving nutrient digestibility in poultry. Iran. J. Vet. Res. 2018, 19, 157. [Google Scholar]
- Lijie, Y.; Xiangfang, Z.; Shiyan, Q. Research progress of non starch polysaccharides in the regulation of intestinal flora in pigs. Biotechnol. Bull. 2020, 36, 9–16. [Google Scholar]
- Ruhnke, I.; Röhe, I.; Goodarzi Boroojeni, F.; Knorr, F.; Mader, A.; Hafeez, A.; Zentek, J. Feed supplemented with organic acids does not affect starch digestibility, nor intestinal absorptive or secretory function in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2015, 99, 29–35. [Google Scholar] [CrossRef]
- Saleh, A.A.; El-Far, A.H.; Abdel-Latif, M.A.; Emam, M.A.; Ghanem, R.; Abd El-Hamid, H.S. Exogenous dietary enzyme formulations improve growth performance of broiler chickens fed a low-energy diet targeting the intestinal nutrient transporter genes. PLoS ONE 2018, 13, e0198085. [Google Scholar] [CrossRef]
- Ferraris, R.P. Dietary and developmental regulation of intestinal sugar transport. Biochem. J. 2001, 360, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Wiseman, J.; Masey O’Neill, H.V.; Scholey, D.V.; Burton, E.J.; Hill, S.E. Understanding the direct and indirect mechanisms of xylanase action on starch digestion in broilers. J. World’s Poult. Res. 2017, 7, 35–47. [Google Scholar]
- Hosseini, S.M.; Manafi, M.; Nazarizadeh, H. Effects of xylanase supplementation and citric acid on performance, ileal nutrients digestibility, and gene expression of intestinal nutrient transporters in broilers challenged with Clostridium perfringens. J. Poult. Sci. 2017, 54, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Arthur, C.P.; Ciferri, C.; Matsumoto, M.L. Structure of the secretory immunoglobulin A core. Science 2020, 367, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Gharib-Naseri, K.; de Paula Dorigam, J.C.; Doranalli, K.; Kheravii, S.; Swick, R.A.; Choct, M.; Wu, S.-B. Modulations of genes related to gut integrity, apoptosis, and immunity underlie the beneficial effects of Bacillus amyloliquefaciens CECT 5940 in broilers fed diets with different protein levels in a necrotic enteritis challenge model. J. Anim. Sci. Biotechnol. 2020, 11, 104. [Google Scholar] [CrossRef]
- Johansson, M.E.; Larsson, J.M.H.; Hansson, G.C. The two mucus layers of colon are organized by the MUC2 mucin, whereas the outer layer is a legislator of host–microbial interactions. Proc. Natl. Acad. Sci. USA 2011, 108, 4659–4665. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, E.E.; Lynch, R.D. The tight junction: A multifunctional complex. Am. J. Physiol. Cell Physiol. 2004, 286, C1213–C1228. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef]
- Farahat, M.; Ibrahim, D.; Kishawy, A.; Abdallah, H.; Hernandez-Santana, A.; Attia, G. Effect of cereal type and plant extract addition on the growth performance, intestinal morphology, caecal microflora, and gut barriers gene expression of broiler chickens. Animal 2021, 15, 100056. [Google Scholar] [CrossRef]
- Sohail, M.U.; Ijaz, A.; Younus, M.; Shabbir, M.Z.; Kamran, Z.; Ahmad, S.; Anwar, H.; Yousaf, M.S.; Ashraf, K.; Shahzad, A. Effect of supplementation of mannan oligosaccharide and probiotic on growth performance, relative weights of viscera, and population of selected intestinal bacteria in cyclic heat-stressed broilers. J. Appl. Poult. Res. 2013, 22, 485–491. [Google Scholar] [CrossRef]
- Kong, X.; Wu, G.; Liao, Y.; Hou, Z.; Liu, H.; Yin, F.; Li, T.; Huang, R.; Zhang, Y.; Deng, D. Dietary supplementation with Chinese herbal ultra-fine powder enhances cellular and humoral immunity in early-weaned piglets. Livest. Sci. 2007, 108, 94–98. [Google Scholar] [CrossRef]
- Kishawy, A.T.; Al-Khalaifah, H.S.; Nada, H.S.; Roushdy, E.M.; Zaglool, A.W.; Ahmed Ismail, T.; Ibrahim, S.M.; Ibrahim, D. Black Pepper or Radish Seed Oils in a New Combination of Essential Oils Modulated Broiler Chickens’ Performance and Expression of Digestive Enzymes, Lipogenesis, Immunity, and Autophagy-Related Genes. Vet. Sci. 2022, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, W.; Liu, S.; Wang, H. Probiotic fermented feed improved the production, health and nutrient utilisation of yellow-feathered broilers reared in high altitude in Tibet. Br. Poult. Sci. 2020, 61, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Sun, H.; Yao, X.; Wu, Y.; Wang, X.; Feng, J. Effects of replacement of soybean meal by fermented cottonseed meal on growth performance, serum biochemical parameters and immune function of yellow-feathered broilers. Asian-Australas. J. Anim. Sci. 2012, 25, 393. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, Z.; Wang, G.; Li, Y.; Qi, Y. Effects of feed supplemented with fermented pine needles (Pinus ponderosa) on growth performance and antioxidant status in broilers. Poult. Sci. 2015, 94, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zeng, X.; Ding, X. Effects of replacing soybean meal with fermented rapeseed meal on performance, serum biochemical variables and intestinal morphology of broilers. Asian-Australas J. Anim. Sci. 2012, 25, 1734. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Effects of dietary Enterococcus faecium NCIMB 11181 supplementation on growth performance and cellular and humoral immune responses in broiler chickens. Poult. Sci. 2019, 98, 150–163. [Google Scholar] [CrossRef]
- Zhu, X.; Tao, L.; Liu, H.; Yang, G. Effects of fermented feed on growth performance, immune organ indices, serum biochemical parameters, cecal odorous compound production, and the microbiota community in broilers. Poult. Sci. 2023, 102, 102629. [Google Scholar] [CrossRef]
- Wells, S.M.; Holian, A. Asymmetric dimethylarginine induces oxidative and nitrosative stress in murine lung epithelial cells. Am. J. Respir. Cell Mol. Biol. 2007, 36, 520–528. [Google Scholar] [CrossRef]
- Berndt, C.; Lillig, C.H. Glutathione, glutaredoxins, and iron. Antioxid. Redox Signal. 2017, 27, 1235–1251. [Google Scholar] [CrossRef]
- Son, H.-K.; Kang, S.-T.; Lee, J.-J. Effects of Peucedanum japonicum Thunb. on lipid metabolism and antioxidative activities in rats fed a high-fat/high-cholesterol diet. J. Korean Soc. Food Sci. Nutr. 2014, 43, 641–649. [Google Scholar] [CrossRef]
- Mayahi, M.; Jalali, M.R.; Kiani, R. Effects of dietary probiotic supplementation on cholesterol and triglyceride levels in broiler chicks’ sera. In Proceedings of the World Poultry Science Association (WPSA), 2nd Mediterranean Summit of WPSA, Antalya, Turkey, 4–7 October 2009; pp. 4–7. [Google Scholar]
- Klaver, F.; Van der Meer, R. The assumed assimilation of cholesterol by Lactobacilli and Bifidobacterium bifidum is due to their bile salt-deconjugating activity. Appl. Environ. Microbiol. 1993, 59, 1120–1124. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Yudiarti, T.; Isroli, I.; Widiastuti, E.; Wahyuni, H.I.; Sartono, T.A. Fermented feed as a potential source of natural antioxidants for broiler chickens—A mini review. Agric. Conspec. Sci. 2019, 84, 313–318. [Google Scholar]
- Ibrahim, D.; Kishawy, A.T.; Khater, S.I.; Hamed Arisha, A.; Mohammed, H.A.; Abdelaziz, A.S.; El-Rahman, A.; Ghada, I.; Elabbasy, M.T. Effect of dietary modulation of selenium form and level on performance, tissue retention, quality of frozen stored meat and gene expression of antioxidant status in Ross broiler chickens. Animals 2019, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Kishawy, A.T.; Ibrahim, D.; Roushdy, E.M.; Moustafa, A.; Eldemery, F.; Hussein, E.M.; Hassan, F.A.; Elazab, S.T.; Elabbasy, M.T.; Kanwal, R. Impact of resveratrol-loaded liposomal nanocarriers on heat-stressed broiler chickens: Effects on performance, sirtuin expression, oxidative stress regulators, and muscle building factors. Front. Vet. Sci. 2023, 10, 1137896. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, Q.; Song, D.; Wang, W.; Zhou, H.; Wang, L.; Li, A. Effects of fermented cottonseed meal on growth performance, serum biochemical parameters, immune functions, antioxidative abilities, and cecal microflora in broilers. Food Agric. Immunol. 2017, 28, 725–738. [Google Scholar] [CrossRef]
- Martins, S.; Mussatto, S.I.; Martínez-Avila, G.; Montañez-Saenz, J.; Aguilar, C.N.; Teixeira, J.A. Bioactive phenolic compounds: Production and extraction by solid-state fermentation. A review. Biotechnol. Adv. 2011, 29, 365–373. [Google Scholar] [CrossRef]
- Sumida, S.; Tanaka, K.; Kitao, H.; Nakadomo, F. Exercise-induced lipid peroxidation and leakage of enzymes before and after vitamin E supplementation. Int. J. Biochem. 1989, 21, 835–838. [Google Scholar]
- Lin, C.-H.; Wei, Y.-T.; Chou, C.-C. Enhanced antioxidative activity of soybean koji prepared with various filamentous fungi. Food Microbiol. 2006, 23, 628–633. [Google Scholar] [CrossRef]
- Deng, G.; Wang, J.; Zhang, Q.; He, H.; Wu, F.; Feng, T.; Zhou, J.; Zou, K.; Hattori, M. Hepatoprotective effects of phloridzin on hepatic fibrosis induced by carbon tetrachloride against oxidative stress-triggered damage and fibrosis in rats. Biol. Pharm. Bull. 2012, 35, 1118–1125. [Google Scholar] [CrossRef]
- Yang, J.; Wu, X.-B.; Chen, H.-L.; Sun-waterhouse, D.; Zhong, H.-B.; Cui, C. A value-added approach to improve the nutritional quality of soybean meal byproduct: Enhancing its antioxidant activity through fermentation by Bacillus amyloliquefaciens SWJS22. Food Chem. 2019, 272, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yan, W.; Ma, C.; Liu, Y.; Gong, L.; Levesque, C.; Dong, B. Effects of supplemented culture media from solid-state fermented Isaria cicadae on performance, serum biochemical parameters, serum immune indexes, antioxidant capacity and meat quality of broiler chickens. Asian-Australas. J. Anim. Sci. 2020, 33, 568. [Google Scholar] [CrossRef] [PubMed]
- El-Ghareeb, W.R.; Kishawy, A.T.; Anter, R.G.; Aboelabbas Gouda, A.; Abdelaziz, W.S.; Alhawas, B.; Meligy, A.M.; Abdel-Raheem, S.M.; Ismail, H.; Ibrahim, D. Novel Antioxidant Insights of Myricetin on the Performance of Broiler Chickens and Alleviating Experimental Infection with Eimeria spp.: Crosstalk between Oxidative Stress and Inflammation. Antioxidants 2023, 12, 1026. [Google Scholar] [CrossRef]
- Ibrahim, D.; Sewid, A.H.; Arisha, A.H.; Abd El-Fattah, A.H.; Abdelaziz, A.M.; Al-Jabr, O.A.; Kishawy, A.T. Influence of Glycyrrhiza glabra Extract on Growth, Gene Expression of Gut Integrity, and Campylobacter jejuni Colonization in Broiler Chickens. Front. Vet. Sci 2020, 7, 612063. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Ingredients | Control | FBEs5% | FBEs10% | FBEs15% |
|---|---|---|---|---|
| Yellow corn | 58.80 | 55.30 | 50.30 | 45.80 |
| Soybean meal, 48% CP | 34.00 | 32.50 | 32.50 | 32.00 |
| Corn gluten | 1.20 | 1.20 | 1.20 | 1.20 |
| FBEs 1 | - | 5.00 | 10.00 | 15.00 |
| Soybean oil | 1.80 | 1.80 | 1.80 | 1.80 |
| Calcium carbonate | 1.20 | 1.20 | 1.20 | 1.20 |
| Calcium diphasic phosphate | 1.50 | 1.50 | 1.50 | 1.50 |
| Common salt | 0.30 | 0.30 | 0.30 | 0.30 |
| Premix 2 | 0.30 | 0.30 | 0.30 | 0.30 |
| L-Lysine | 0.35 | 0.35 | 0.35 | 0.35 |
| DL-Methionine | 0.25 | 0.25 | 0.25 | 0.25 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 |
| Anti-mycotoxin | 0.10 | 0.10 | 0.10 | 0.10 |
| Analyzed and calculated composition | ||||
| Metabolizable energy (kcal/kg) 3 | 3048 | 3038 | 3015 | 3000 |
| Crude protein (%) | 23.40 | 23.07 | 23.16 | 23.06 |
| Ether extract% | 4.28 | 4.22 | 4.12 | 4.03 |
| Crude fiber (%) | 2.63 | 2.67 | 2.70 | 2.75 |
| Calcium (%) | 1.04 | 1.04 | 1.03 | 1.03 |
| Available phosphorous (%) | 0.46 | 0.49 | 0.54 | 0.57 |
| Lysine (%) | 1.33 | 1.30 | 1.30 | 1.29 |
| Methionine (%) | 0.58 | 0.58 | 0.57 | 0.57 |
| Ingredients | Control | FBEs5% | FBEs10% | FBEs15% |
|---|---|---|---|---|
| Yellow corn | 63.30 | 59.30 | 55.20 | 50.80 |
| Soybean meal, 48% CP | 27.90 | 26.90 | 26.00 | 25.40 |
| Corn gluten | 2.00 | 2.00 | 2.00 | 2.00 |
| FBEs 1 | - | 5.00 | 10.00 | 15.00 |
| Soybean oil | 2.50 | 2.50 | 2.50 | 2.50 |
| Calcium carbonate | 1.20 | 1.20 | 1.20 | 1.20 |
| Calcium diphasic phosphate | 1.50 | 1.50 | 1.50 | 1.50 |
| Common salt | 0.30 | 0.30 | 0.30 | 0.30 |
| Premix 2 | 0.30 | 0.30 | 0.30 | 0.30 |
| L-Lysine | 0.45 | 0.45 | 0.45 | 0.45 |
| DL-Methionine | 0.25 | 0.25 | 0.25 | 0.25 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 |
| Anti-mycotoxin | 0.10 | 0.10 | 0.10 | 0.10 |
| Analyzed and calculated composition | ||||
| Metabolizable energy (kcal/kg) 3 | 3122 | 3128 | 3113 | 3105 |
| Crude protein (%) | 21.30 | 21.30 | 21.14 | 20.95 |
| Ether extract% | 5.01 | 5.01 | 4.93 | 4.85 |
| Crude fiber (%) | 2.52 | 2.56 | 2.60 | 2.63 |
| Calcium (%) | 1.02 | 1.02 | 1.02 | 1.01 |
| Available phosphorous (%) | 0.50 | 0.54 | 0.58 | 0.62 |
| Lysine (%) | 1.25 | 1.25 | 1.23 | 1.22 |
| Methionine (%) | 0.56 | 0.56 | 0.57 | 0.55 |
| Ingredients | Control | FBEs5% | FBEs10% | FBEs15% |
|---|---|---|---|---|
| Yellow corn | 68.00 | 63.00 | 59.00 | 54.90 |
| Soybean meal, 48% CP | 21.90 | 21.90 | 21.00 | 20.00 |
| Corn gluten | 2.50 | 2.50 | 2.50 | 2.50 |
| FBEs 1 | - | 5.00 | 10.00 | 15.00 |
| Soybean oil | 3.40 | 3.40 | 3.40 | 3.40 |
| Calcium carbonate | 1.20 | 1.20 | 1.20 | 1.20 |
| Calcium diphasic phosphate | 1.50 | 1.50 | 1.50 | 1.50 |
| Common salt | 0.30 | 0.30 | 0.30 | 0.30 |
| Premix 2 | 0.30 | 0.30 | 0.30 | 0.300 |
| L-Lysine | 0.35 | 0.35 | 0.35 | 0.35 |
| DL-Methionine | 0.25 | 0.25 | 0.25 | 0.25 |
| Choline chloride | 0.20 | 0.20 | 0.20 | 0.20 |
| Anti-mycotoxin | 0.10 | 0.10 | 0.10 | 0.100 |
| Analyzed and calculated composition | ||||
| Metabolizable energy (kcal/kg) 3 | 3214 | 3205 | 3209 | 3199 |
| Crude protein (%) | 19.47 | 19.56 | 19.32 | 19.15 |
| Ether extract% | 6.08 | 5.95 | 5.90 | 5.83 |
| Crude fiber (%) | 2.41 | 2.47 | 2.50 | 2.53 |
| Calcium (%) | 1.01 | 1.01 | 1.01 | 1.00 |
| Available phosphorous (%) | 0.44 | 0.47 | 0.50 | 0.52 |
| Lysine (%) | 1.05 | 1.05 | 1.05 | 1.03 |
| Methionine (%) | 0.54 | 0.54 | 0.54 | 0.50 |
| Encoding Gene | Primer Sequence (5′–3′) | Accession No. |
|---|---|---|
| Barrier functions | ||
| β-Defensin-1 | F: AAACCATTGTCAGCCCTGTG R: TTCCTAGAGCCTGGGAGGAT | NM_204993.1 |
| MUC-2 | F: AAACAACGGCCATGTTTCAT R: GTGTGACACTGGTGTGCTGA | NM_001318434 |
| CLDN-1 | F: GGTGAAGAAGATGCGGATGG R: TCTGGTGTTAACGGGTGTGA | NM_001013611 |
| Occludin | F: ACGGCAAAGCCAACATCTAC R: ATCCGCCACGTTCTTCAC | XM_031604121.1 |
| JAM-2 | F: AGACAGGAACAGGCAGTGCT R: TCCAATCCCATTTGAGGCTA | XM_031556661.1 |
| Nutrient transporters | ||
| GLUT1 | F: ACAACACCG GCGTCATCAA R: TTGACATCAGCATGGAGTTACG | NM_205209.1 |
| CAT1 | F: ATGTAGGTTGGGATGGAGCC R: AACGAGTAAGCCAGGAGGGT | XM_015277949.1 |
| LAT1 | F: CTCTCTCTCATCATCTGGGC R: TCATTCCTGGGTCTGTTGCT | XM_415975 |
| PepT1 | F: TTTCCTTTACATCCCTCTCC R:TCACTTCTACTCTCACTC | NM-204365 |
| House keeping | ||
| GAPDH | F: CAACCCCCAATGTCTCTGTT R: TCAGCAGCAGCCTTCACTAC | NM205518 |
| TBP | F: GTCCACGGTGAATCTTGGTT R: GCGCAGTAGTACGTGGTTCTC | Acc:8484 |
| Parameter | UFB 1 | FBEs 2 | SEM | p-Value |
|---|---|---|---|---|
| Dry matter (%) | 89.60 b | 90.90 a | 0.39 | <0.030 |
| Crude protein (%) | 10.75 b | 11.92 a | 0.11 | <0.001 |
| Crude fiber (%) | 5.30 a | 3.25 b | 0.39 | 0.040 |
| Ether extract (%) | 1.70 | 1.62 | 0.33 | 0.060 |
| Lignin (%) | 12.23 a | 8.96 b | 0.24 | <0.001 |
| Cellulose (%) | 14.69 a | 10.35 b | 0.77 | <0.001 |
| Hemicellulose (%) | 27.19 a | 28.22 b | 0.31 | <0.006 |
| Neutral detergent fiber (%) | 54.11 a | 47.53 b | 0.27 | <0.001 |
| Acid detergent fiber (%) | 26.96 a | 19.31 b | 0.37 | 0.030 |
| Reduced sugar (g/100 g) | 2.36 b | 8.19 a | 0.68 | 0.020 |
| pH | 6.25 a | 4.25 b | 0.15 | <0.006 |
| Total lactic acid bacteria (log CFU/g feed) | 4.57 b | 7.96 a | 0.50 | <0.001 |
| Bacillus spp. log CFU/g feed | 3.96 b | 6.33 a | 0.35 | <0.001 |
| Non-phytate phosphorus (g/kg) | 2.03 b | 2.82 a | 0.16 | <0.007 |
| Phytase activity (FTU/kg) | 59.36 b | 225.51 a | 0.83 | <0.001 |
| Item | Control | FBEs5% | FBEs10% | FBEs15% | SEM | p-Value |
|---|---|---|---|---|---|---|
| Starter (1–10 days) | ||||||
| Initial BW | 44.33 | 46.00 | 45.00 | 46.33 | 0.96 | 0.280 |
| BW (g/bird) | 307 b | 310 b | 335 a | 299 b | 13.36 | 0.020 |
| BWG (g/bird) | 262 b | 264 b | 290 a | 252 b | 14.35 | 0.004 |
| FCR | 1.39 a | 1.38 a | 1.24 b | 1.44 a | 0.01 | 0.007 |
| FI (g/bird) | 365 | 364 | 359 | 363 | 12.47 | 0.480 |
| Grower (11–20 days) | ||||||
| BW (g/bird) | 1157 b | 1176 b | 1221 a | 1181 b | 6.30 | <0.001 |
| BWG (g/bird) | 850 b | 865 ab | 886 a | 882 a | 8.40 | <0.001 |
| FCR | 1.79 a | 1.74 a | 1.64 b | 1.67 b | 0.02 | <0.001 |
| FI (g/bird) | 1521 a | 1507 ab | 1449 b | 1467 ab | 3.51 | 0.020 |
| Finisher (21–38 days) | ||||||
| BW (g/bird) | 2532 c | 2597 b | 2650 a | 2588 b | 12.83 | <0.001 |
| BWG (g/bird) | 1376 c | 1421 ab | 1429 a | 1407 b | 9.20 | 0.010 |
| FCR | 1.83 a | 1.75 b | 1.71 c | 1.76 b | <0.001 | <0.001 |
| FI (g/bird) | 2512 a | 2486 ab | 2447 b | 2480 ab | 2.60 | <0.001 |
| Overall (1–38 days) | ||||||
| BWG (g/bird) | 2488 c | 2551 b | 2605 a | 2541 b | 12.39 | 0.030 |
| FI (g/bird) | 4398 a | 4357 ab | 4255 c | 4301 bc | 11.32 | 0.020 |
| FCR | 1.77 a | 1.71 b | 1.63 c | 1.69 b | 0.01 | <0.001 |
| Item | Control | FBEs5% | FBEs10% | FBEs15% | SEM | p-Value |
|---|---|---|---|---|---|---|
| Nutrients metabolizability (%) | ||||||
| Dry matter | 78.99 c | 81.78 b | 83.26 a | 82.00 b | 0.63 | 0.001 |
| Crude protein | 72.12 c | 74.36 b | 76.36 a | 74.12 b | 0.75 | 0.020 |
| Crude fiber | 22.99 c | 25.56 b | 27.36 a | 25.24 b | 0.83 | 0.030 |
| Phosphorus | 48.69 d | 54.66 c | 60.36 b | 65.47 a | 0.29 | 0.040 |
| Duodenal pH | 6.12 a | 5.64 b | 5.42 bc | 5.12 c | 0.13 | 0.030 |
| Jejunal digesta viscosity | 3.65 | 3.69 | 3.52 | 3.72 | 0.11 | 0.060 |
| Digestive enzymes (U/mg protein) | ||||||
| Lipase | 30.93 b | 36.68 a | 37.53 a | 38.07 a | 0.44 | 0.001 |
| Amylase | 140.57 b | 145.60 a | 149.33 a | 146.27 a | 1.29 | 0.001 |
| Trypsin | 410 c | 469 b | 475 b | 492 a | 4.08 | 0.030 |
| Chymotrypsin | 12.80 c | 15.30 b | 16.40 ab | 20.91 a | 0.24 | 0.010 |
| Maltase | 153 c | 155 b | 163 a | 165 a | 1.32 | 0.001 |
| Sucrase | 54.10 c | 58.90 b | 60.40 a | 61.40 a | 0.0.12 | 0.010 |
| Item | Control | FBEs5% | FBEs10% | FBEs15% | SEM | p-Value |
|---|---|---|---|---|---|---|
| IgM (ng/L) | 2.10 b | 0.55 | 3.60 ab | 3.77 a | 0.55 | 0.020 |
| IgG (ng/L) | 1.83 b | 0.38 | 2.35 ab | 2.60 a | 0.38 | 0.090 |
| Lysozyme (U/mL) | 109.43 c | 3.30 | 116.73 ab | 118.63 a | 3.30 | <0.001 |
| NO (μmol/L) | 4.37 a | 0.26 | 4.10 ab | 3.87 b | 0.26 | <0.001 |
| Uric acid (mg/dL) | 11.70 | 0.11 | 11.20 | 11.43 | 0.11 | 0.060 |
| Creatinine (mg/dL) | 0.63 | 0.05 | 0.58 | 0.57 | 0.05 | 0.030 |
| ALT (U/L) | 11.67 | 0.05 | 11.87 | 11.67 | 0.05 | 0.090 |
| AST (U/L) | 56.67 | 5.31 | 56.23 | 57.53 | 5.31 | 0.120 |
| Cholesterol (mg/dL) | 149.17 a | 2.63 | 145.60 a | 140.57 b | 2.63 | 0.020 |
| Triglycerides (mg/dL) | 89.23 a | 4.28 | 88.20 ab | 81.63 b | 4.28 | <0.001 |
| HDL-c (mg/dL) | 34.33 b | 1.82 | 37.53 ab | 38.07 a | 1.82 | 0.030 |
| LDL-c (mg/dL) | 80.90 a | 2.04 | 74.49 a | 66.44 b | 2.04 | 0.060 |
| Item | Control | FBEs5% | FBEs10% | FBEs15% | SEM | p-Value |
|---|---|---|---|---|---|---|
| Intestinal tissues | ||||||
| GSH-Px (U/mg protein) | 144.10 b | 151.01 b | 150.47 b | 163.01 a | 1.12 | <0.001 |
| SOD (U/mg protein) | 20.27 b | 21.03 b | 22.06 ab | 23.80 a | 1.06 | 0.010 |
| Catalase (U/mg protein) | 15.80 c | 18.20 c | 25.13 b | 25.08 a | 0.93 | <0.001 |
| T-AOC (U/mg protein) | 1.59 c | 1.56 c | 1.71 b | 1.83 a | 0.26 | <0.001 |
| ROS (μL/g tissue) | 6.31 a | 5.50 b | 5.10 bc | 4.86 c | 0.39 | 0.030 |
| H2O2 (μmoL/g tissue) | 3.96 a | 3.60 b | 3.41 bc | 3.08 c | 0.11 | <0.001 |
| Malondialdehyde (nmoL/mL) | 6.50 a | 6.07 a | 5.21 b | 4.45 c | 0.22 | 0.030 |
| Breast muscle | ||||||
| GSH-Px (U/mg protein) | 152.83 d | 155.03 c | 160.07 b | 168.33 a | 0.86 | 0.030 |
| SOD (U/mg protein) | 36.40 d | 50.40 c | 59.43 b | 66.07 a | 0.12 | <0.001 |
| Catalase (U/mg protein) | 10.17 d | 12.87 c | 13.97 b | 15.90 a | 0.21 | 0.030 |
| T-AOC (U/mg protein) | 1.80 b | 2.13 ab | 2.53 a | 2.50 a | 0.11 | 0.010 |
| ROS | 7.86 a | 5.63 b | 4.99 bc | 4.20 c | 0.09 | 0.230 |
| H2O2 (μmoL/g tissue) | 3.89 a | 3.72 b | 3.12 c | 3.10 c | 1.20 | 0.120 |
| Malondialdehyde (nmoL/mL) | 7.50 a | 7.30 ab | 6.10 ab | 5.37 b | 0.63 | 0.290 |
| Item | Control | FBEs5% | FBEs10% | FBEs15% | SEM | p-Value |
|---|---|---|---|---|---|---|
| Intestinal tissues | ||||||
| Villus height duodenum, µm | 1166 c | 1229 c | 1245 a | 1233 a | 1.12 | <0.001 |
| Crypt depth duodenum, µm | 254.33 d | 215.32 c | 182.67 b | 195.00 a | 1.06 | 0.010 |
| Villus height: Crypt depth | 4.58 d | 5.71 c | 6.82 b | 6.33 a | 0.93 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, D.; El-sayed, H.I.; Mahmoud, E.R.; El-Rahman, G.I.A.; Bazeed, S.M.; Abdelwarith, A.A.; Elgamal, A.; Khalil, S.S.; Younis, E.M.; Kishawy, A.T.Y.; et al. Impacts of Solid-State Fermented Barley with Fibrolytic Exogenous Enzymes on Feed Utilization, and Antioxidant Status of Broiler Chickens. Vet. Sci. 2023, 10, 594. https://doi.org/10.3390/vetsci10100594
Ibrahim D, El-sayed HI, Mahmoud ER, El-Rahman GIA, Bazeed SM, Abdelwarith AA, Elgamal A, Khalil SS, Younis EM, Kishawy ATY, et al. Impacts of Solid-State Fermented Barley with Fibrolytic Exogenous Enzymes on Feed Utilization, and Antioxidant Status of Broiler Chickens. Veterinary Sciences. 2023; 10(10):594. https://doi.org/10.3390/vetsci10100594
Chicago/Turabian StyleIbrahim, Doaa, Hassainen I. El-sayed, Elsabbagh R. Mahmoud, Ghada I. Abd El-Rahman, Shefaa M. Bazeed, Abdelwahab A. Abdelwarith, Aya Elgamal, Samah S. Khalil, Elsayed M. Younis, Asmaa T. Y. Kishawy, and et al. 2023. "Impacts of Solid-State Fermented Barley with Fibrolytic Exogenous Enzymes on Feed Utilization, and Antioxidant Status of Broiler Chickens" Veterinary Sciences 10, no. 10: 594. https://doi.org/10.3390/vetsci10100594