
Mitigating Grapevine Red Blotch Virus Impact on Final Wine Composition

 ,
, 
Abstract
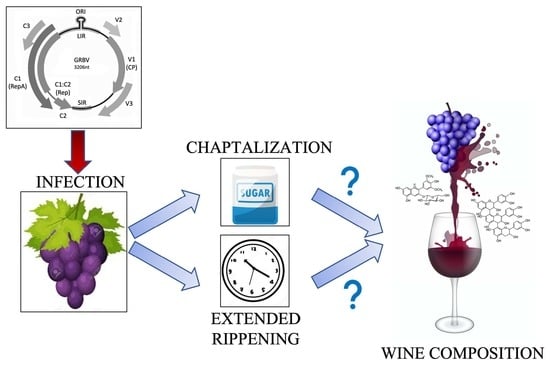
:
1. Introduction
2. Materials and Methods
2.1. Grape Harvest and Winemaking
2.2. Phenolic Analysis
2.2.1. Phenolic Extraction through Fermentation
2.2.2. Wine Phenolic Analysis
2.3. Volatile Profile Analysis
2.4. Sensory Evaluation
2.5. Statistical Analysis
3. Results
3.1. Basic Grape Chemical Composition at Harvest
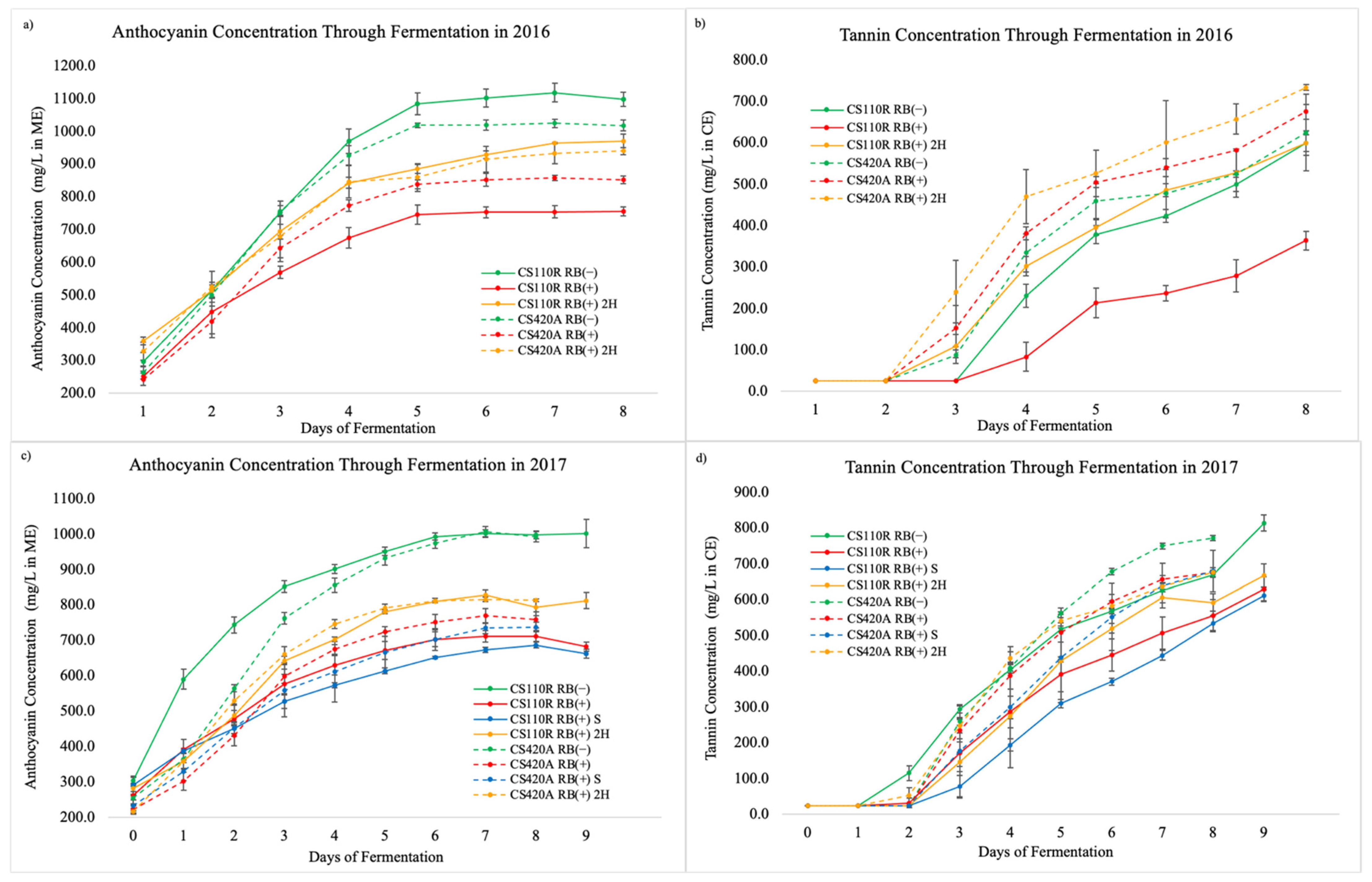
3.2. Phenolic Extractability
3.3. Final Wine Composition
3.3.1. Chemical Parameters at Bottling
3.3.2. Phenolic Compound Composition
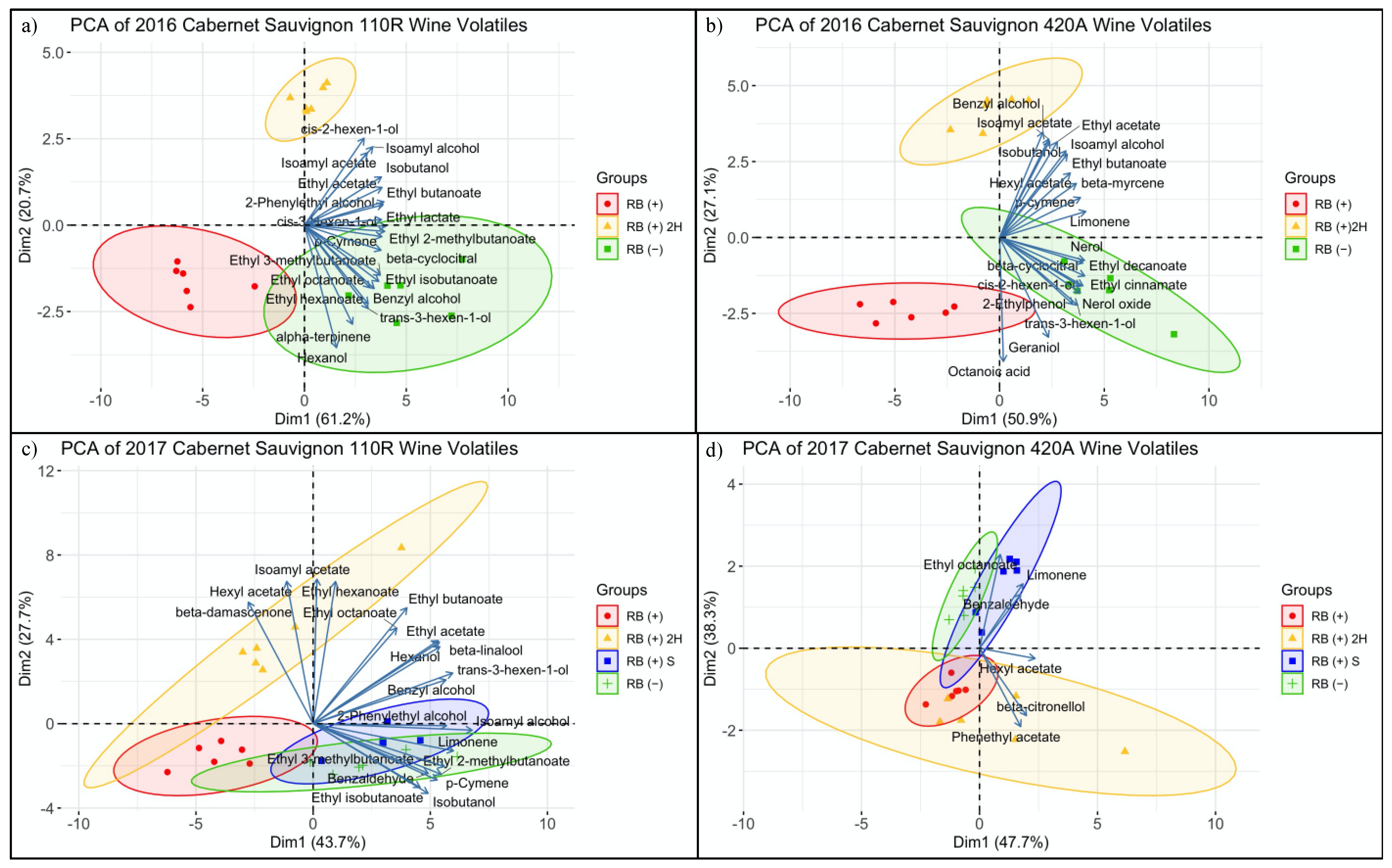
3.3.3. Volatile Compound Composition
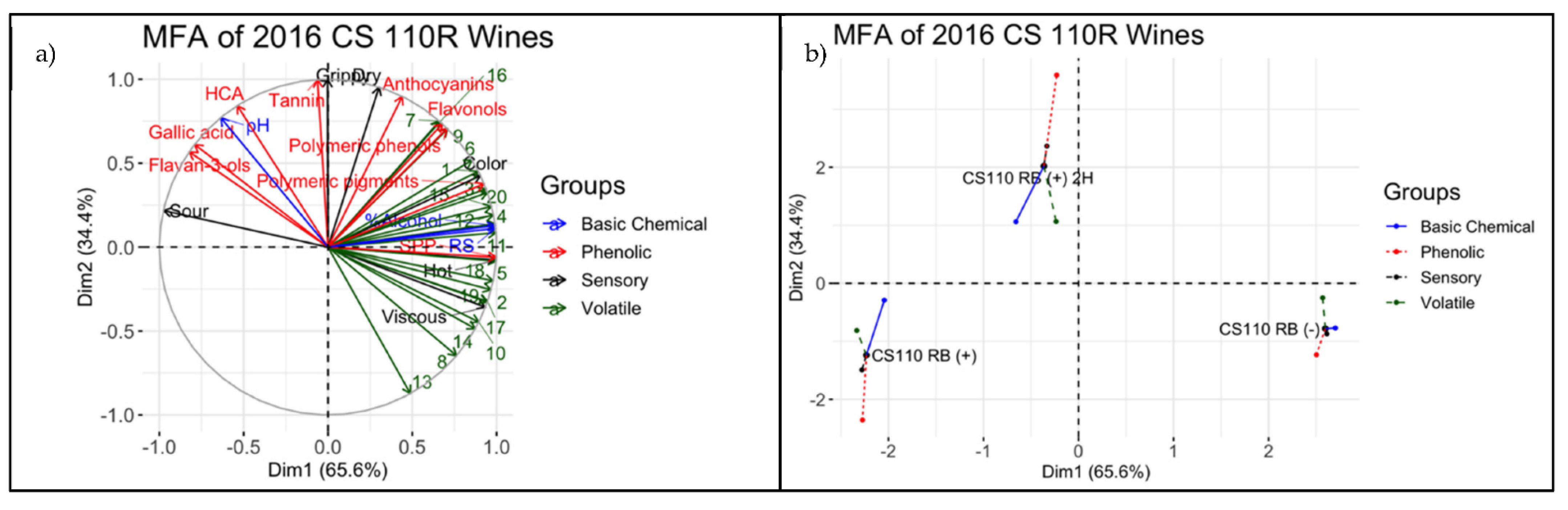
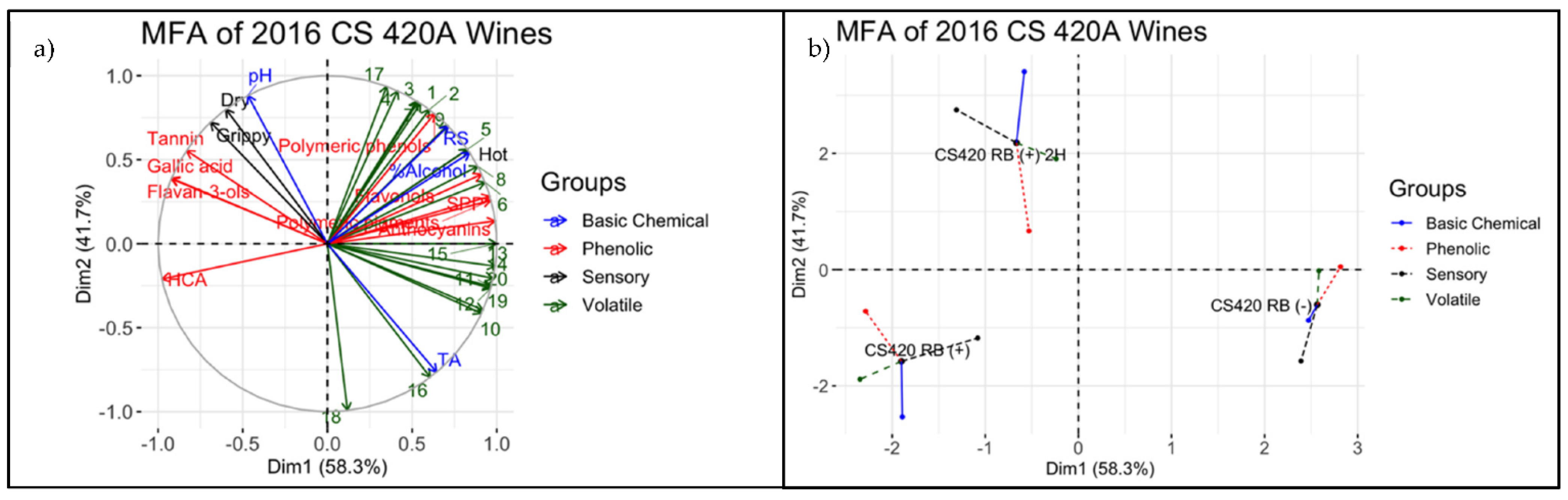
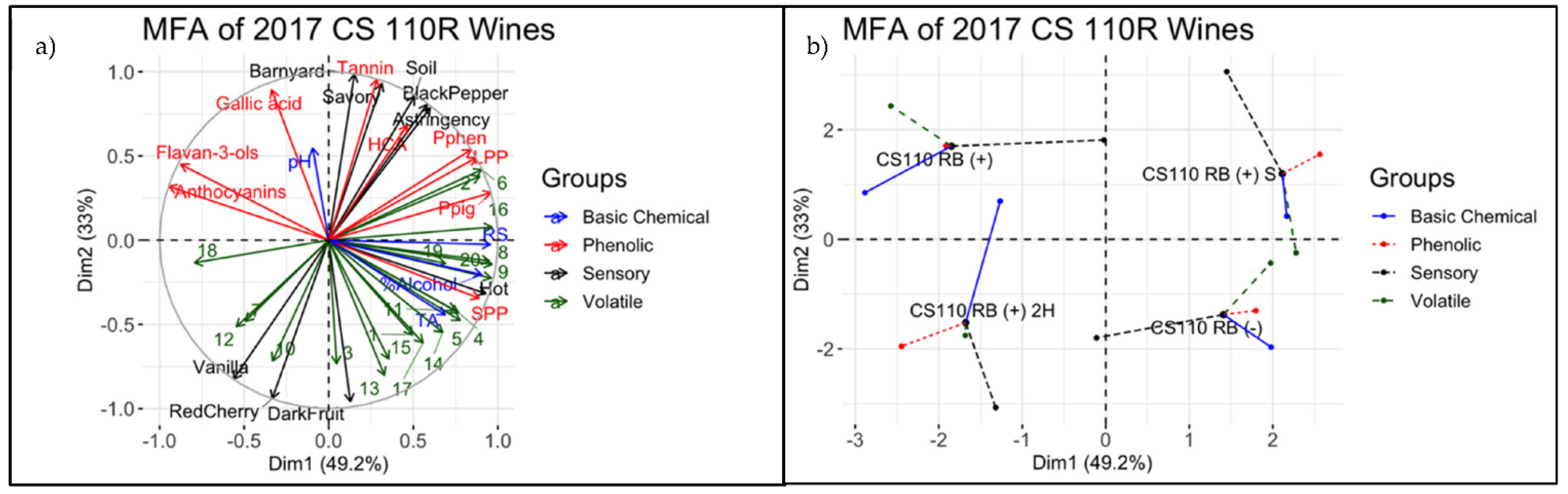
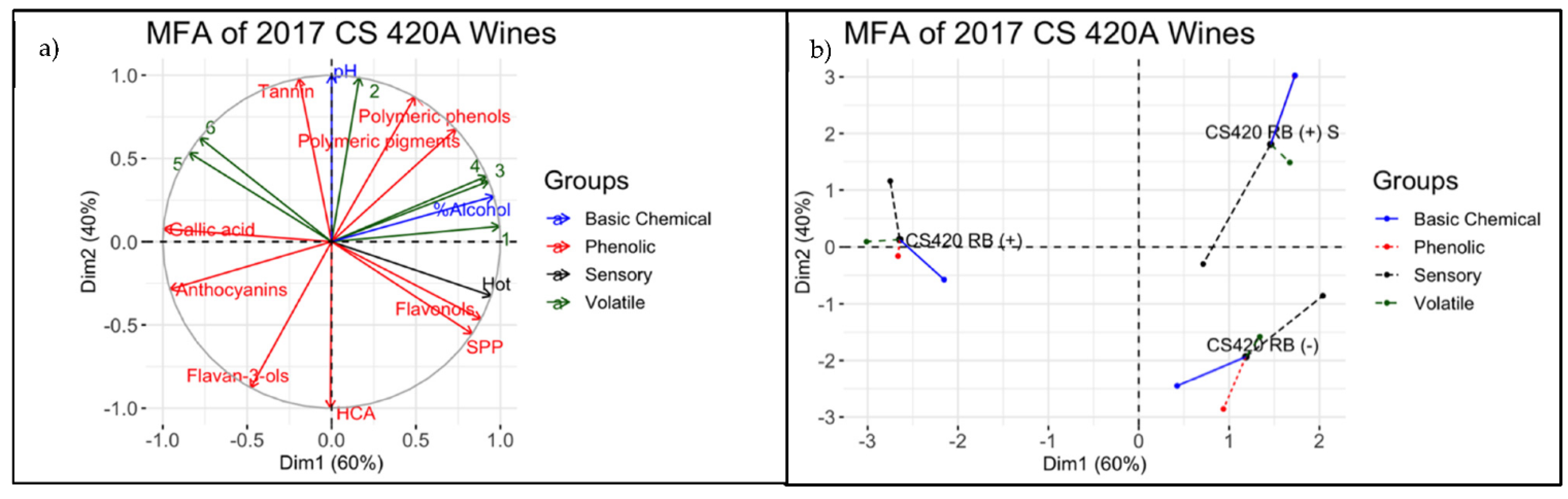
3.4. Descriptive Analysis of Final Wines
4. Discussion
4.1. Phenolic Extractability
4.2. The Effect of Ethanol and Ripeness Stage on Wine Chemical Composition
4.3. Integrating Chemical and Sensorial Observations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Ethics Statement
References
- Dolja, V.V.; Meng, B.; Martelli, G.P. Evolutionary Aspects of Grapevine Virology. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer International Publishing: New York, NY, USA, 2017; pp. 659–688. ISBN 978-3-319-57706-7. [Google Scholar]
- Krenz, B.; Thompson, J.R.; Fuchs, M.; Perry, K.L. Complete Genome Sequence of a New Circular DNA Virus from Grapevine. J. Virol. 2012, 86, 7715. [Google Scholar] [CrossRef] [Green Version]
- Rwahnih, M.A.; Dave, A.; Anderson, M.M.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Association of a DNA Virus with Grapevines Affected by Red Blotch Disease in California. Phytopathology 2013, 103, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Krenz, B.; Thompson, J.R.; Mclane, H.L.; Fuchs, M.; Perry, K.L. Grapevine red blotch-associated virus is widespread in the United States. Phytopathology 2014, 104, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Poojari, S.; Lowery, D.T.; Rott, M.; Schmidt, A.M.; Úrbez-Torres, J.R. Incidence, distribution and genetic diversity of Grapevine red blotch virus in British Columbia. Can. J. Plant Pathol. 2017, 39, 201–211. [Google Scholar] [CrossRef]
- Reynard, J.; Gugerli, P. Effects of Grapevine red blotch-associated virus on vine physiology and fruit composition of field grown grapevine cv. Gamay. In Proceedings of the 18th Congress of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine (ICVG), Ankara, Turquie, 7–11 September 2015; pp. 234–235. [Google Scholar]
- Lim, S.; Igori, D.; Zhao, F.; Moon, J.; Cho, I.-S.; Choi, G.-S. First report of Grapevine red blotch-associated virus on grapevine in Korea. Plant Dis. 2016, 100, 1957. [Google Scholar] [CrossRef]
- Gasperin-Bulbarela, J.; Licea-Navarro, A.F.; Pino-Villar, C.; Hernandez-Martínez, R.; Carrillo-Tripp, J. First report of grapeviné red blotch virus in Mexico. Plant Dis. 2019, 103, 381. [Google Scholar] [CrossRef]
- Luna, F.; Debat, H.; Gomez-Talquenca, S.; Moyano, S.; Zavallo, D.; Asurmendi, S. First report of grapevine red blotch virus infecting grapevine in Argentina. J. Plant Pathol. 2019, 101, 1239. [Google Scholar] [CrossRef] [Green Version]
- Marwal, A.; Kumar, R.; Paul Khurana, S.M.; Gaur, R.K. Complete nucleotide sequence of a new geminivirus isolated from Vitis vinifera in India: A symptomless host of Grapevine red blotch virus. Virusdisease 2019, 30, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Yepes, L.M.; Cieniewicz, E.; Krenz, B.; McLane, H.; Thompson, J.R.; Perry, K.L.; Fuchs, M. Causative Role of Grapevine Red Blotch Virus in Red Blotch Disease. Phytopathology 2018, 108, 902–909. [Google Scholar] [CrossRef] [Green Version]
- Calvi, B.L. Effects of Red-leaf Disease on Cabernet Sauvignon at the Oakville Experimental Vineyard and Mitigation by Harvest Delay and Crop Adjustment; University of California, Davis: Davis, CA, USA, 2011. [Google Scholar]
- Sudarshana, M.R.; Perry, K.L.; Fuchs, M.F. Grapevine Red Blotch-Associated Virus, an Emerging Threat to the Grapevine Industry Mysore. Phytopathology 2015, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girardello, R.C.; Cooper, M.L.; Smith, R.J.; Lerno, L.A.; Bruce, R.C.; Eridon, S.; Oberholster, A. Impact of Grapevine Red Blotch Disease on Grape Composition of Vitis vinifera Cabernet Sauvignon, Merlot, and Chardonnay. J. Agric. Food Chem. 2019, 67, 5496–5511. [Google Scholar] [CrossRef]
- Martínez-Lüscher, J.; Plank, C.M.; Brillante, L.; Cooper, M.L.; Smith, R.J.; Al-Rwahnih, M.; Yu, R.; Oberholster, A.; Girardello, R.; Kurtural, S.K. Grapevine Red Blotch Virus May Reduce Carbon Translocation Leading to Impaired Grape Berry Ripening. J. Agric. Food Chem. 2019, 67, 2437–2448. [Google Scholar] [CrossRef]
- Wallis, C.M.; Sudarshana, M.R.; Girardello, R.C.; Cooper, M.L.; Smith, R.J.; Lerno, L.A.; Bruce, R.C.; Eridon, S.; Oberholster, A. Effects of Grapevine red blotch-associated virus (GRBaV) infection on foliar metabolism of grapevines. Can. J. Plant Pathol. 2016, 38, 5496–5511. [Google Scholar] [CrossRef]
- Rumbaugh, A.C.; Girardello, R.C.; Cooper, M.L.; Plank, C.M.; Kurtural, S.K.; Oberholster, A. Impact of Rootstock and Season on Red Blotch Disease Expression in Cabernet Sauvignon (V. vinifera). Plants 2021, 10, 1583. [Google Scholar] [CrossRef] [PubMed]
- Girardello, R.C.; Cooper, M.L.; Lerno, L.A.; Brenneman, C.; Eridon, S.; Sokolowsky, M.; Heymann, H.; Oberholster, A. Impact of Grapevine Red Blotch Disease on Cabernet Sauvignon and Merlot Wine Composition and Sensory Attributes. Molecules 2020, 25, 3299. [Google Scholar] [CrossRef]
- Bowen, P.; Bogdanoff, C.; Poojari, S.; Usher, K.; Lowery, T.; Úrbez-Torres, J.R. Effects of grapevine red blotch disease on cabernet franc vine physiology, bud hardiness, and fruit and wine quality. Am. J. Enol. Vitic. 2020, 71, 308–318. [Google Scholar] [CrossRef]
- Coombe, B.G.; Mccarthy, M.G. Dynam ics of grape berry grow th and physiology of ripening. Aust. J. Grape Wine Res. 2000, 6, 131–135. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines: Anatomy and Physiology, 2nd ed.; Elsevier Science: San Diego, CA, USA, 2015. [Google Scholar]
- Bondada, B.; Harbertson, E.; Shrestha, P.M.; Keller, M. Temporal extension of ripening beyond its physiological limits imposes physical and osmotic challenges perturbing metabolism in grape (Vitis vinifera L.) berries. Sci. Hortic. (Amsterdam) 2017, 219, 135–143. [Google Scholar] [CrossRef]
- Coombe, B.G. The Grape Berry as a Sink. Acta Hortic. 1989, 239, 149–158. [Google Scholar] [CrossRef]
- Casassa, L.F.; Beaver, C.W.; Mireles, M.; Larsen, R.C.; Hopfer, H.; Heymann, H.; Harbertson, J.F. Influence of fruit maturity, maceration length, and ethanol amount on chemical and sensory properties of Merlot wines. Am. J. Enol. Vitic. 2013, 64, 437–449. [Google Scholar] [CrossRef]
- Bautista-Ortín, A.B.; Fernández-Fernández, J.I.; López-Roca, J.M.; Gómez-Plaza, E. The effect of grape ripening stage on red wine color. J. Int. Sci. Vigne Vin 2006, 40, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Canuti, V.; Conversano, M.; Calzi, M.L.; Heymann, H.; Matthews, M.A.; Ebeler, S.E. Headspace solid-phase microextraction-gas chromatography-mass spectrometry for profiling free volatile compounds in Cabernet Sauvignon grapes and wines. J. Chromatogr. A 2009, 1216, 3012–3022. [Google Scholar] [CrossRef]
- Kalua, C.M.; Boss, P.K. Evolution of volatile compounds during the development of cabernet sauvignon grapes (Vitis vinifera L.). J. Agric. Food Chem. 2009, 57, 3818–3830. [Google Scholar] [CrossRef] [PubMed]
- Palomo, E.S.; Díaz-Maroto, M.C.; Viñas, M.A.G.; Soriano-Pérez, A.; Pérez-Coello, M.S. Aroma profile of wines from Albillo and Muscat grape varieties at different stages of ripening. Food Control 2007, 18, 398–403. [Google Scholar] [CrossRef]
- Bindon, K.; Holt, H.; Williamson, P.O.; Varela, C.; Herderich, M.; Francis, I.L. Relationships between harvest time and wine composition in Vitis vinifera L. cv. Cabernet Sauvignon 2. Wine sensory properties and consumer preference. Food Chem. 2014, 154, 90–101. [Google Scholar] [CrossRef]
- Lerno, L.; Reichwage, M.; Panprivech, S.; Ponangi, R.; Hearne, L.; Oberholster, A.; Block, D.E. Chemical gradients in pilot-scale cabernet sauvignon fermentations and their effect on phenolic extraction. Am. J. Enol. Vitic. 2017, 68, 401–411. [Google Scholar] [CrossRef]
- Bindon, K.A.; Madani, S.H.; Pendleton, P.; Smith, P.A.; Kennedy, J.A. Factors affecting skin tannin extractability in ripening grapes. J. Agric. Food Chem. 2014, 62, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
- Medina-Plaza, C.; Beaver, J.W.; Lerno, L.; Dokoozlian, N.; Ponangi, R.; Blair, T.; Block, D.E.; Oberholster, A. Impact of temperature, ethanol and cell wall material composition on cell wall-anthocyanin interactions. Molecules 2019, 24, 3350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassara, S.; Kennedy, J.A. Relationship between red wine grade and phenolics. 2. Tannin composition and size. J. Agric. Food Chem. 2011, 59, 8409–8412. [Google Scholar] [CrossRef] [PubMed]
- Sherman, E.; Greenwood, D.R.; Villas-Boâs, S.G.; Heymann, H.; Harbertson, J.F. Impact of grape maturity and ethanol concentration on sensory properties of Washington State merlot wines. Am. J. Enol. Vitic. 2017, 68, 344–356. [Google Scholar] [CrossRef]
- Costello, P.J.; Francis, I.L.; Bartowsky, E.J. Variations in the effect of malolactic fermentation on the chemical and sensory properties of cabernet sauvignon wine: Interactive influences of oenococcus oeni strain and wine matrix composition. Aust. J. Grape Wine Res. 2012, 18, 287–301. [Google Scholar] [CrossRef]
- López, R.; López-Alfaro, I.; Gutiérrez, A.R.; Tenorio, C.; Garijo, P.; González-Arenzana, L.; Santamaría, P. Malolactic fermentation of Tempranillo wine: Contribution of the lactic acid bacteria inoculation to sensory quality and chemical composition. Int. J. Food Sci. Technol. 2011, 46, 2373–2381. [Google Scholar] [CrossRef]
- Iland, P.; Bruer, N.; Edwards, G.; Weeks, S.; Wilkes, E. Chemical Analysis of Grapes and Wine: Techniques and Concepts; Patrick Iland Wine Promotions PTY LTD: Campbelltown SA, Australia, 2004. [Google Scholar]
- Skogerson, K.; Downey, M.; Mazza, M.; Boulton, R. Rapid determination of phenolic components in red wines from UV-visible spectra and the method of partial least squares. Am. J. Enol. Vitic. 2007, 58, 318–325. [Google Scholar]
- Harbertson, J.F.; Kennedy, J.A.; Adams, D.O. Tannin in skins and seeds of Cabernet Sauvignon, Syrah, and Pinot noir berries during ripening. Am. J. Enol. Vitic. 2002, 53, 54–59. [Google Scholar]
- Harbertson, J.F.; Picciotto, E.A.; Adams, D.O. Measurement of Polymeric Pigments in Grape Berry Extract sand Wines Using a Protein Precipitation Assay Combined with Bisulfite Bleaching. Am. J. Enol. Vitic. 2003, 54, 301–306. [Google Scholar]
- Harbertson, J.F.; Mireles, M.; Yu, Y. Improvement of BSA tannin precipitation assay by reformulation of resuspension buffer. Am. J. Enol. Vitic. 2015, 66, 95–99. [Google Scholar] [CrossRef]
- Peng, Z.; Iland, P.G.; Oberholster, A.; Sefton, M.A.; Waters, E.J. Analysis of pigmented polymers in red wine by reverse phase HPLC. Aust. J. Grape Wine Res. 2002, 8, 70–75. [Google Scholar] [CrossRef]
- Hendrickson, D.A.; Lerno, L.A.; Hjelmeland, A.K.; Ebeler, S.E.; Heymann, H.; Hopfer, H.; Block, K.L.; Brenneman, C.A.; Oberholster, A. Impact of mechanical harvesting and optical berry sorting on grape and wine composition. Am. J. Enol. Vitic. 2016, 67, 385–397. [Google Scholar] [CrossRef]
- Lawless, H.; Heymann, H. Sensory Evaluation of Food: Principles and Practices; Springer: New York, NY, USA, 1998. [Google Scholar]
- STHDA. Practical Guide to Principal Component Methods in R. 2017. Available online: http://www.sthda.com/english/wiki/practical-guide-to-principal-component-methods-in-r (accessed on 26 November 2021).
- Marquez, A.; Perez-Serratosa, M.; Varo, M.A.; Merida, J. Effect of temperature on the anthocyanin extraction and color evolution during controlled dehydration of tempranillo grapes. J. Agric. Food Chem. 2014, 62, 7897–7902. [Google Scholar] [CrossRef]
- Hernández-Hierro, J.M.; Quijada-Morín, N.; Martínez-Lapuente, L.; Guadalupe, Z.; Ayestarán, B.; Rivas-Gonzalo, J.C.; Escribano-Bailón, M.T. Relationship between skin cell wall composition and anthocyanin extractability of Vitis vinifera L. cv. Tempranillo at different grape ripeness degree. Food Chem. 2014, 146, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Bindon, K.A.; Bacic, A.; Kennedy, J.A. Tissue-specific and developmental modifications of grape cell walls influence the adsorption of proanthocyanidins. J. Agric. Food Chem. 2012, 60, 9249–9260. [Google Scholar] [CrossRef]
- Beaver, J.W.; Medina-Plaza, C.; Miller, K.; Dokoozlian, N.; Ponangi, R.; Blair, T.; Block, D.; Oberholster, A. Effects of the Temperature and Ethanol on the Kinetics of Proanthocyanidin Adsorption in Model Wine Systems. J. Agric. Food Chem. 2019, 68, 2891–2899. [Google Scholar] [CrossRef] [PubMed]
- Springer, L.F.; Sacks, G.L. Protein-precipitable tannin in wines from Vitis vinifera and interspecific hybrid grapes (Vitis ssp.): Differences in concentration, extractability, and cell wall binding. J. Agric. Food Chem. 2014, 62, 7515–7523. [Google Scholar] [CrossRef]
- Bautista-Ortín, A.B.; Molero, N.; Marín, F.; Ruiz-García, Y.; Gómez-Plaza, E. Reactivity of pure and commercial grape skin tannins with cell wall material. Eur. Food Res. Technol. 2014, 240, 645–654. [Google Scholar] [CrossRef]
- Bautista-Ortín, A.B.; Rodríguez-Rodríguez, P.; Gil-Muñoz, R.; Jiménez-Pascual, E.; Busse-Valverde, N.; Martínez-Cutillas, A.; López-Roca, J.M.; Gómez-Plaza, E. Influence of berry ripeness on concentration, qualitative composition and extractability of grape seed tannins. Aust. J. Grape Wine Res. 2012, 18, 123–130. [Google Scholar] [CrossRef]
- Pedneault, K.; Dorais, M.; Angers, P. Flavor of cold-hardy grapes: Impact of berry maturity and environmental conditions. J. Agric. Food Chem. 2013, 61, 10418–10438. [Google Scholar] [CrossRef] [PubMed]
- Deluc, L.G.; Grimplet, J.; Wheatley, M.D.; Tillett, R.L.; Quilici, D.R.; Osborne, C.; Schooley, D.A.; Schlauch, K.A.; Cushman, J.C.; Cramer, G.R.; et al. Transcriptomic and metabolite analyses of Cabernet Sauvignon grape berry development. BMC Genomics 2013, 61, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldner, M.C.; Zamora, M.C.; Lira, P.D.L.; Gianninoto, H.; Bandoni, A. Effect of ethanol level in the perception of aroma attributes and the detection of volatile compounds in red wine. J. Sens. Stud. 2009, 24, 243–257. [Google Scholar] [CrossRef]
- Robinson, A.L.; Ebeler, S.E.; Heymann, H.; Boss, P.K.; Solomon, P.S.; Trengove, R.D. Interactions between wine volatile compounds and grape and wine matrix components influence aroma compound headspace partitioning. J. Agric. Food Chem. 2009, 57, 10313–10322. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.L.; Sacks, G.L.; Jeffery, D.W. Understanding Wine Chemistry; John Wiley & Sons, Ltd.: West Sussex, UK, 2016; ISBN 9781118627808. [Google Scholar]
- Cameleyre, M.; Lytra, G.; Tempere, S.; Barbe, J.C. Olfactory Impact of Higher Alcohols on Red Wine Fruity Ester Aroma Expression in Model Solution. J. Agric. Food Chem. 2015, 63, 9777–9788. [Google Scholar] [CrossRef]
- Josse, J.; Pagès, J.; Husson, F. Testing the significance of the RV coefficient. Comput. Stat. Data Anal. 2008, 53, 82–91. [Google Scholar] [CrossRef]
- Heymann, H.; Licalzi, M.; Conversano, M.R.; Bauer, A.; Skogerson, K.; Matthews, M. Effects of extended grape ripening with or without must and wine alcohol manipulations on cabernet sauvignon wine sensory characteristics. S. Afr. J. Enol. Vitic. 2013, 34, 86–99. [Google Scholar] [CrossRef]
- Escudero, A.; Campo, E.; Fariña, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007, 55, 4501–4510. [Google Scholar] [CrossRef]
- Villamor, R.R.; Evans, M.A.; Ross, C.F. Effects of ethanol, tannin, and fructose concentrations on sensory properties of model red wines. Am. J. Enol. Vitic. 2013, 64, 342–348. [Google Scholar] [CrossRef]
- De-La-Fuente-Blanco, A.; Sáenz-Navajas, M.P.; Ferreira, V. On the effects of higher alcohols on red wine aroma. Food Chem. 2016, 210, 107–114. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Harvest Date | °Brix | pH | TA (g/L) | YAN (mg/L) | Malic Acid (mg/L) |
|---|---|---|---|---|---|---|
| CS110 RB(−) | 20 September 2016 | 25.6 ± 0.1 a | 3.62 ± 0.0 a | 3.84 ± 0.3 b | 81.1 ± 7.1 b | 1460.0 ± 55.1 c |
| CS110 RB(+) | 20 September 2016 | 21.7 ± 0.1 c | 3.45 ± 0. b | 4.75 ± 0.1 a | 121.8 ± 9.8 a | 2275.0 ± 48.6 a |
| CS110 RB(+) 2H | 27 September 2016 | 23.8 ± 0.1 b | 3.59 ± 0.0 a | 4.49 ± 0.2 a | 127.2 ± 7.1 a | 1970.3 ± 29.5 b |
| CS420 RB(−) | 20 September 2016 | 24.3 ± 0.1 a | 3.50 ± 0.0 b | 4.23 ± 0.1 b | 99.7 ± 2.5 a | 1625.7 ± 48.0 c |
| CS420 RB(+) | 20 September 2016 | 22.1 ± 0.1 b | 3.48 ± 0.0 b | 4.53 ± 0.1 a | 83.6 ± 17.8 a | 1852.0 ± 13.9 b |
| CS420 RB(+) 2H | 27 September 2016 | 23.7 ± 0.1 a | 3.55 ± 0.0 a | 4.56 ± 0.2 a | 104.3 ± 3.2 a | 1953.3 ± 56.3 a |
| CS110 (–) | 26 September 2017 | 25.5 ± 0.1 b | 3.62 ± 0.0 b | 3.97 ± 0.0 c | 145.9 ±0.6 b | 2649.3 ± 45.7 b |
| CS110 (+) | 26 September 2017 | 23.4 ± 0.0 d | 3.57 ± 0.0 b | 4.87 ± 0.1 a | 150.2 ±1.8 b | 2779.0 ± 68.6 ab |
| CS110 (+) S | 26 September 2017 | 28.2 ± 0.5 a | 3.57 ± 0.1 b | 4.83 ± 0.1 a | 143.7 ±6.8 b | 2831.7 ± 140.4 ab |
| CS110 (+) 2H | 6 October 2017 | 24.7 ± 0.2 c | 3.86 ± 0.0 a | 4.18 ± 0.1 b | 164.0 ±1.4 a | 2971.7 ± 47.7 a |
| CS420 (–) | 6 October 2017 | 25.3 ± 0.1 a | 3.56 ± 0.0 b | 4.62 ± 0.2 a | 127.9 ±15.9 a | 2201.0 ± 34.7 c |
| CS420 (+) | 6 October 2017 | 23.6 ± 0.3 b | 3.51 ± 0.0 b | 4.82 ± 0.0 a | 106.3 ± 4.1 a | 2870.0 ± 21.0 a |
| CS420 (+) S | 6 October 2017 | 25.9 ± 0.6 a | 3.51 ± 0.0 b | 4.82 ± 0.1 a | 111.0 ± 13.5 a | 2823.7 ± 16.4 a |
| CS420 (+) 2H | 17 October 2017 | 24.2 ± 0.1 b | 3.70 ± 0.0 a | 4.05 ± 0.0 b | 117.1 ± 2.4 a | 2477.0 ± 39.0 b |
| 2016 | ||||||||
|---|---|---|---|---|---|---|---|---|
| 110R | 420A | |||||||
| Bottling Chemical Parameters | RB(−) | RB(+) | RB(+) 2H | RB(−) | RB(+) | RB(+) 2H | ||
| % Alcohol (% v/v) | 15.10 ± 0.20 a | 11.99 ± 0.24 c | 13.54 ± 0.09 b | 14.07 ± 0.18 a | 12.12 ± 0.18 c | 13.66 ± 0.09 b | ||
| pH | 3.55 ± 0.07 b | 3.6 ± 0.01 ab | 3.65 ± 0.03 a | 3.44 ± 0.04 b | 3.47 ± 0.01 b | 3.59 ± 0.05 a | ||
| TA (g/L) | 6.79 ± 0.21 a | 6.45 ± 0.12 a | 6.61 ± 0.13 a | 7.13 ± 0.21 a | 6.86 ± 0.04 ab | 6.51 ± 0.20 b | ||
| RS (g/L) | 0.27 ± 0.03 a | 0.17 ± 0.01 c | 0.22 ± 0.00 b | 0.20 ± 0.01 a | 0.19 ± 0.01 b | 0.20 ± 0.01 a | ||
| 2017 | ||||||||
| 110R | 420A | |||||||
| Bottling Chemical Parameters | RB(−) | RB(+) | RB(+) S | RB(+) 2H | RB(−) | RB(+) | RB(+) S | RB(+) 2H |
| % Alcohol (% v/v) | 15.42 ± 0.12 b | 13.77 ± 0.15 d | 16.06 ± 0.03 a | 14.72 ± 0.06 c | 15.01 ± 0.09 b | 14.02 ± 0.18 c | 15.51 ± 0.16 a | 14.30 ± 0.02 c |
| pH | 3.78 ± 0.05 b | 3.88 ± 0.01 a | 3.92 ± 0.04 a | 3.88 ± 0.03 ab | 3.58 ± 0.02 b | 3.67 ± 0.03 a | 3.71 ± 0.02 a | 3.70 ± 0.03 a |
| TA (g/L) | 6.74 ± 0.12 a | 5.83 ± 0.11 b | 6.06 ± 0.11 b | 5.94 ± 0.10 b | 6.41 ± 0.16 a | 6.05 ± 0.21 a | 6.42 ± 0.16 a | 6.21 ± 0.21 a |
| RS (g/L) | 0.74 ± 0.15 a | 0.23 ± 0.10 b | 0.96 ± 0.03 a | 0.42 ± 0.10 b | 0.87 ± 0.45 a | 0.47 ± 0.15 a | 0.61 ± 0.18 a | 0.24 ± 0.02 a |
| 2016 | ||||||
|---|---|---|---|---|---|---|
| 110R | 420A | |||||
| Phenolic Compound | RB(−) | RB(+) | RB(+) 2H | RB(−) | RB(+) | RB(+) 2H |
| Total Flavan-3-ols (mg/L) | 29.14 ± 0.60 b | 32.93 ± 0.95 a | 33.99 ± 0.96 a | 30.00 ± 0.77 b | 33.86 ± 0.65 a | 34.47 ± 0.29 a |
| Total HCA (mg/L) | 33.34 ± 0.50c | 35.57 ± 0.06 b | 38.58 ± 0.92a | 42.79 ± 2.84 b | 47.68 ± 3.74 a | 45.41 ± 1.64 ab |
| Total Flavonols (mg/L) | 70.64 ± 5.37 a | 47.66 ± 2.45 b | 75.74 ± 1.76 a | 82.45 ± 1.38 a | 73.00 ± 2.41 b | 79.16 ± 1.89 a |
| Total Anthocyanins (mg/L) | 334.42 ± 16.07 b | 295.61 ± 15.86 c | 365.64 ± 8.56 a | 370.96 ± 4.96 a | 329.98 ± 20.1 b | 346.31 ± 23.24 ab |
| Gallic Acid (mg/L) | 7.73 ± 0.28 c | 10.03 ± 0.20 b | 10.88 ± 0.44 a | 7.22 ± 0.37 b | 9.29 ± 0.06 a | 9.63 ± 0.47 a |
| Polymeric Pigments (mg/L) | 20.97 ± 4.12 a | 10.16 ± 2.07 b | 18.03 ± 1.90 a | 18.48 ± 0.22 a | 14.03 ± 0.06 c | 16.29 ± 1.44 b |
| Polymeric Phenols (mg/L) | 233.81 ± 44.59 a | 136.41 ± 24.48 b | 250.05 ± 32.61 a | 232.59 ± 10.83 a | 198.46 ± 9.55 b | 237.22 ± 22.76 a |
| SPP (Au520) | 2.34 ± 0.08 a | 1.30 ± 0.11 c | 1.64 ± 0.01 b | 1.60 ± 0.06 a | 1.15 ± 0.05 c | 1.39 ± 0.06 b |
| LPP (Au520) | 0.72 ± 0.21 a | 0.29 ± 0.18 a | 0.70 ± 0.12 a | 0.55 ± 0.21 a | 0.50 ± 0.06 a | 0.55 ± 0.05 a |
| Tannin (mg/L CE) | 173.53 ± 77.14 ab | 154.77 ± 19.76 b | 405.14 ± 87.81 a | 386.77 ± 41.76 a | 456.08 ± 26.96 a | 488.43 ± 41.87 a |
| 2017 | ||||||||
|---|---|---|---|---|---|---|---|---|
| 110R | 420A | |||||||
| Phenolic Compound | RB(−) | RB(+) | RB(+) S | RB(+) 2H | RB(−) | RB(+) | RB(+) S | RB(+) 2H |
| Total Flavan-3-ols (mg/L) | 37.98 ± 1.05 c | 46.45 ± 1.28 a | 37.93 ± 0.29 c | 42.26 ± 1.66 b | 41.94 ± 1.20 a | 42.14 ± 0.86 a | 37.65 ± 1.90 b | 42.01 ± 1.43 a |
| Total HCA (mg/L) | 26.89 ± 1.41 a | 27.39 ± 0.46 a | 26.22 ± 1.25 a | 17.17 ± 1.59 b | 28.99 ± 1.45 a | 24.03 ± 1.99 b | 21.55 ± 2.25 b | 16.40 ± 1.64 c |
| Total Flavonols (mg/L) | 36.51 ± 2.71 ab | 41.38 ± 2.29 a | 34.04 ± 2.38 b | 41.70 ± 7.43 a | 45.60 ± 3.27 ab | 47.36 ± 1.90 a | 44.14 ± 1.26 ab | 38.79 ± 4.47 b |
| Total Anthocyanins (mg/L) | 92.74 ± 26.48 b | 214.11 ± 11.70 a | 100.47 ± 28.10 b | 189.61 ± 29.54 a | 170.35 ± 11.66 a | 185.50 ± 5.43 a | 143.27 ± 28.5 b | 182.85 ± 16.75 a |
| Gallic Acid (mg/L) | 16.38 ± 0.17 c | 19.71 ± 0.86 a | 19.11 ± 0.11 a | 17.83 ± 0.17 b | 14.69 ± 0.19 b | 15.89 ± 0.39 a | 15.05 ± 0.69 b | 15.79 ± 0.44 a |
| Polymeric Pigments (mg/L) | 38.05 ± 11.50 a | 19.54 ± 0.65 b | 40.46 ± 4.45 a | 17.43 ± 6.63 b | 27.22 ± 5.40 ab | 22.17 ± 0.73 bc | 31.85 ± 2.64 a | 19.94 ± 5.57 c |
| Polymeric Phenols (mg/L) | 379.98 ± 98.00 a | 253.26 ± 2.36 b | 417.65 ± 25.17 a | 217.08 ± 81.58 b | 341.40 ± 38.37 ab | 298.81 ± 16.46 bc | 380.55 ± 31.61 a | 245.89 ± 68.18 c |
| SPP (Au520) | 3.14 ± 0.30 a | 1.50 ± 0.01 c | 2.53 ± 0.07 b | 1.74 ± 0.35 c | 2.49 ± 0.24 a | 1.47 ± 0.03 c | 2.00 ± 0.06 b | 2.00 ± 0.40 b |
| LPP (Au520) | 1.06 ± 0.72 ab | 0.54 ± 0.03 bc | 1.42 ± 0.12 a | 0.29 ± 0.28 c | 0.85 ± 0.25 ab | 0.65 ± 0.03 ab | 1.07 ± 0.09 a | 0.19 ± 0.32 b |
| Tannin (mg/L CE) | 297.99 ± 171.60 ab | 440.52 ± 33.49 a | 460.33 ± 25.76 a | 175.89 ± 133.11 b | 379.17 ± 61.19 b | 452.88 ± 37.89 ab | 542.14 ± 17.27 a | 230.01 ± 128.61 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rumbaugh, A.; Girardello, R.C.; Cantu, A.; Brenneman, C.; Heymann, H.; Oberholster, A. Mitigating Grapevine Red Blotch Virus Impact on Final Wine Composition. Beverages 2021, 7, 76. https://doi.org/10.3390/beverages7040076
Rumbaugh A, Girardello RC, Cantu A, Brenneman C, Heymann H, Oberholster A. Mitigating Grapevine Red Blotch Virus Impact on Final Wine Composition. Beverages. 2021; 7(4):76. https://doi.org/10.3390/beverages7040076
Chicago/Turabian StyleRumbaugh, Arran, Raul Cauduro Girardello, Annegret Cantu, Charles Brenneman, Hildegarde Heymann, and Anita Oberholster. 2021. "Mitigating Grapevine Red Blotch Virus Impact on Final Wine Composition" Beverages 7, no. 4: 76. https://doi.org/10.3390/beverages7040076