
Genomic Insight into Shimazuella Soli Sp. Nov. Isolated from Soil and Its Putative Novel Class II Lasso Peptide

Abstract
:1. Introduction
2. Materials and Methods
2.1. The Media and Reagents
2.2. Bacteria and Culture Condition
2.3. Phylogenetic Analysis
2.4. Genome Sequencing and Genomic Analysis
2.5. Morphological, Phenotypic and Physiological Analysis
2.6. Chemotaxonomy
2.7. Molecular Modelling and Docking
3. Results
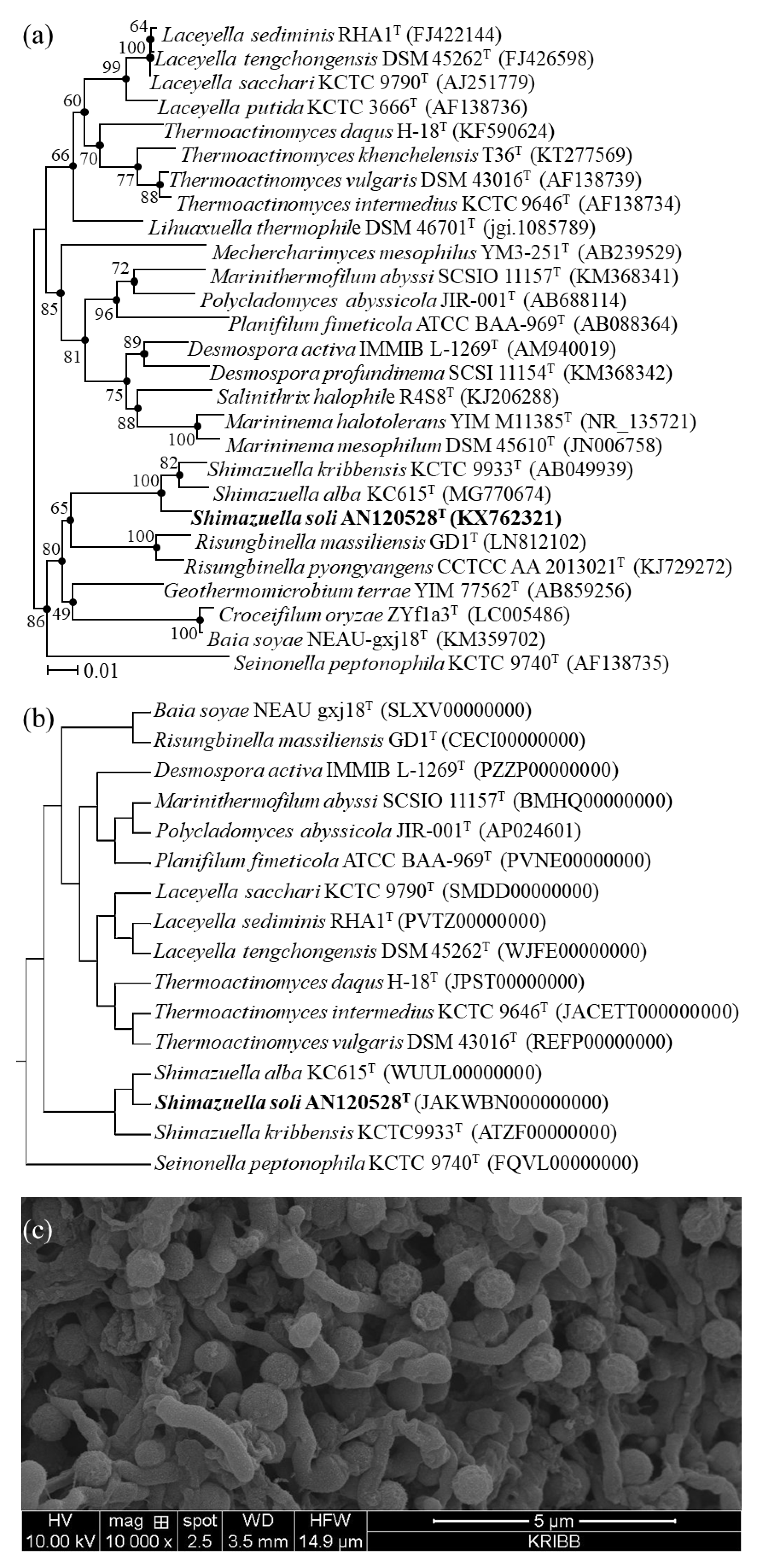
3.1. Phylogenetic Analysis
3.2. Morphological, Physiological and Biochemical Characteristics
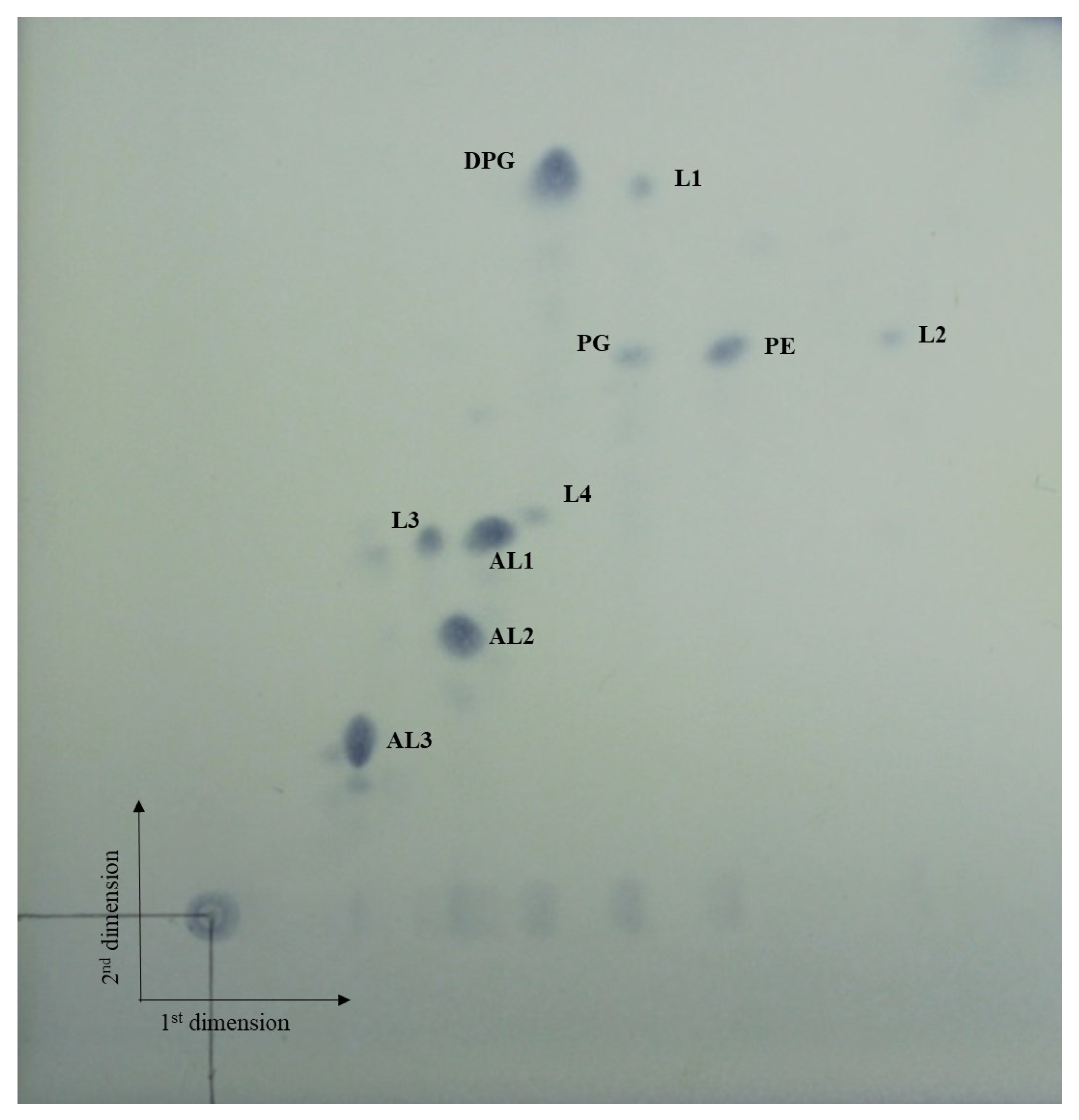
3.3. Chemotaxonomic Characteristics
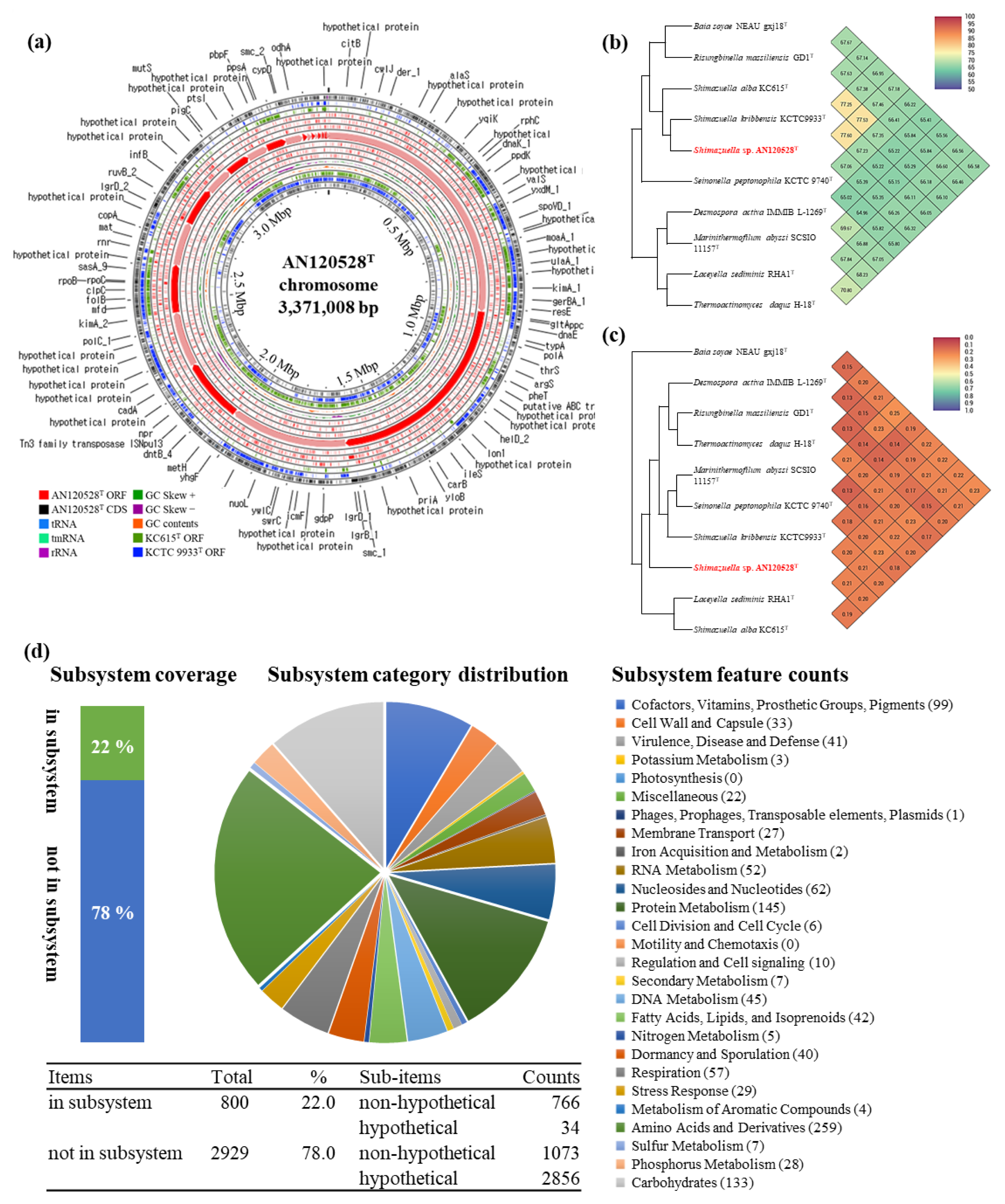
3.4. Genome Analysis
3.5. In Silico Analysis of the Novel Lasso Peptide Shimazuellin of AN120528T
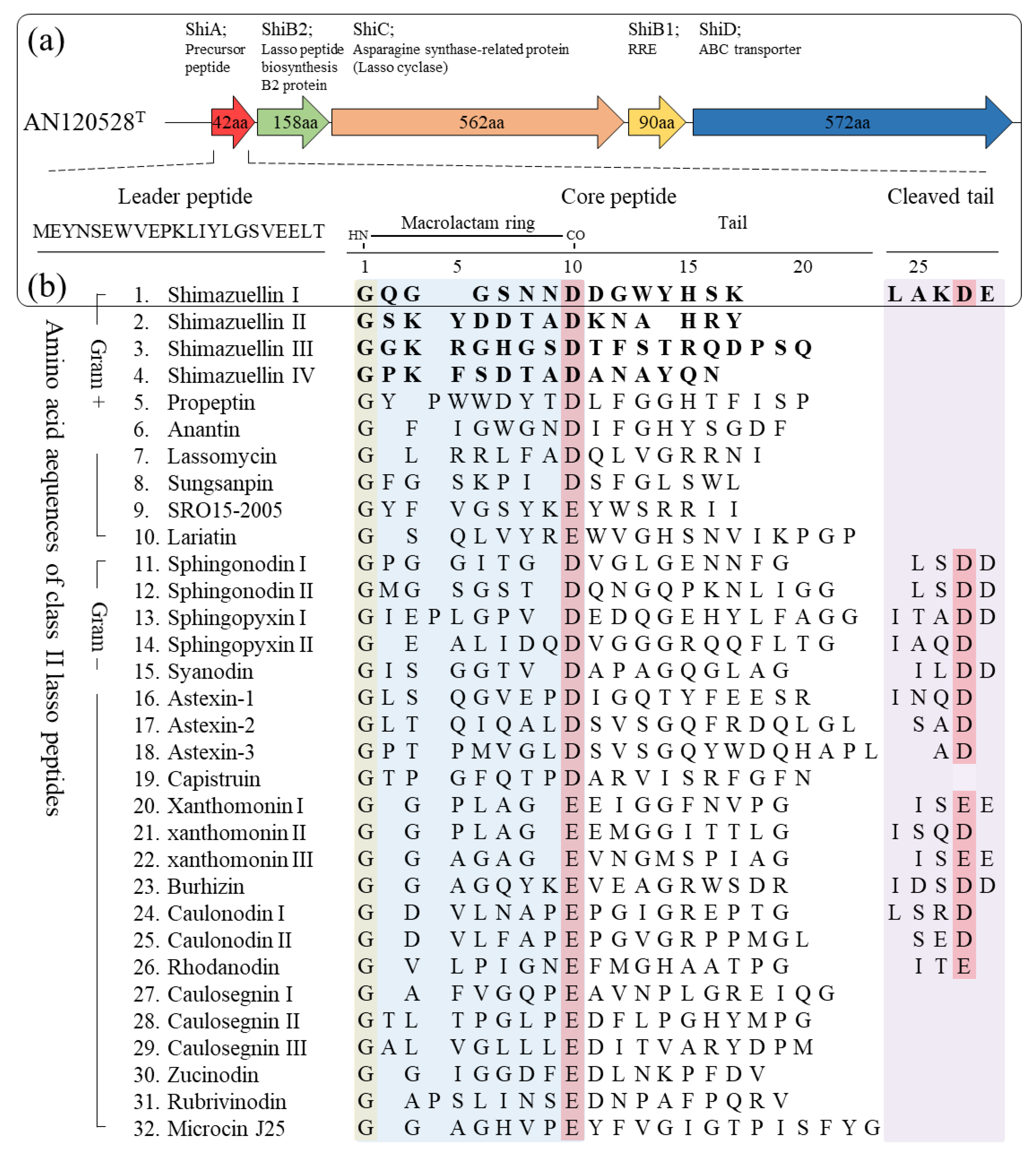
3.5.1. Genome Mining and Identification of Shimazuellin BGCs
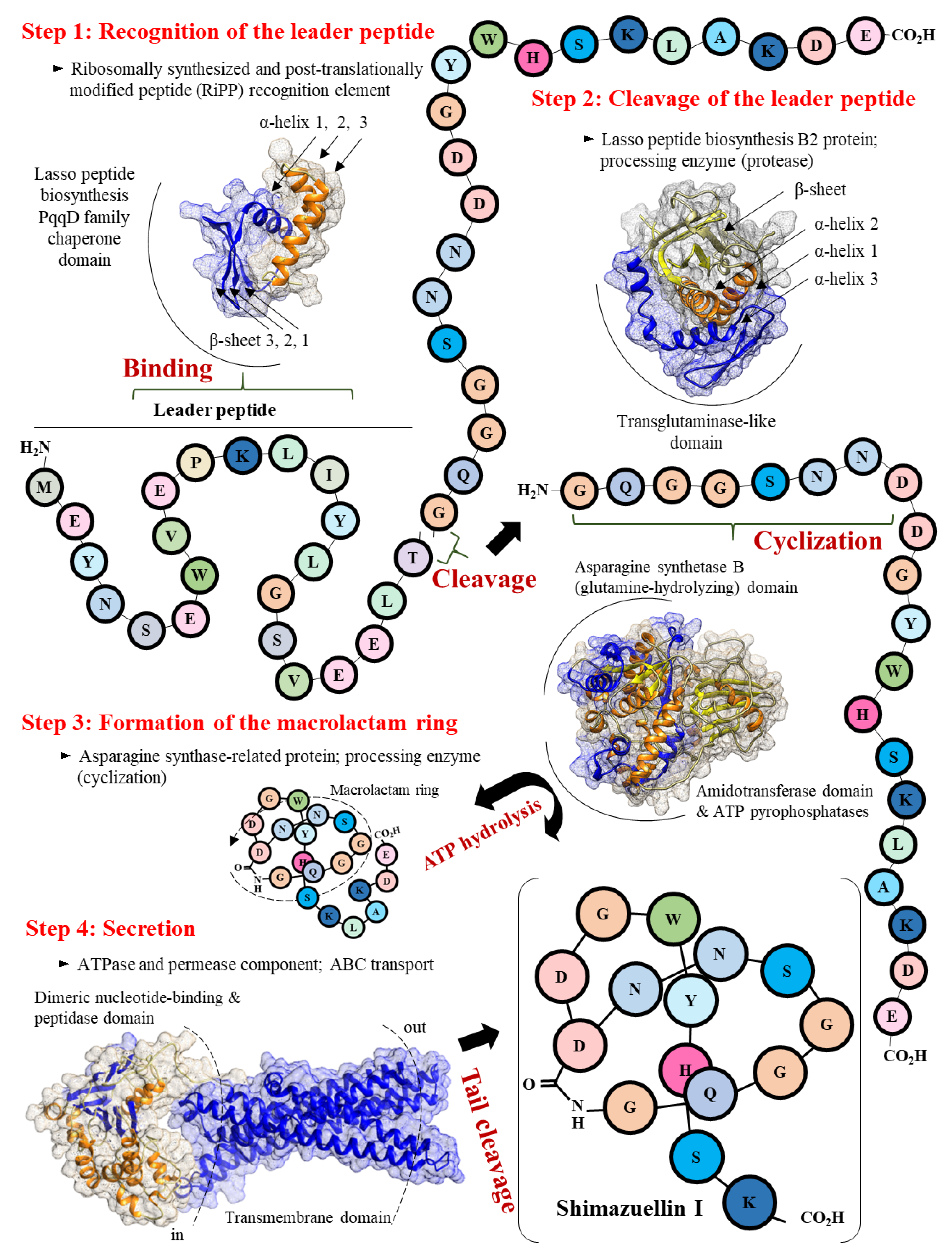
3.5.2. Scheme of the Putative Biosynthetic Mechanism of Shimazuellin in AN120528T
3.6. Description of Shimazuella Soli Sp. Nov.
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Park, D.-J.; Dastager, S.G.; Lee, J.-C.; Yeo, S.-H.; Yoon, J.-H.; Kim, C.-J. Shimazuella kribbensis gen. nov., sp. nov., a mesophilic representative of the family Thermoactinomycetaceae. Int. J. Syst. Evol. Microbiol. 2007, 57, 2660–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parte, A.C.; Sarda Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Saygin, H.; Ay, H.; Sahin, N. Shimazuella alba sp. nov. isolated from desert soil and emended description of the genus Shimazuella Park et al. 2007. Arch. Microbiol. 2020, 202, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Singh, A. Materials modulate immunity and gut microbiome. Nat. Mater. 2020, 19, 3–4. [Google Scholar] [CrossRef]
- Sumida, T.; Dubiley, S.; Wilcox, B.; Severinov, K.; Tagami, S. Structural basis of leader peptide recognition in lasso peptide biosynthesis pathway. ACS Chem. Biol. 2019, 14, 1619–1627. [Google Scholar] [CrossRef]
- Cheng, C.; Hua, Z.-C. Lasso Peptides: Heterologous production and potential medical application. Front. Bioeng. Biotechnol. 2020, 8, 571165. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Fitch, W.M. Toward defining the course of evolution: Minimum change for a specific tree topology. Syst. Biol. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.; Kim, Y.O.; Park, S.-C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef] [PubMed]
- Auch, A.F.; Von Jan, M.; Klenk, H.-P.; Göker, M. Digital DNA-DNA hybridization for microbial species delineation by means of genome-to-genome sequence comparison. Stand. Genom. Sci. 2010, 2, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Lee, J.M.; Jin, C.-Z.; Kang, M.-K.; Park, S.H.; Park, D.-J.; Kim, D.-G.; Kim, C.-J. Nocardioides humilatus sp. nov., isolated from farmland soil in the Republic of Korea. Int. J. Syst. Evol. Microbiol. 2022, 72, 004928. [Google Scholar] [CrossRef]
- Lee, J.M.; Jin, C.-Z.; Park, S.H.; Kang, M.-K.; Park, D.-J.; Kim, C.-J. Nocardioides antri sp. nov., Isolated from Soil in a Rock Cave. Curr. Microbiol. 2021, 78, 2130–2135. [Google Scholar] [CrossRef]
- Skerman, V.B.D. A Guide to the Identification of the Genera of Bacteria. J. Med. Educ. 1960, 35, 92. [Google Scholar]
- Jin, C.-Z.; Jin, L.; Liu, M.-J.; Lee, J.-M.; Park, D.-J.; Kim, C.-J. Solihabitans fulvus gen. nov., sp. nov., a member of the family Pseudonocardiaceae isolated from soil. Int. J. Syst. Evol. Microbiol. 2022, 72, 005110. [Google Scholar] [CrossRef]
- Jin, C.-Z.; Jin, L.; Kang, D.H.; Liu, M.-J.; Lee, J.M.; Park, D.-J.; Kim, C.-J. Description of desferrioxamine-producing bacterium Chitinophaga agrisoli sp. nov., isolated from soil. Antonie van Leeuwenhoek 2021, 114, 741–750. [Google Scholar] [CrossRef]
- Gonzalez, C.; Gutierrez, C.; Ramirez, C. Halobacterium vallismortis sp. nov. An amylolytic and carbohydrate-metabolizing, extremely halophilic bacterium. Can. J. Microbiol. 1978, 24, 710–715. [Google Scholar] [CrossRef]
- Kämpfer, P.; Kroppenstedt, R.M. Numerical analysis of fatty acid patterns of coryneform bacteria and related taxa. Can. J. Microbiol. 1996, 42, 989–1005. [Google Scholar] [CrossRef]
- Tamaoka, J.; Katayama-Fujimura, Y.; Kuraishi, H. Analysis of bacterial menaquinone mixtures by high performance liquid chromatography. J. Appl. Bacteriol. 1983, 54, 31–36. [Google Scholar] [CrossRef]
- Staneck, J.L.; Roberts, G.D. Simplified approach to identification of aerobic actinomycetes by thin-layer chromatography. Appl. Environ. Microbiol. 1974, 28, 226–231. [Google Scholar] [CrossRef]
- Komagata, K.; Suzuki, K.-I. 4 Lipid and cell-wall analysis in bacterial systematics. In Current Methods for Classification and Identification of Microorganisms; Colwell, R.R., Grigorova, R., Eds.; Methods in Microbiology; Academic Press: Cambridge, MA, USA, 1988; Volume 19, pp. 161–207. [Google Scholar]
- Lee, J.M.; Moon, S.Y.; Kim, Y.-R.; Kim, K.W.; Lee, B.-J.; Kong, I.-S. Improvement of thermostability and halostability of β-1,3-1,4-glucanase by substituting hydrophobic residue for Lys 48. Int. J. Biol. Macromol. 2017, 94, 594–602. [Google Scholar] [CrossRef]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef] [Green Version]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Res. 2011, 39 (Suppl. 2), W270–W277. [Google Scholar] [CrossRef] [Green Version]
- Tina, K.G.; Bhadra, R.; Srinivasan, N. PIC: Protein interactions calculator. Nucleic Acids Res. 2007, 35 (Suppl. 2), W473–W476. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, T.; Ri, S.; Jiang, F.; Chang, X.; Liu, Y.; Ren, L.; Qiu, X.; Deng, S.; Fang, C.; et al. Risungbinella pyongyangensis gen. nov., sp. nov., a mesophilic member of the family Thermoactinomycetaceae isolated from an agricultural soil sample. Int. J. Syst. Evol. Microbiol. 2015, 65, 2726–2733. [Google Scholar] [CrossRef] [PubMed]
- Burkhart, B.J.; Hudson, G.A.; Dunbar, K.L.; Mitchell, D.A. A prevalent peptide-binding domain guides ribosomal natural product biosynthesis. Nat. Chem. Biol. 2015, 11, 564–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksimov, M.O.; Link, A.J. Discovery and characterization of an isopeptidase that linearizes lasso peptides. J. Am. Chem. Soc. 2013, 135, 12038–12047. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Fage, C.D.; Hegemann, J.D.; Mielcarek, A.; Yan, D.; Linne, U.; Marahiel, M.A. The B1 Protein Guides the Biosynthesis of a Lasso Peptide. Sci. Rep. 2016, 6, 35604. [Google Scholar] [CrossRef] [Green Version]
- Cortés-Albayay, C.; Jarmusch, S.A.; Trusch, F.; Ebel, R.; Andrews, B.A.; Jaspars, M.; Asenjo, J.A. Downsizing class II lasso peptides: Genome mining-guided isolation of huascopeptin containing the first Gly1-Asp7 macrocycle. J. Org. Chem. 2020, 85, 1661–1667. [Google Scholar] [CrossRef]
- Oman, T.J.; Van Der Donk, W.A. Follow the leader: The use of leader peptides to guide natural product biosynthesis. Nat. Chem. Biol. 2010, 6, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Zirah, S.; Afonso, C.; Linne, U.; Knappe, T.A.; Marahiel, M.A.; Rebuffat, S.; Tabet, J.-C. Topoisomer differentiation of molecular knots by FTICR MS: Lessons from class II lasso peptides. J. Am. Soc. Mass Spectrom. 2011, 22, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, F.; Sementa, D.; Jain, A.; Kumar, M.; Tayarani-Najjaran, M.; Kroiss, D.; Ulijn, R.V. Peptide-based supramolecular systems chemistry. Chem. Rev. 2021, 121, 13869–13914. [Google Scholar] [CrossRef]
- Wang, L.; Wang, N.; Zhang, W.; Cheng, X.; Yan, Z.; Shao, G.; Wang, X.; Wang, R.; Fu, C. Therapeutic peptides: Current applications and future directions. Signal Transduct. Target. Ther. 2022, 7, 48. [Google Scholar] [CrossRef]
- Boolchandani, M.; D’Souza, A.W.; Dantas, G. Sequencing-based methods and resources to study antimicrobial resistance. Nat. Rev. Genet. 2019, 20, 356–370. [Google Scholar] [CrossRef]
- Sakai, K.; Passioura, T.; Sato, H.; Ito, K.; Furuhashi, H.; Umitsu, M.; Takagi, J.; Kato, Y.; Mukai, H.; Warashina, S.; et al. Macrocyclic peptide-based inhibition and imaging of hepatocyte growth factor. Nat. Chem. Biol. 2019, 15, 598–606. [Google Scholar] [CrossRef]
- Vinogradov, A.A.; Yin, Y.; Suga, H. Macrocyclic peptides as drug candidates: Recent progress and remaining challenges. J. Am. Chem. Soc. 2019, 141, 4167–4181. [Google Scholar] [CrossRef]
- Hegemann, J.D. Factors governing the thermal stability of lasso peptides. Chembiochem 2019, 21, 7–18. [Google Scholar] [CrossRef]
- Knappe, T.A.; Manzenrieder, F.; Mas-Moruno, C.; Linne, U.; Sasse, F.; Kessler, H.; Xie, X.; Marahiel, M.A. Introducing lasso peptides as molecular scaffolds for drug design: Engineering of an integrin antagonist. Angew. Chem. Int. Ed. 2011, 50, 8714–8717. [Google Scholar] [CrossRef] [Green Version]
- Mihara, E.; Watanabe, S.; Bashiruddin, N.K.; Nakamura, N.; Matoba, K.; Sano, Y.; Maini, R.; Yin, Y.; Sakai, K.; Arimori, T.; et al. Lasso-grafting of macrocyclic peptide pharmacophores yields multi-functional proteins. Nat. Commun. 2021, 12, 1543. [Google Scholar] [CrossRef]
- Rodríguez, V. Insights into post-translational modification enzymes from RiPPs: A toolkit for applications in peptide synthesis. Biotechnol. Adv. 2022, 56, 107908. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | AN120528T | KC615T | KCTC 9933T |
|---|---|---|---|
| Spores (μm) | 1.1–1.2 | 0.6–0.9 | 1.0–1.4 |
| Growth conditions | |||
| Temperature range (°C) | 20–45 | 28–37 | 20–50 |
| pH range | 6.0–7.0 | 6.0–8.0 | 6.0–9.0 |
| NaCl tolerance (%) | 0–1 | 0–1 | 0–2 |
| Degradation of | |||
| Starch | – | – | + |
| Gelatin | – | – | – |
| Tween 40 | + | + | – |
| Tween 80 | + | + | – |
| Carbon utilisation | |||
| D-Galactose | + | – | – |
| D-Mannose | + | – | – |
| D-Raffinose | – | + | – |
| Adonitol | – | + | – |
| Nitrogen utilisation | |||
| L-Alanine | – | + | – |
| L-Arginine | – | + | – |
| L-Asparagine | – | – | + |
| L-Cysteine | + | + | – |
| L-Methionine | – | – | + |
| L-Tyrosine | + | – | + |
| L-Valine | – | + | – |
| Enzymatic assay | |||
| Arbutin hydrolysis | – | + | – |
| β-glucosidase | + | – | – |
| Major polar lipids * | DPG, PE, PG, 3AL, 4L | DPG, PE, OH-PE, AL, GL, L | DPG, PE, PG, PME, APL, 4AL, 2L |
| Fatty acid (%) | AN120528T | KC615T | KCTC 9933T |
|---|---|---|---|
| Saturated | |||
| C13:0 | tr | - | - |
| C14:0 | 1.1 | 1.4 | 1.4 |
| C16:0 | 4.9 | 1.7 | 6.3 |
| C17:0 | 1.4 | - | tr |
| C18:0 | 1.1 | 10.1 | - |
| C19:0 | 1.1 | 1.1 | - |
| C20:0 | - | 15.8 | - |
| Branched | |||
| iso C13:0 | tr | - | tr |
| iso C14:0 | 9.5 | 5.2 | 5.5 |
| iso C15:0 | 31.8 | 6.2 | 13.2 |
| iso C16:0 | 9.3 | 2.4 | 4.4 |
| iso C17:0 | 1.5 | 2.7 | 1.1 |
| iso C18:0 | - | 2.5 | - |
| iso C19:0 | - | 4.9 | - |
| iso C17:1 ω10c | tr | - | |
| anteiso-C13:0 | tr | - | - |
| anteiso-C15:0 | 32.3 | 37.91 | 59.1 |
| anteiso-C17:0 | 1.5 | 5.1 | 3.2 |
| anteiso-C19:0 | - | 3.3 | - |
| C16:1 ω11c | 1.8 | - | 3.4 |
| C16:1 ω7c alcohol | 1.9 | - | 5.5 |
| C18:1 ω9c | Tr | - | - |
| Summed feature 4 | - | - | tr |
| Features | AN120528T | KC615T | KCTC 9933T |
|---|---|---|---|
| Genome size (bp) | 3,371,008 | 3,989,583 | 4,185,101 |
| Contigs | 25 | 44 | 42 |
| N50 (bp) | 408,672 | 278,159 | 244,454 |
| L50 | contig 3 | contig 6 | contig 6 |
| Total genes | 3548 | 4054 | 4273 |
| Pseudogene | 74 | 203 | 109 |
| CDSs | 3408 | 3796 | 4087 |
| rRNA | 10 | 3 | 20 |
| tRNA | 52 | 48 | 53 |
| G+C content (%) | 39.0 | 38.5 | 38.4 |
| Antibiotic resistance gene clusters | 1 (glycopeptide resistance) | 3 (one glycopeptide resistance and two antibiotic efflux) | 1 (glycopeptide resistance) |
| CRISPR system (Number of spacers) | Type III-B (13) | Type I-C (30) | Type I-C (17) |
| Number of BGCs | 10 | 11 | 16 |
| In the Genome of AN120528T | NCBI Blast | Putative | ||||
|---|---|---|---|---|---|---|
| Protein ID | Locus (Contig 1) | Annotation | Description (Accession No.) | Scientific Name | Identity (%) | Protein (Functions) |
| 00326 | 295741_ 295869 | hypothetical protein | hypothetical protein PPOP_1752 (GAC42395.1) | Paenibacillus popilliae ATCC 14706 | 47.06 | ShiA (precursor peptide) |
| 00327 | 295926_ 296402 | hypothetical protein | lasso peptide biosynthesis protein (WP_028776449.1) | S. kribbensis KCTC 9933T | 67.09 | ShiB2 (B2 element; protease) |
| 00328 | 296415_ 298103 | hypothetical protein | asparagine synthase-related protein (WP_028776448.1) | S. kribbensis KCTC 9933T | 65.54 | ShiC (cyclase) |
| 00329 | 298066_ 298338 | hypothetical protein | hypothetical protein (WP_028776447.1) | S. kribbensis KCTC 9933T | 60.70 | ShiB1 (B1 element; RRE) |
| 00330 | 298360_ 300078 | putative ATP-binding protein | ABC transporter ATP-binding protein/ permease (WP_028776446.1) | S. kribbensis KCTC 9933T | 74.17 | ShiD (ABC transporter) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, C.-Z.; Lee, J.M.; Kim, C.-J.; Lee, H.-G.; Shin, K.-S. Genomic Insight into Shimazuella Soli Sp. Nov. Isolated from Soil and Its Putative Novel Class II Lasso Peptide. Bioengineering 2022, 9, 812. https://doi.org/10.3390/bioengineering9120812
Jin C-Z, Lee JM, Kim C-J, Lee H-G, Shin K-S. Genomic Insight into Shimazuella Soli Sp. Nov. Isolated from Soil and Its Putative Novel Class II Lasso Peptide. Bioengineering. 2022; 9(12):812. https://doi.org/10.3390/bioengineering9120812
Chicago/Turabian StyleJin, Chun-Zhi, Jong Min Lee, Chang-Jin Kim, Hyung-Gwan Lee, and Kee-Sun Shin. 2022. "Genomic Insight into Shimazuella Soli Sp. Nov. Isolated from Soil and Its Putative Novel Class II Lasso Peptide" Bioengineering 9, no. 12: 812. https://doi.org/10.3390/bioengineering9120812