
Decellularized Human Umbilical Tissue-Derived Hydrogels Promote Proliferation and Chondrogenic Differentiation of Mesenchymal Stem Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval and Umbilical Cord Collection
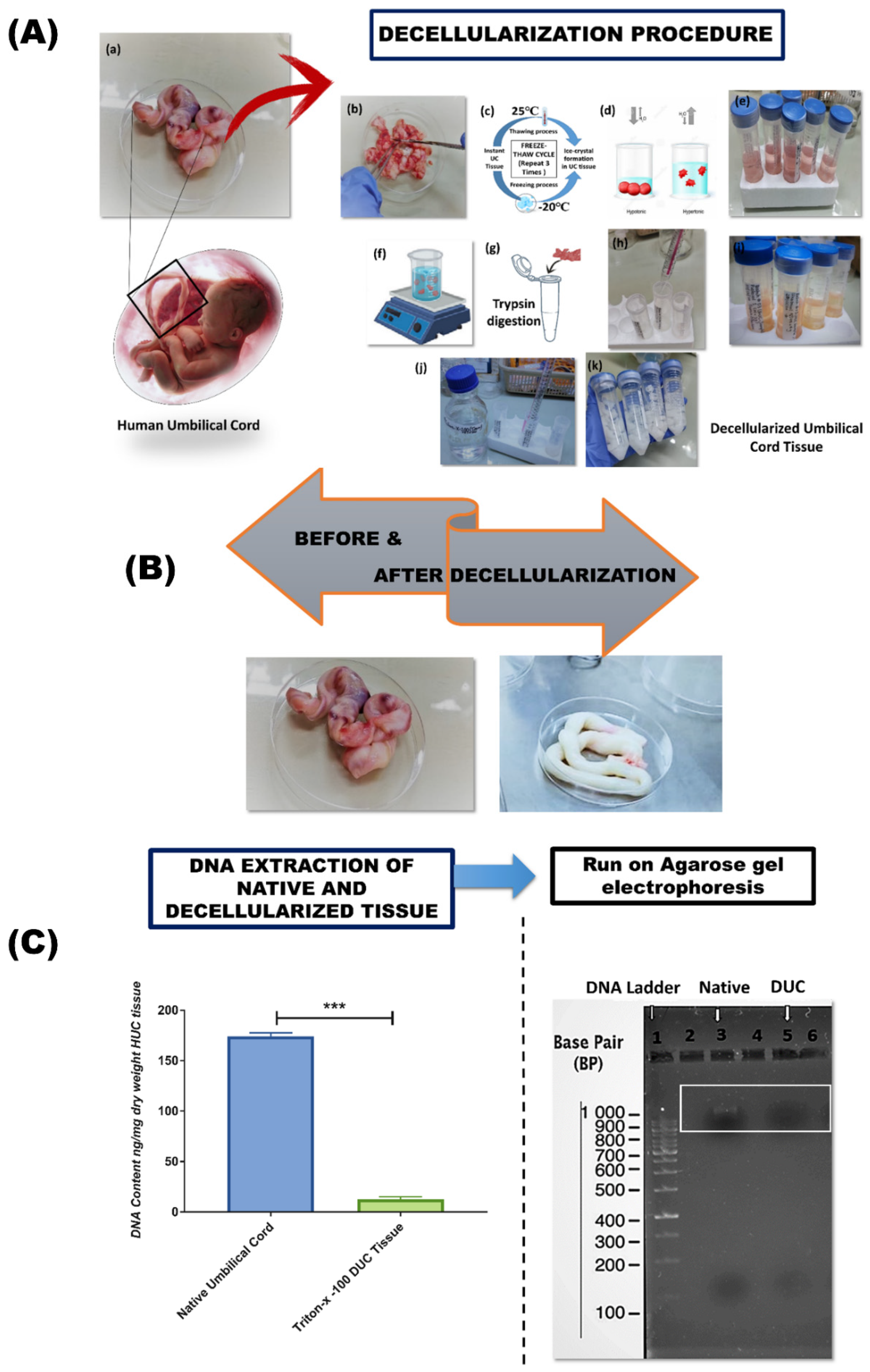
2.2. Preparation of Decellularized Human Umbilical Cord (DUC) Tissue
2.3. Confirmation of Decellularization
2.3.1. DNA Content Determination
2.3.2. Agarose Gel Electrophoresis
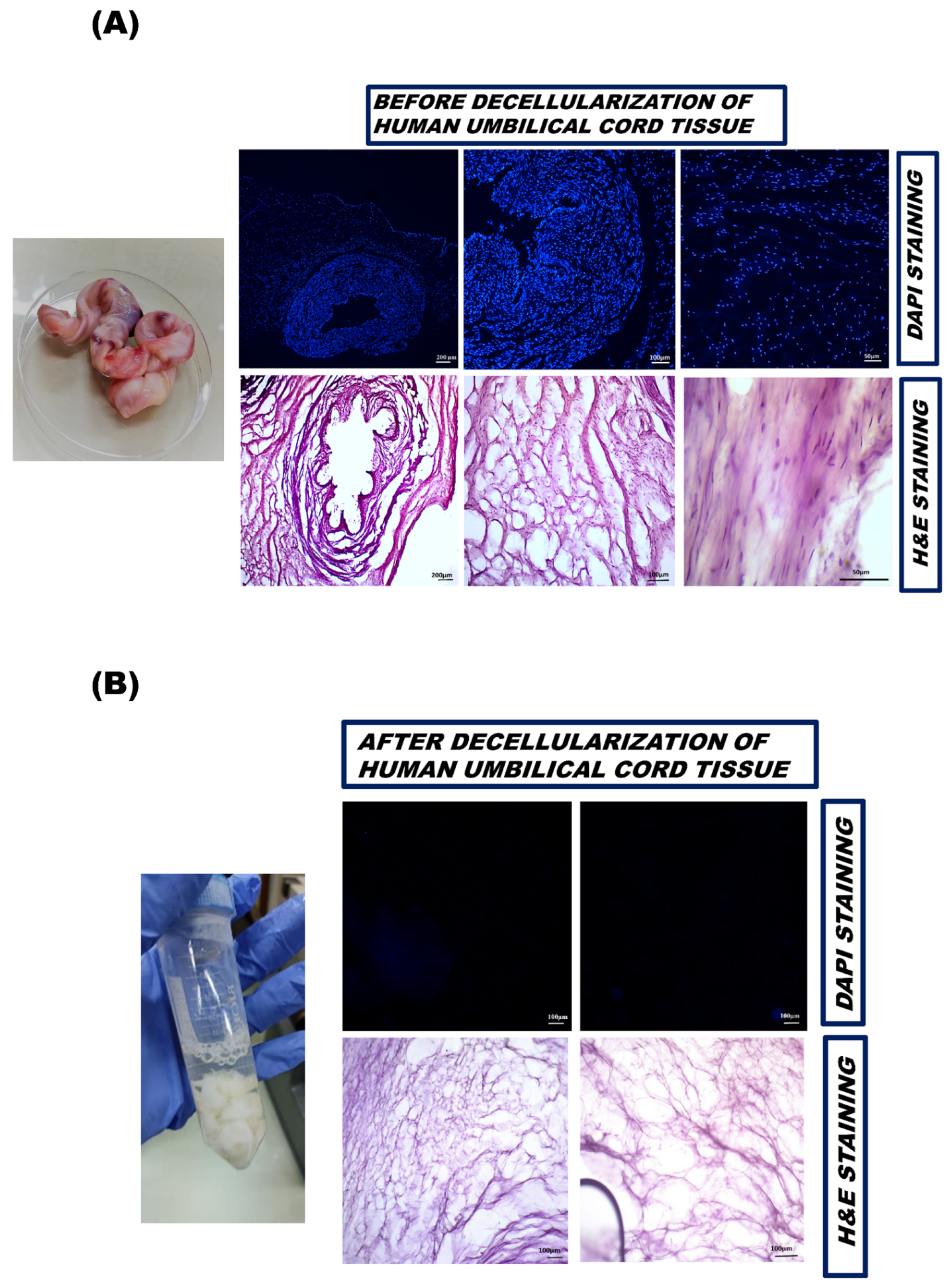
2.4. Histological Examination
2.4.1. Fixation, Embedding, and Cryosectioning of Harvested Tissue
2.4.2. DAPI Staining
2.4.3. Hematoxylin and Eosin Staining
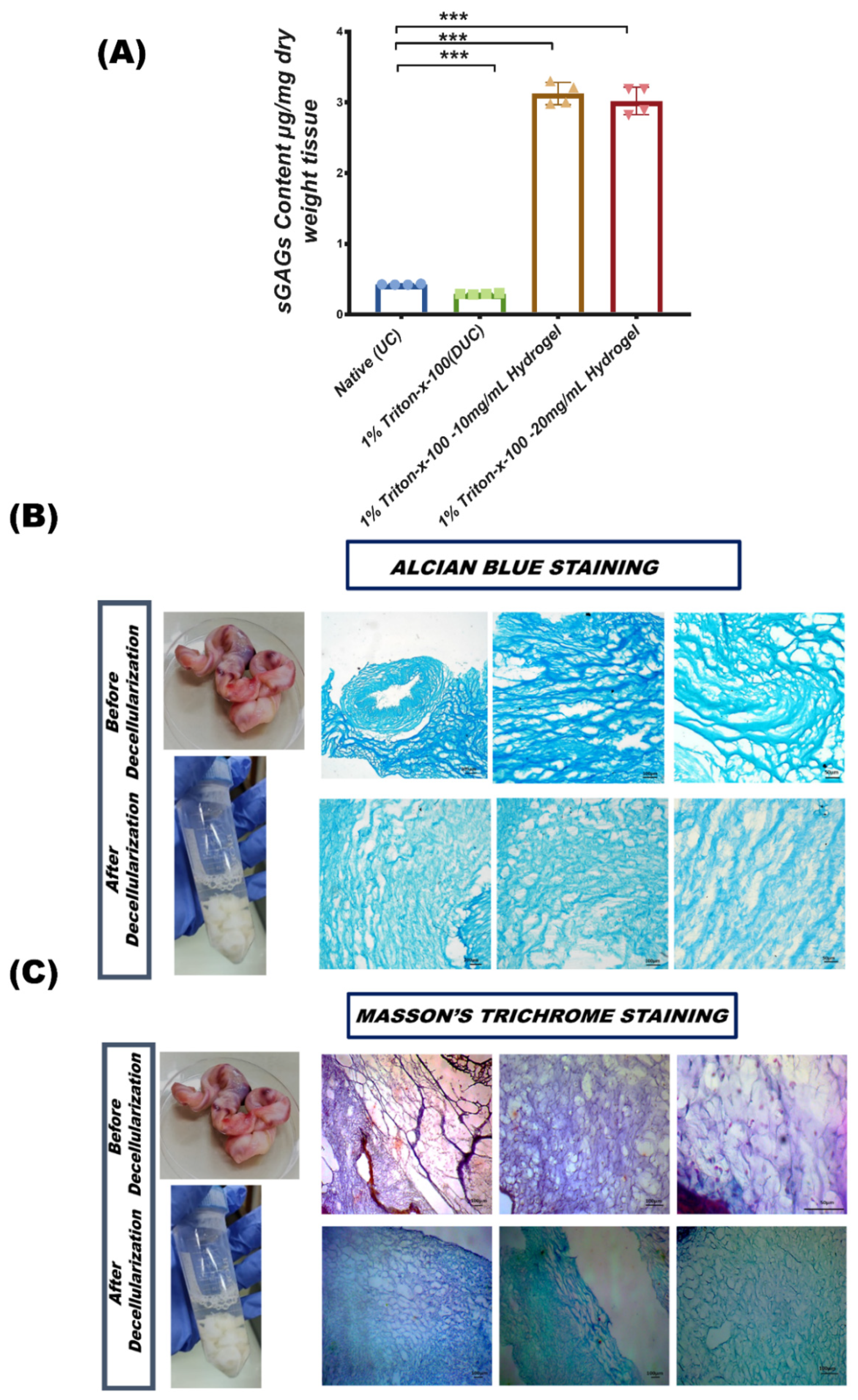
2.4.4. Quantification of GAG
2.4.5. Alcian Blue Staining
2.4.6. Masson’s Trichrome Staining
2.5. 3D Hydrogel Preparation from Decellularized Umbilical Cord Tissue
2.6. Hydrogel Characterization
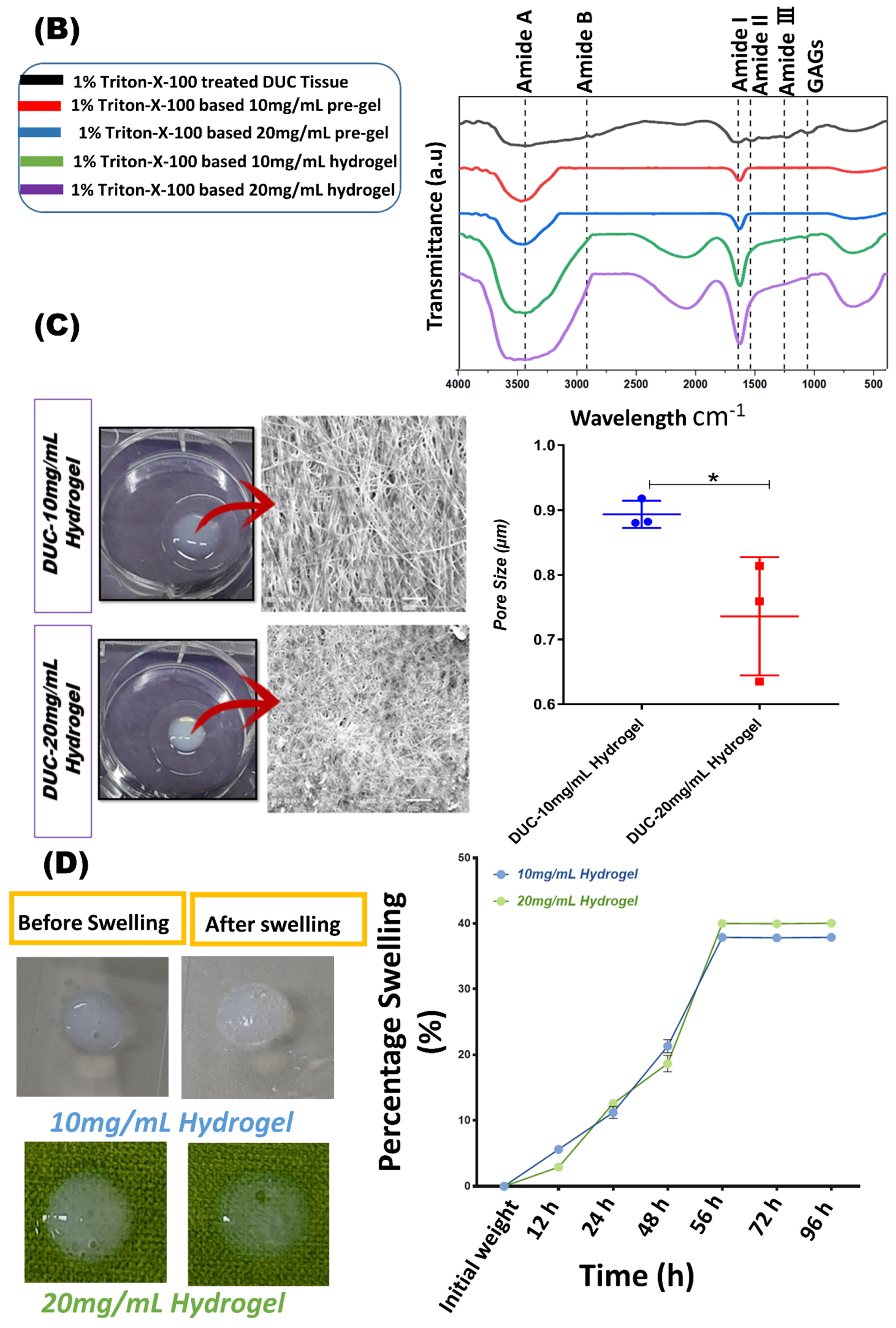
2.6.1. Material Composition by FTIR (Fourier-Transform Infrared) Spectroscopy
2.6.2. Scanning Electron Microscopy (SEM)
2.6.3. Swelling Behavior
2.7. Isolation and Characterization of Umbilical Cord-Derived MSCs
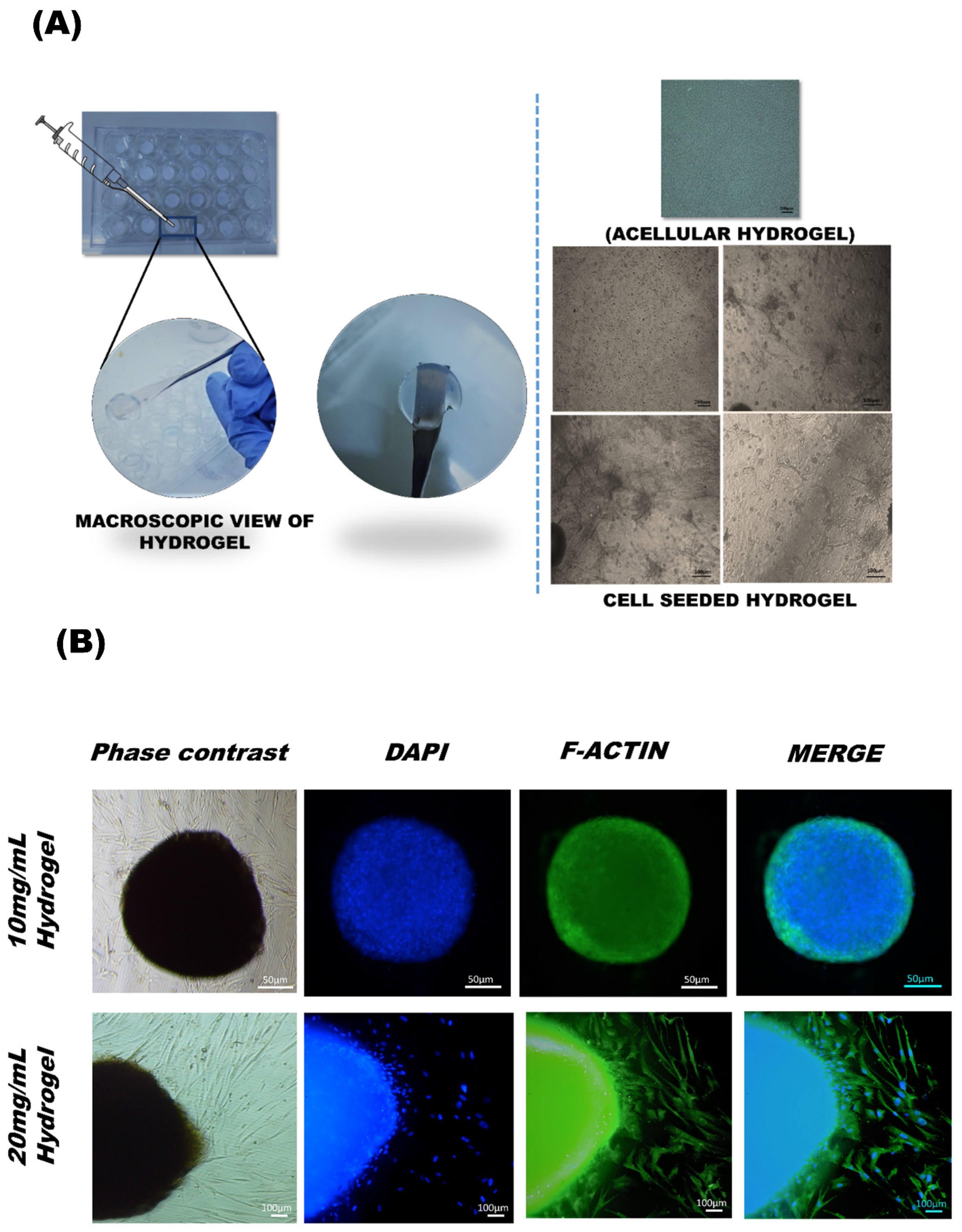
2.8. MSCs Cultured in Decellularized Umbilical Cord (DUC) Hydrogel
F-Actin Staining
2.9. Assessment of Cell Viability and Proliferation in Hydrogel
2.9.1. Live Cell Imaging in 3D Scaffold
2.9.2. Alamar Blue Assay
2.10. Gene Expression Analysis
2.11. Statistical Analysis
3. Results
3.1. Decellularization of Human Umbilical Cord Tissue
DNA Quantification and Gel Electrophoresis
3.2. Biochemical Composition of Native and DUC Tissue
3.2.1. DAPI Staining
3.2.2. Hematoxylin and Eosin Staining
3.3. ECM Structure and Composition of Decellularized Tissue
3.3.1. sGAG Quantification
3.3.2. Glycosaminoglycans (GAGs) Content of Extracellular Matrix
3.3.3. Masson’s Trichrome Staining for Collagen
3.4. Hydrogel Structural and Mechanical Properties
3.4.1. Biochemical Analysis of Hydrogel by FTIR
3.4.2. Scanning Electron Microscopy
3.4.3. Swelling Behavior
3.5. UC-MSCs Encapsulated in 3D Hydrogel
F-Actin Staining
3.6. Proliferation and Viability in 3D Culture MSCs
3.6.1. Calcein AM Staining
3.6.2. Alamar Blue Assay
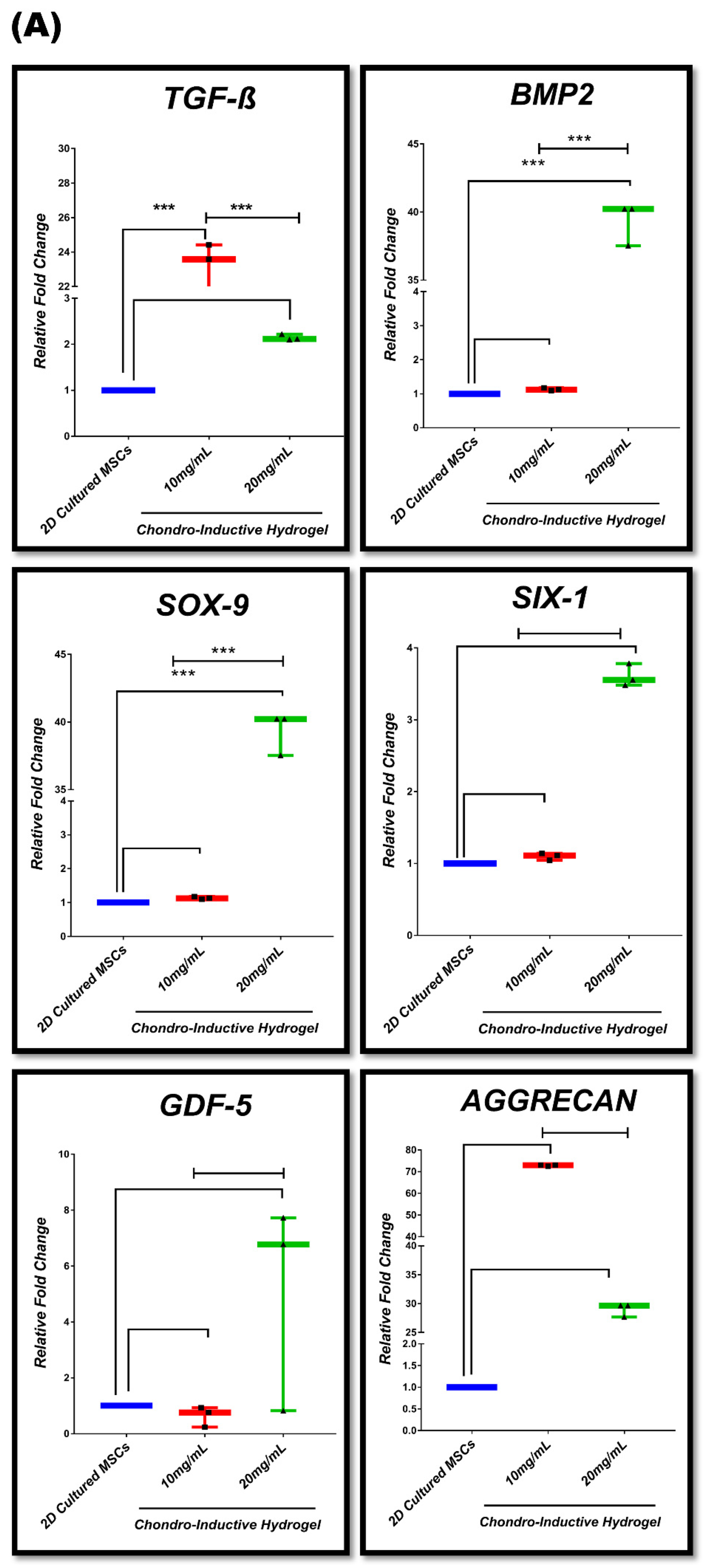
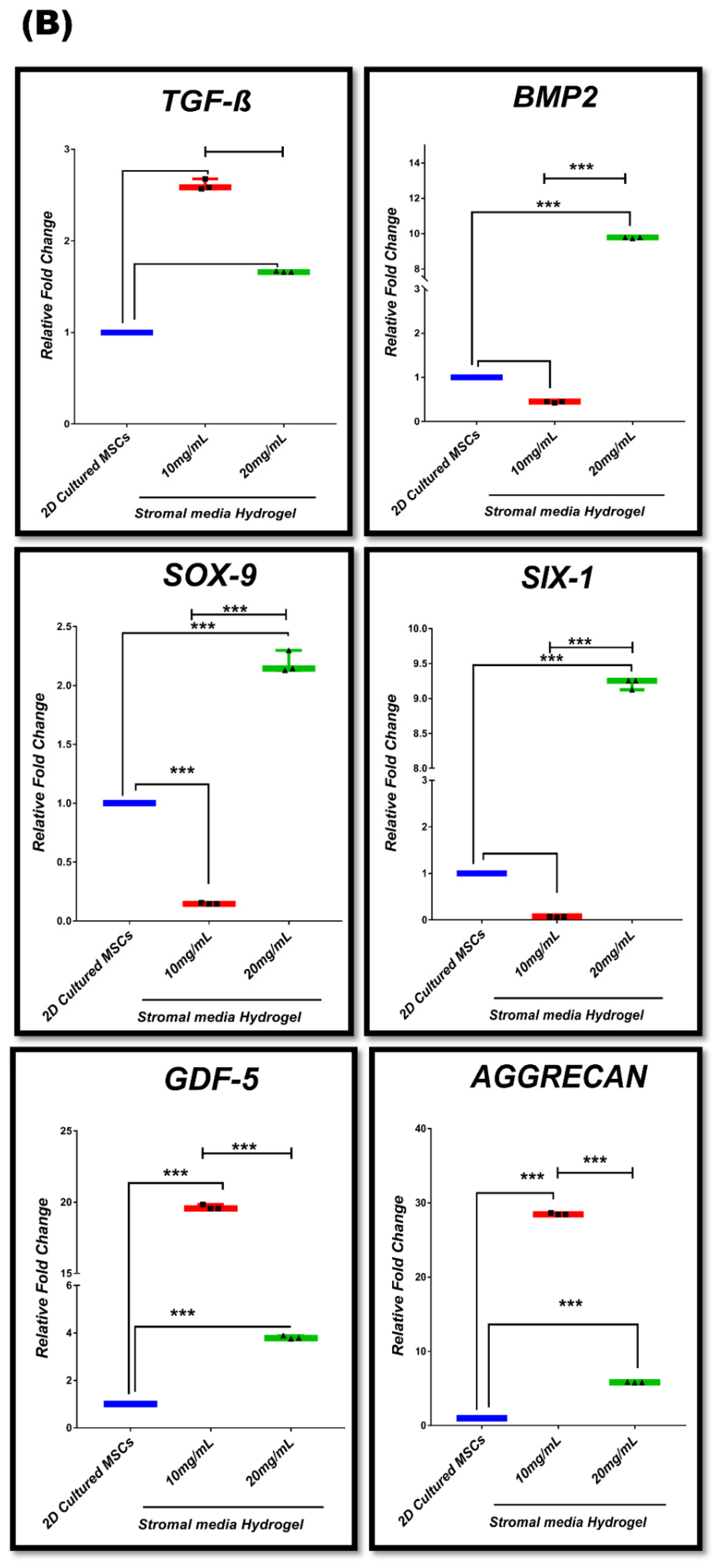
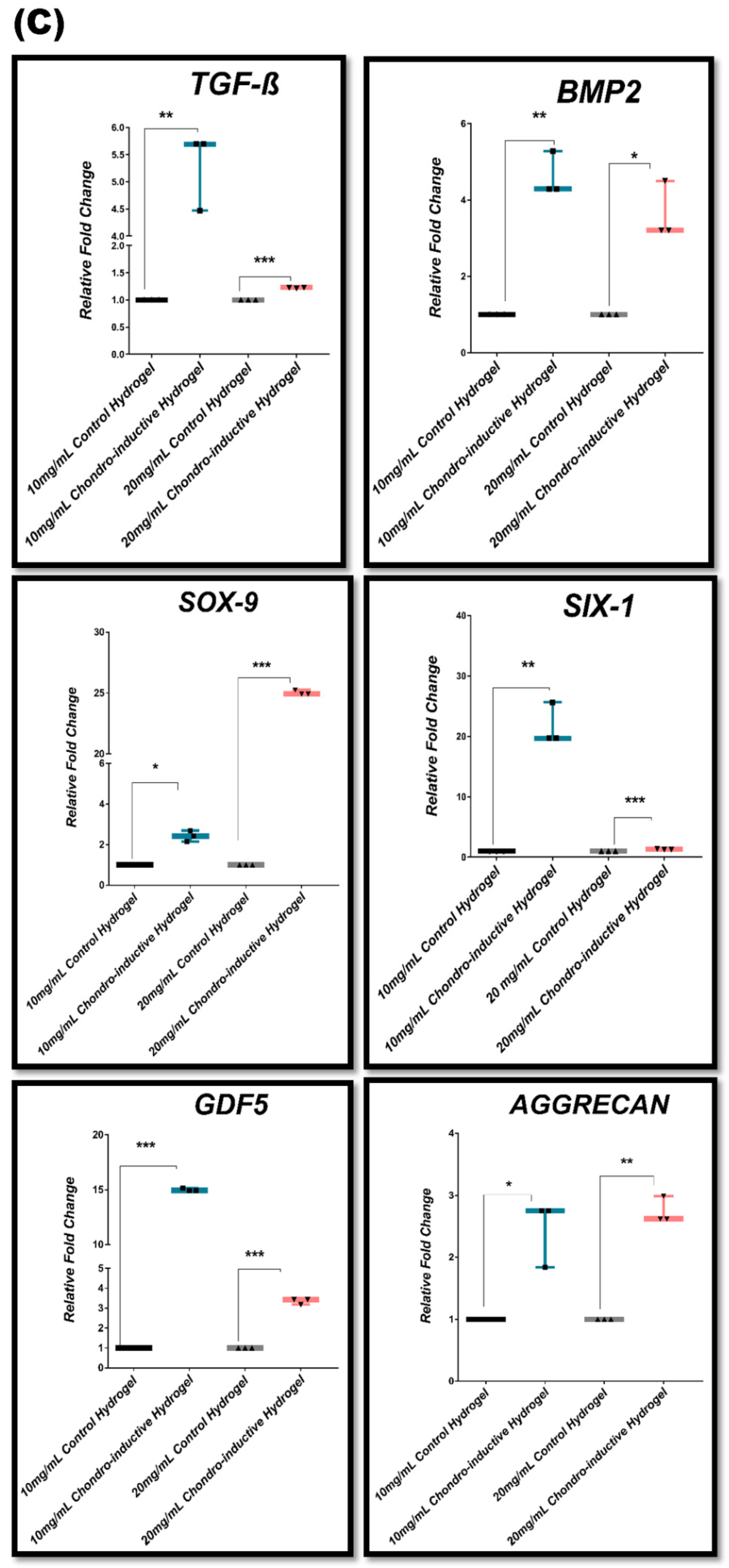
3.7. In Vitro Chondrogenic Gene Expression Dynamic
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Basiri, A.; Farokhi, M.; Azami, M.; Ebrahimi-Barough, S.; Mohamadnia, A.; Rashtbar, M.; Hasanzadeh, E.; Mahmoodi, N.; Eslaminejad, M.B.; Ai, J. A silk fibroin/decellularized extract of Wharton’s jelly hydrogel intended for cartilage tissue engineering. Prog. Biomater. 2019, 8, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.-X.; He, Y.; Bi, L.; Qu, Z.-H.; Zou, J.-W.; Pan, Z.; Fan, J.-J.; Chen, L.; Dong, X.; Liu, X.-N.; et al. Enhancing the bioactivity of Poly(lactic-co-glycolic acid) scaffold with a nano-hydroxyapatite coating for the treatment of segmental bone defect in a rabbit model. Int. J. Nanomed. 2013, 8, 1855–1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blahnová, V.; Vocetková, K.; Hlinkova, J.; Divin, R.; Amler, E.; Filová, E. PCL Scaffold for Osteochondral Defect Treatment. Key Eng. Mater. 2020, 834, 141–147. [Google Scholar] [CrossRef]
- Gomoll, A.H.; Madry, H.; Knutsen, G.; Van Dijk, N.; Seil, R.; Brittberg, M.; Kon, E. The subchondral bone in articular cartilage repair: Current problems in the surgical management. Knee Surg. Sports Traumatol. Arthrosc. 2010, 18, 434–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J.P.; Bhuiyan, D.B.; Ogle, B.M. Solid organ fabrication: Comparison of decellularization to 3D bioprinting. Biomater. Res. 2016, 20, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [Green Version]
- Gentile, P.; Nandagiri, V.K.; Daly, J.; Chiono, V.; Mattu, C.; Tonda-Turo, C.; Ciardelli, G.; Ramtoola, Z. Localised controlled release of simvastatin from porous chitosan–gelatin scaffolds engrafted with simvastatin loaded PLGA-microparticles for bone tissue engineering application. Mater. Sci. Eng. C 2016, 59, 249–257. [Google Scholar] [CrossRef]
- Gentili, C.; Cancedda, R. Cartilage and Bone Extracellular Matrix. Curr. Pharm. Des. 2009, 15, 1334–1348. [Google Scholar] [CrossRef]
- Pati, F.; Song, T.H.; Rijal, G.; Jiang, J.; Kim, S.-W.; Cho, D.-W. Ornamenting 3D printed scaffolds with cell-laid extracellular matrix for bone tissue regeneration. Biomaterials 2015, 37, 230–241. [Google Scholar] [CrossRef]
- Alford, A.I.; Kozloff, K.M.; Hankenson, K.D. Extracellular matrix networks in bone remodeling. Int. J. Biochem. Cell Biol. 2015, 65, 20–31. [Google Scholar] [CrossRef]
- Macías, I.; Alcorta-Sevillano, N.; Rodríguez, C.I.; Infante, A. Osteoporosis and the Potential of Cell-Based Therapeutic Strategies. Int. J. Mol. Sci. 2020, 21, 1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegenscheid, K.; Puls, R.; Rosenberg, C. Imaging strategies for knee injuries. Radiologe 2012, 52, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.X. Biomimetic materials for tissue engineering. Adv. Drug Deliv. Rev. 2008, 60, 184–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natu, M.V.; Sardinha, J.P.; Correia, I.J.; Gil, M.H. Controlled release gelatin hydrogels and lyophilisates with potential application as ocular inserts. Biomed. Mater. 2007, 2, 241. [Google Scholar] [CrossRef]
- Rana, D.; Zreiqat, H.; Benkirane-Jessel, N.; Ramakrishna, S. Development of decellularized scaffolds for stem cell-driven tissue engineering. J. Tissue Eng. Regen. Med. 2017, 11, 942–965. [Google Scholar] [CrossRef]
- Thakkar, S.; Fernandes, H.; Moroni, L. Decellularized extracellular matrix scaffolds for cartilage regeneration. Methods Mol. Biol. 2015, 1340, 133–151. [Google Scholar]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef]
- Hutter, H.; Vogel, B.E.; Plenefisch, J.D.; Norris, C.R.; Proenca, R.B.; Spieth, J.; Guo, C.; Mastwal, S.; Zhu, X.; Scheel, J.; et al. Conservation and Novelty in the Evolution of Cell Adhesion and Extracellular Matrix Genes. Science 2000, 287, 989–994. [Google Scholar] [CrossRef]
- Leor, J.; Amsalem, Y.; Cohen, S. Cells, scaffolds, and molecules for myocardial tissue engineering. Pharmacol. Ther. 2005, 105, 151–163. [Google Scholar] [CrossRef]
- Sabetkish, S.; Kajbafzadeh, A.M.; Sabetkish, N.; Khorramirouz, R.; Akbarzadeh, A.; Seyedian, S.L.; Pasalar, P.; Orangian, S.; Beigi, R.S.H.; Aryan, Z.; et al. Whole-organ tissue engineering: Decellularization and recellularization of three-dimensional matrix liver scaffolds. J. Biomed. Mater. Res.—Part A 2015, 103, 1498–1508. [Google Scholar] [CrossRef]
- Hoshiba, T.; Lu, H.; Kawazoe, N.; Chen, G. Decellularized matrices for tissue engineering. Expert Opin. Biol. Ther. 2010, 10, 1717–1728. [Google Scholar] [CrossRef] [PubMed]
- DeQuach, J.A.; Mezzano, V.; Miglani, A.; Lange, S.; Keller, G.; Sheikh, F.; Christman, K.L. Simple and High Yielding Method for Preparing Tissue Specific Extracellular Matrix Coatings for Cell Culture. PLoS ONE 2010, 5, e13039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, M.; Payne, T.R.; Oshima, H.; Momoi, N.; Tobita, K.; Huard, J. Differential efficacy of gels derived from small intestinal submucosa as an injectable biomaterial for myocardial infarct repair. Biomaterials 2010, 31, 7678–7683. [Google Scholar] [CrossRef] [PubMed]
- Tukmachev, D.; Forostyak, S.; Koci, Z.; Zaviskova, K.; Vackova, I.; Vyborny, K.; Sandvig, I.; Sandvig, A.; Medberry, C.J.; Badylak, S.; et al. Injectable Extracellular Matrix Hydrogels as Scaffolds for Spinal Cord Injury Repair. Tissue Eng. Part A 2016, 22, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, A.; Oh, S.J. Age related changes of the extracellular matrix and stem cell maintenance. Prev. Med. 2012, 54, S50–S56. [Google Scholar] [CrossRef]
- Hoshiba, T.; Chen, G.; Endo, C.; Maruyama, H.; Wakui, M.; Nemoto, E.; Kawazoe, N.; Tanaka, M. Decellularized Extracellular Matrix as an In Vitro Model to Study the Comprehensive Roles of the ECM in Stem Cell Differentiation. Stem Cells Int. 2015, 2016, 6397820. [Google Scholar] [CrossRef] [Green Version]
- Badylak, S.F. Decellularized allogeneic and xenogeneic tissue as a bioscaffold for regenerative medicine: Factors that influence the host response. Ann. Biomed. Eng. 2014, 42, 1517–1527. [Google Scholar] [CrossRef]
- Freytes, D.O.; Martin, J.; Velankar, S.S.; Lee, A.S.; Badylak, S.F. Preparation and rheological characterization of a gel form of the porcine urinary bladder matrix. Biomaterials 2008, 29, 1630–1637. [Google Scholar] [CrossRef]
- Kočí, Z.; Výborný, K.; Dubišová, J.; Lee, A.S.; Badylak, S.F. Extracellular matrix hydrogel derived from human umbilical cord as a scaffold for neural tissue repair and its comparison with extracellular matrix from porcine tissues. Tissue Eng—Part C Methods 2017, 23, 333–345. [Google Scholar] [CrossRef]
- Safari, F.; Fani, N.; Eglin, D.; Alini, M.; Stoddart, M.J.; Eslaminejad, M.B. Human umbilical cord-derived scaffolds for cartilage tissue engineering. J. Biomed. Mater. Res. Part A 2019, 107, 1793–1802. [Google Scholar] [CrossRef]
- Beiki, B.; Zeynali, B.; Seyedjafari, E. Fabrication of a three dimensional spongy scaffold using human Wharton’s jelly derived extra cellular matrix for wound healing. Mater. Sci. Eng. C 2017, 78, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Ekram, S.; Salim, A.; Chaudhry, G.R.; Khan, I. Transcription regulators differentiate mesenchymal stem cells into chondroprogenitors, and their in vivo implantation regenerated the intervertebral disc degeneration. World J. Stem. Cells 2022, 14, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Ekram, S.; Khalid, S.; Bashir, I.; Salim, A.; Khan, I. Human umbilical cord-derived mesenchymal stem cells and their chondroprogenitor derivatives reduced pain and inflammation signaling and promote regeneration in a rat intervertebral disc degeneration model. Mol. Cell. Biochem. 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, O.A.; O’Donnell, B.; Poche, J.N.; Iftikhar, R.; Wise, R.M.; Motherwell, J.M.; Campbell, B.; Savkovic, S.D.; Bunnell, B.A.; Hayes, D.; et al. Human Adipose-Derived Hydrogel Characterization Based on In Vitro ASC Biocompatibility and Differentiation. Stem Cells Int. 2019, 2019, 9276398. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.R.; Ahmad, W.; Naeem, N.; Salim, A.; Khan, I. Small molecule 2′-deoxycytidine differentiates human umbilical cord-derived MSCs into cardiac progenitors in vitro and their in vivo xeno-transplantation improves cardiac function. Mol. Cell. Biochem. 2020, 470, 99–113. [Google Scholar] [CrossRef]
- Jacob, G.; Shimomura, K.; Nakamura, N. Osteochondral Injury, Management and Tissue Engineering Approaches. Front. Cell Dev. Biol. 2020, 8, 1118. [Google Scholar] [CrossRef]
- Sorrento, D.L.; Mlodzienski, A. Incidence of lateral talar dome lesions in SER, I.V. ankle fractures. J. Foot Ankle Surg. 2000, 39, 354–358. [Google Scholar] [CrossRef]
- Kumaresan, S.; Chokalingam, K.; Sridhar, K.N.; Veerichetty, V. Development of Human Umbilical cord based scaffold for tissue engineering application. AIP Conf. Proc. 2020, 2270, 020005. [Google Scholar]
- Solorio, L.D.; Phillips, L.M.; Mcmillan, A.; Cheng, C.W.; Dang, P.N.; Samorezov, J.E.; Yu, X.H.; Murphy, W.L.; Alsberg, E. Spatially Organized Differentiation of Mesenchymal Stem Cells within Biphasic Microparticle-Incorporated High Cell Density Osteochondral Tissues. Adv. Health Mater. 2015, 4, 2306–2313. [Google Scholar] [CrossRef] [Green Version]
- Crapo, P.M.; Gilbert, T.W.; Badylak, S.F. An overview of tissue and whole organ decellularization processes. Biomaterials 2011, 32, 3233–3243. [Google Scholar] [CrossRef] [Green Version]
- Badylak, S.F.; Gilbert, T.W. Immune response to biologic scaffold materials. Semin. Immunol. 2008, 20, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forraz, N.; Mcguckin, C.P. The umbilical cord: A rich and ethical stem cell source to advance regenerative medicine. Cell Prolif. 2011, 44, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.; Guo, W.; Chen, M.; Hao, C.; Gao, S.; Huang, J.; Yuan, Z.; Zhang, Y.; Wang, M.; Li, P.; et al. Fabrication and In Vitro Study of Tissue-Engineered Cartilage Scaffold Derived from Wharton’s Jelly Extracellular Matrix. BioMed Res. Int. 2017, 2017, 5839071. [Google Scholar] [CrossRef] [PubMed]
- Jadalannagari, S.; Converse, G.; McFall, C.; Buse, E.; Filla, M.; Villar, M.T.; Artigues, A.; Mellot, A.J.; Wang, J.; Detamore, M.S.; et al. Decellularized Wharton’s Jelly from human umbilical cord as a novel 3D scaffolding material for tissue engineering applications. PLoS ONE 2017, 12, e0172098. [Google Scholar] [CrossRef]
- Gilpin, A.; Yang, Y. Decellularization Strategies for Regenerative Medicine: From Processing Techniques to Applications. Biomed Res. Int. BioMed Res. Int. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.N.; Badylak, S.F. Expanded applications, shifting paradigms and an improved understanding of host-biomaterial interactions. Acta Biomater. 2013, 9, 4948–4955. [Google Scholar] [CrossRef]
- Seif-Naraghi, S.B.; Salvatore, M.A.; Schup-Magoffin, P.J.; Hu, D.P.; Christman, K.L. Design and Characterization of an Injectable Pericardial Matrix Gel: A Potentially Autologous Scaffold for Cardiac Tissue Engineering. Tissue Eng. Part A 2010, 16, 2017–2027. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.D.; Lin, S.Y.; Christman, K.L. Tailoring material properties of a nanofibrous extracellular matrix derived hydrogel. Nanotechnology 2011, 22, 494015. [Google Scholar] [CrossRef] [Green Version]
- DeQuach, J.A.; Yuan, S.H.; Goldstein, L.S.; Christman, K.L. Decellularized Porcine Brain Matrix for Cell Culture and Tissue Engineering Scaffolds. Tissue Eng. Part A 2011, 17, 2583–2592. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Ji, T.; Wang, X.; Fu, W.; Ye, L.-C.; Zhang, H.; Li, F. Engineering cartilage tissue based on cartilage-derived extracellular matrix cECM/PCL hybrid nanofibrous scaffold. Mater. Des. 2020, 193, 108773. [Google Scholar] [CrossRef]
- Kliemt, S.; Lange, C.; Otto, W.; Hintze, V.; Möller, S.; Von Bergen, M.; Hempel, U.; Kalkhof, S. Sulfated Hyaluronan Containing Collagen Matrices Enhance Cell-Matrix-Interaction, Endocytosis, and Osteogenic Differentiation of Human Mesenchymal Stromal Cells. J. Proteome Res. 2012, 12, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Van Susante, J.L.C.; Pieper, J.; Buma, P.; Kuppevelt, T.-H.-V.; Beuningen, H.-V.; Kraan, P.-M.-V.-D.; Veerkamp, J.-H.; Berg, W.-B.-V.-D.; Veth, R.-R.-H. Linkage of chondroitin-sulfate to type, I. collagen scaffolds stimulates the bioactivity of seeded chondrocytes in vitro. Biomaterials 2001, 22, 2359–2369. [Google Scholar] [CrossRef]
- Smith, L.R.; Cho, S.; Discher, D.E. Stem cell differentiation is regulated by extracellular matrix mechanics. Physiology 2018, 33, 16–25. [Google Scholar] [CrossRef]
- Haugh, M.G.; Vaughan, T.J.; Madl, C.M.; Raftery, R.; McNamara, L.M.; O’Brien, F.J.; Heilshorn, S.C. Investigating the interplay between substrate stiffness and ligand chemistry in directing mesenchymal stem cell differentiation within 3D macro-porous substrates. Biomaterials 2018, 171, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhang, D.; Zhou, C.; Yuan, Q.; Ye, L.; Zhou, X.-D. Substrate elasticity regulates adipose-derived stromal cell differentiation towards osteogenesis and adipogenesis through β-catenin transduction. Acta Biomater. 2018, 79, 83–95. [Google Scholar] [CrossRef]
- Wen, J.H.; Vincent, L.G.; Fuhrmann, A.; Choi, Y.S.; Hribar, K.C.; Taylor-Weiner, H.; Chen, S.; Engler, A.J. Interplay of matrix stiffness and protein tethering in stem cell differentiation. Nat. Mater. 2014, 13, 979–987. [Google Scholar] [CrossRef] [Green Version]
- Khodadi, E.; Asnafi, A.A.; Shahrabi, S.; Shahjahani, M.; Saki, N. Bone marrow niche in immune thrombocytopenia: A focus on megakaryopoiesis. Ann. Hematol. 2016, 95, 1765–1776. [Google Scholar] [CrossRef]
- Sobolewski, K.; Małkowski, A.; Bańkowski, E.; Jaworski, S. Wharton’s jelly as a reservoir of peptide growth factors. Placenta 2005, 26, 747–752. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Nalinanon, S.; Benjakul, S.; Kishimura, H. Characteristics of Pepsin-Solubilised Collagen from the Skin of Splendid Squid (Loligo formosana). J. Chem. 2015, 2015, 482354. [Google Scholar] [CrossRef] [Green Version]
- Belbachir, K.; Noreen, R.; Gouspillou, G.; Petibois, C. Collagen types analysis and differentiation by FTIR spectroscopy. Anal. Bioanal. Chem. 2009, 395, 829–837. [Google Scholar] [CrossRef]
- Yacob, N.; Hashim, K. Morphological effect on swelling behaviour of hydrogel. AIP Conf. Proc. 2014. [Google Scholar] [CrossRef] [Green Version]
- Mohiuddin, O.A.; Motherwell, J.M.; Rogers, E.; Bratton, M.R.; Zhang, Q.; Wang, G.; Bunnell, B.; Hayes, D.J.; Gimble, J.M. Characterization and Proteomic Analysis of Decellularized Adipose Tissue Hydrogels Derived from Lean and Overweight/Obese Human Donors. Adv. Biosyst. 2020, 4. [Google Scholar] [CrossRef] [PubMed]
- Alam, K.; Iqbal, M.; Hasan, A.; Al-Maskari, N. Rheological Characterization of Biological Hydrogels in Aqueous State. J. Appl. Biotechnol. Rep. 2020, 7, 171–175. [Google Scholar] [CrossRef]
- Huang, J.; Huang, Z.; Liang, Y.; Yuan, W.; Bian, L.; Duan, L.; Rong, Z.; Xiong, J.; Wang, D.; Xia, J. 3D printed gelatin/hydroxyapatite scaffolds for stem cell chondrogenic differentiation and articular cartilage repair. Biomater. Sci. 2021, 9, 2620–2630. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.; Liao, Q.; Gee, C.W. Surgical Management of Osteochondral Defects of the Knee: An Educational Review. Curr. Rev. Musculoskelet. Med. 2021, 14, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Chaboissier, M.-C.; Martin, J.F.; Schedl, A.; de Crombrugghe, B. The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takigawa, Y.; Hata, K.; Muramatsu, S.; Amano, K.; Ono, K.; Wakabayashi, M.; Matsuda, A.; Takada, K.; Nishimura, R.; Yoneda, T. The transcription factor Znf219 regulates chondrocyte differentiation by assembling a transcription factory with Sox9. J. Cell Sci. 2010, 123, 3780–3788. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, H.; Yokoyama, S.; Asahara, H. Growth and differentiation of the developing limb bud from the perspective of chondrogenesis. Dev. Growth Differ. 2007, 49, 449–454. [Google Scholar] [CrossRef]
- Djouad, F.; Delorme, B.; Maurice, M.; Bony, C.; Apparailly, F.; Louis-Plence, P.; Canovas, F.; Charbord, P.; Noël, D.; Jorgensen, C. Microenvironmental changes during differentiation of mesenchymal stem cells towards chondrocytes. Arthritis Res. Ther. 2007, 9, R33. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, Y.; Tuan, R.S.; Shum, L. Distinct functions of BMP4 and GDF5 in the regulation of chondrogenesis. J. Cell. Biochem. 2004, 91, 1204–1217. [Google Scholar] [CrossRef]
- Goldring, M.B.; Tsuchimochi, K.; Ijiri, K. The control of chondrogenesis. J. Cell. Biochem. 2005, 97, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Furumatsu, T.; Tsuda, M.; Taniguchi, N.; Tajima, Y.; Asahara, H. Smad3 Induces Chondrogenesis through the Activation of SOX9 via CREB-binding Protein/p300 Recruitment. J. Biol. Chem. 2005, 280, 8343–8350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, B.; Steinberg, T.; Giselbrecht, S.; Gottwald, E.; Tomakidi, P.; Bächle-Haas, M.; Kohal, R.-J. Promotion of osteoblast differentiation in 3D biomaterial micro-chip arrays comprising fibronectin-coated poly(methyl methacrylate) polycarbonate. Biomaterials 2011, 32, 8947–8956. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.S.; Rasmussen, P.L.; Larsen, K.H.; Schrøder, H.D.; Kassem, M. Parameters in Three-Dimensional Osteospheroids of Telomerized Human Mesenchymal (Stromal) Stem Cells Grown on Osteoconductive Scaffolds That Predict In Vivo Bone-Forming Potential. Tissue Eng. Part A 2010, 16, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hantash, B.M. TGF-β1 Regulates Differentiation of Bone Marrow Mesenchymal Stem Cells. Vitam. Horm. 2011, 87, 127–141. [Google Scholar] [PubMed]
- Schagemann, J.C.; Paul, S.; Casper, M.E.; Rohwedel, J.; Kramer, J.; Kaps, C.; Mittelstaedt, H.; Fehr, M.; Reinholz, G.G. Chondrogenic differentiation of bone marrow-derived mesenchymal stromal cells via biomimetic and bioactive poly-ε-caprolactone scaffolds. J. Biomed. Mater. Res. Part A 2012, 101A, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- De Mara, C.S.; Duarte, A.S.S.; Sartori, A.; Luzo, A.-C.; Sara, S.-T.-O.; Coimbra, I.-B. Regulation of chondrogenesis by transforming growth factor-β3 and insulin-like growth factor-1 from human mesenchymal umbilical cord blood cells. J. Rheumatol. 2010, 37, 1519–1526. [Google Scholar] [CrossRef]
- Lee, P.T.; Li, W.J. Chondrogenesis of Embryonic Stem Cell-Derived Mesenchymal Stem Cells Induced by TGFβ1 and BMP7 Through Increased TGFβ Receptor Expression and Endogenous TGFβ1 Production. J. Cell. Biochem. 2016, 118, 172–181. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Primers | Primer Sequences (5′-3′) | Annealing Temperatures (°C) |
|---|---|---|
| GAPDH | 5′-CACCATGGGGAAGGTGAAGG -3′ 5′-AGCATCGCCCCACTTGATTT -3′ | 58 |
| TGF-β1 | 5′-CAAGGCACAGGGGACCAG -3′ 5′-CAGGTTCCTGGTGGGCAG -3′ | 58 |
| BMP2 | 5′-AGCTGGGCCGCAGGA -3′ 5′-TCGGCTGGCTGCCCT -3′ | 58 |
| SOX-9 | 5′-CGGGCAAGGCTGACCTG -3′ 5′-GGTGCTGCTGATGCCGT -3′ | 58 |
| SIX-1 | 5′-CTCCAGTCTGGTGGACTTGG-3′ 5′-AGCTTGAGATCGCTGTTGGT -3′ | 58 |
| GDF5 | 5′-CACATCCCAAGAGCCCCTTC -3′ 5′-GCCCAGGTGAGGAGAAATGG -3′ | 58 |
| ACCAN | 5′- AGTTCTGTGAATCTCACAATGCC-3′ 5′- CCAGAGGGACTGACATTTTCTTG-3′ | 58 |
| Group #01 | 2D-cultured MSCs in stromal media (control) vs. 10 mg/mL chondro-inductive hydrogel 20 mg/mL chondro-inductive hydrogel |
| Group #02 | 2D-cultured MSCs in stromal media (control) vs. 10 mg/mL stromal media hydrogel 20 mg/mL stromal media hydrogel |
| Group #03 | 10 mg/mL stromal media hydrogel (control) 20 mg/mL stromal media hydrogel vs. 10 mg/mL chondro-inductive hydrogel 20 mg/mL chondro-inductive hydrogel |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramzan, F.; Ekram, S.; Frazier, T.; Salim, A.; Mohiuddin, O.A.; Khan, I. Decellularized Human Umbilical Tissue-Derived Hydrogels Promote Proliferation and Chondrogenic Differentiation of Mesenchymal Stem Cells. Bioengineering 2022, 9, 239. https://doi.org/10.3390/bioengineering9060239
Ramzan F, Ekram S, Frazier T, Salim A, Mohiuddin OA, Khan I. Decellularized Human Umbilical Tissue-Derived Hydrogels Promote Proliferation and Chondrogenic Differentiation of Mesenchymal Stem Cells. Bioengineering. 2022; 9(6):239. https://doi.org/10.3390/bioengineering9060239
Chicago/Turabian StyleRamzan, Faiza, Sobia Ekram, Trivia Frazier, Asmat Salim, Omair Anwar Mohiuddin, and Irfan Khan. 2022. "Decellularized Human Umbilical Tissue-Derived Hydrogels Promote Proliferation and Chondrogenic Differentiation of Mesenchymal Stem Cells" Bioengineering 9, no. 6: 239. https://doi.org/10.3390/bioengineering9060239