
Bioprocess Optimization for the Production of Arthrospira (Spirulina) platensis Biomass Enriched in the Enzyme Alkaline Phosphatase


Abstract: Background
1. Introduction
2. Materials and Methods
2.1. Microorganism and Cultivation Conditions
2.2. Experimental Design
2.3. Analytical Methods
2.4. Statistical Analysis
3. Results and Discussion
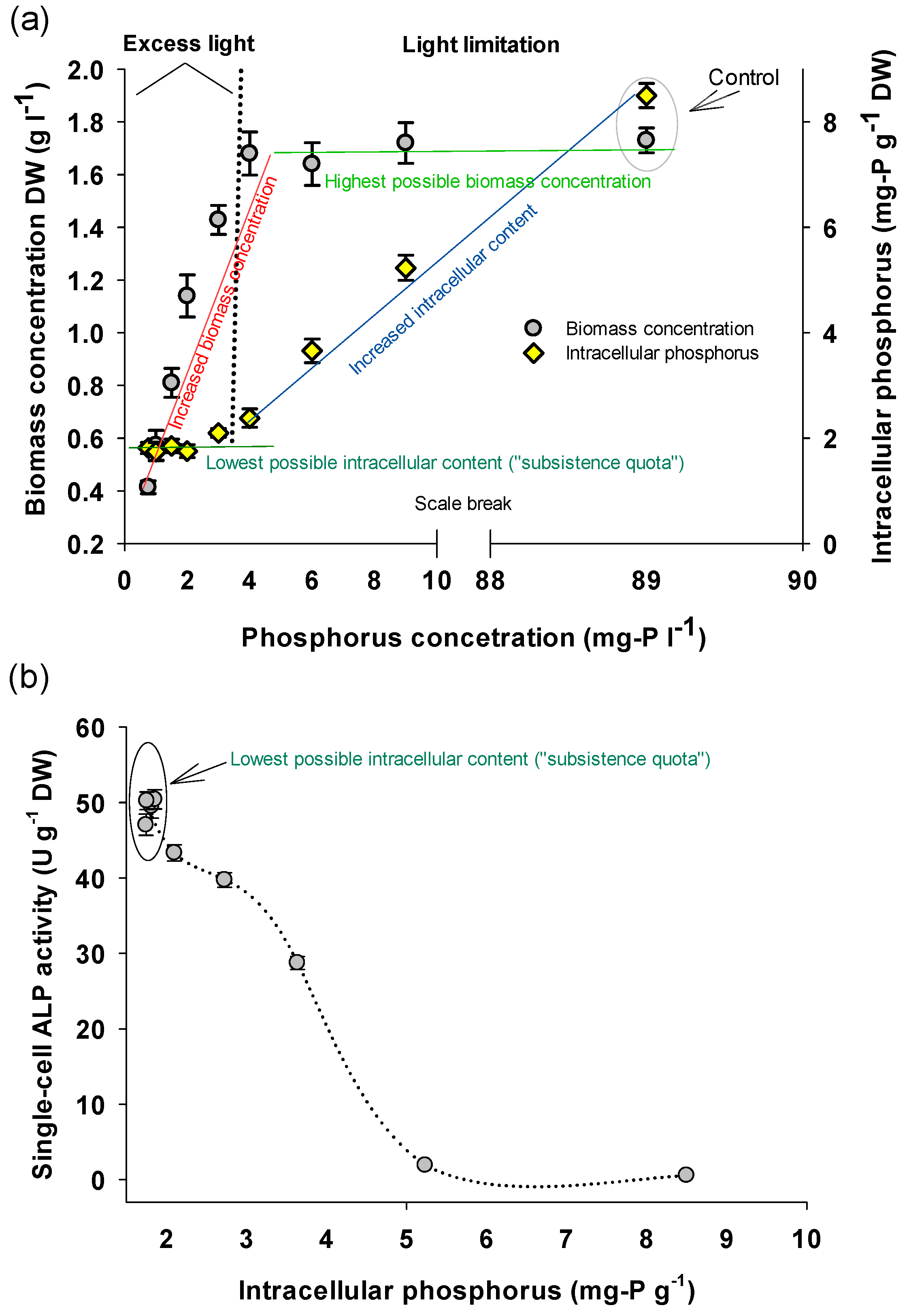
3.1. Phosphorus Concentration Effect on Biomass Production and Biochemical Composition
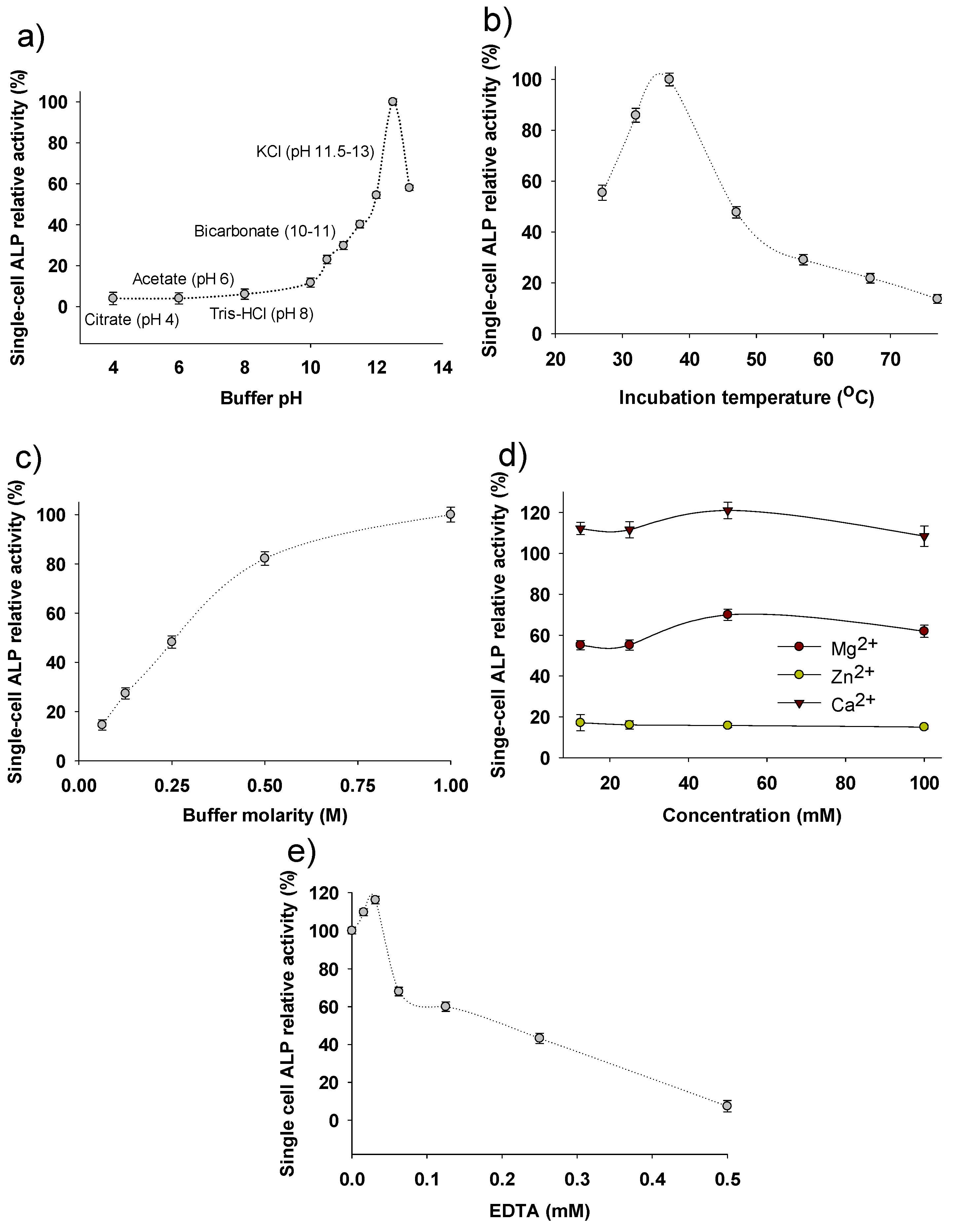
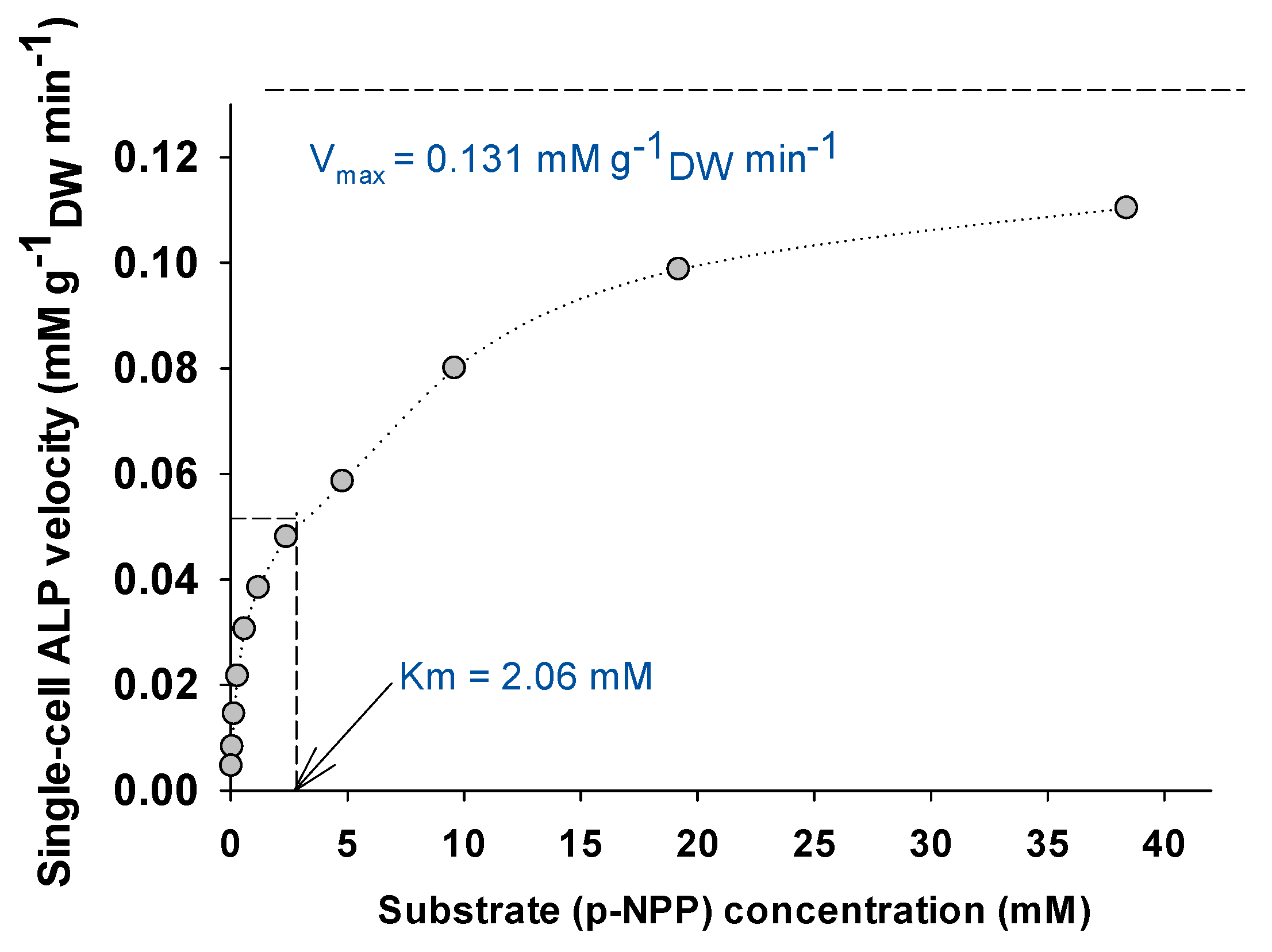
3.2. Parameters Affecting the Enzymatic Activity of Single-Cell Alkaline Phosphatase
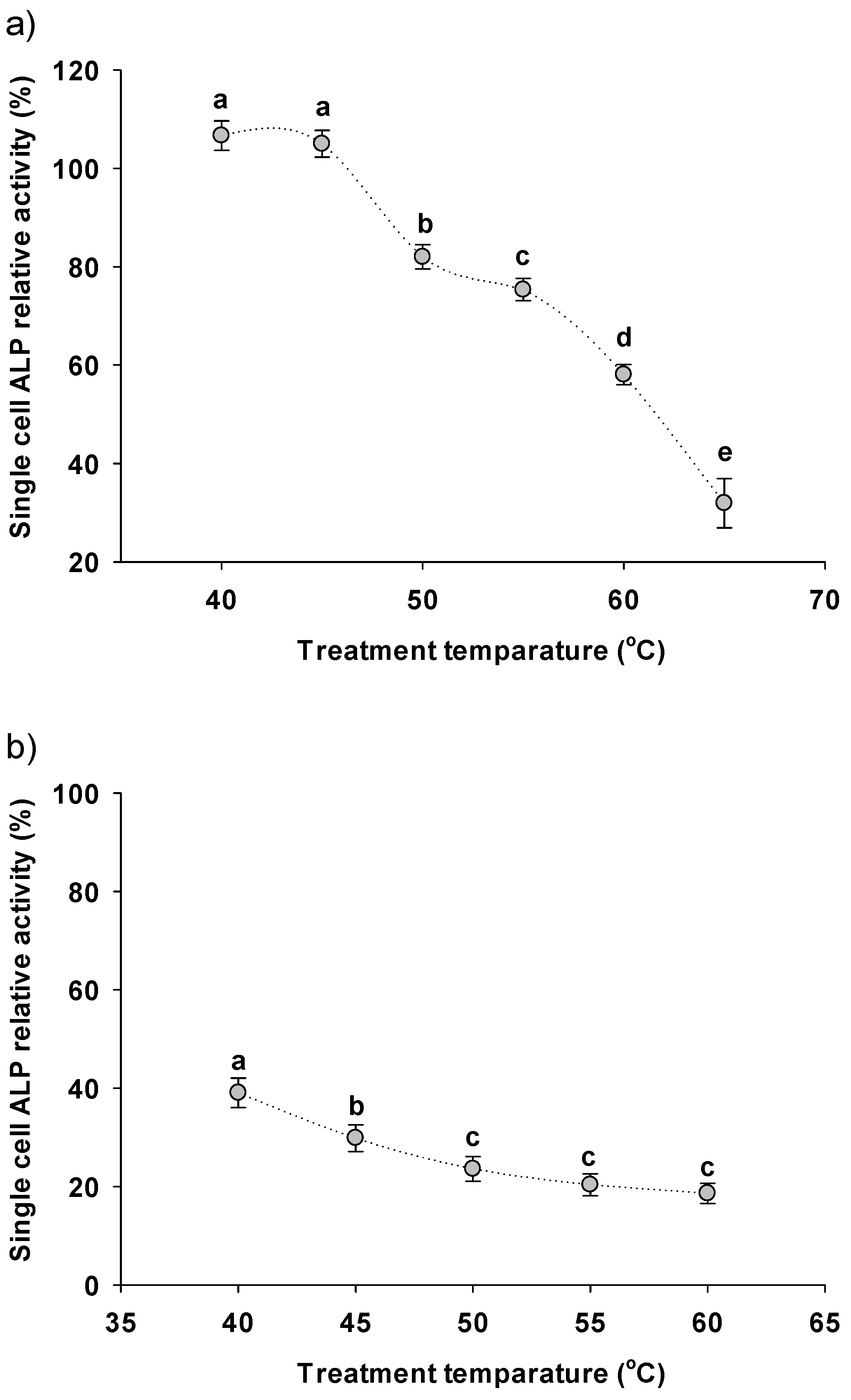
3.3. Downstream Treatments Affecting the Single-Cell ALP Activity
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Green, M.R.; Sambrook, J. Alkaline Phosphatase. Cold Spring Harb. Protoc. 2020, 2020, 100768. [Google Scholar] [CrossRef]
- Zhao, D.; Li, J.; Peng, C.; Zhu, S.; Sun, J.; Yang, X. Fluorescence Immunoassay Based on the Alkaline Phosphatase Triggered In Situ Fluorogenic Reaction of o-Phenylenediamine and Ascorbic Acid. Anal. Chem. 2019, 91, 2978–2984. [Google Scholar] [CrossRef] [PubMed]
- Meng, D.; Jiang, W.; Li, J.; Huang, L.; Zhai, L.; Zhang, L.; Guan, Z.; Cai, Y.; Liao, X. An Alkaline Phosphatase from Bacillus amyloliquefaciens YP6 of New Application in Biodegradation of Five Broad-Spectrum Organophosphorus Pesticides. J. Environ. Sci. Health Part B 2019, 54, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Moya, P.; Ortega-González, M.; González, R.; Anzola, A.; Ocón, B.; Hernández-Chirlaque, C.; López-Posadas, R.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; et al. Exogenous Alkaline Phosphatase Treatment Complements Endogenous Enzyme Protection in Colonic Inflammation and Reduces Bacterial Translocation in Rats. Pharmacol. Res. 2012, 66, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Haarhaus, M.; Brandenburg, V.; Kalantar-Zadeh, K.; Stenvinkel, P.; Magnusson, P. Alkaline Phosphatase: A Novel Treatment Target for Cardiovascular Disease in CKD. Nat. Rev. Nephrol. 2017, 13, 429. [Google Scholar] [CrossRef]
- Fawley, J.; Gourlay, D.M. Intestinal Alkaline Phosphatase: A Summary of its Role in Clinical Disease. J. Surg. Res. 2016, 202, 225–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallès, J.-P. Recent Advances in Intestinal Alkaline Phosphatase, Inflammation, and Nutrition. Nutr. Rev. 2019, 77, 710–724. [Google Scholar] [CrossRef]
- Kühn, F.; Adiliaghdam, F.; Cavallaro, P.M.; Hamarneh, S.R.; Tsurumi, A.; Hoda, R.S.; Munoz, A.R.; Dhole, Y.; Ramirez, J.M.; Liu, E. Intestinal Alkaline Phosphatase Targets the Gut Barrier to Prevent Aging. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Osathanon, T.; Giachelli, C.M.; Somerman, M.J. Immobilization of Alkaline Phosphatase on Microporous Nanofibrous Fibrin Scaffolds for Bone Tissue Engineering. Biomaterials 2009, 30, 4513–4521. [Google Scholar] [CrossRef] [Green Version]
- Torres-Tiji, Y.; Fields, F.J.; Mayfield, S.P. Microalgae as a Future Food Source. Biotechnol. Adv. 2020, 41, 107536. [Google Scholar] [CrossRef]
- Markou, G.; Eliopoulos, C.; Argyri, A.; Arapoglou, D. Production of Arthrospira (Spirulina) platensis Enriched in β-Glucans through Phosphorus Limitation. Appl. Sci. 2021, 11, 8121. [Google Scholar] [CrossRef]
- Bernaerts, T.M.; Gheysen, L.; Kyomugasho, C.; Kermani, Z.J.; Vandionant, S.; Foubert, I.; Hendrickx, M.E.; Van Loey, A.M. Comparison of Microalgal Biomasses as Functional Food Ingredients: Focus on the Composition of Cell Wall Related Polysaccharides. Algal Res. 2018, 32, 150–161. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Schulze, P.S.; Kiron, V.; Wijffels, R.H. Production of Carbohydrates, Lipids and Polyunsaturated Fatty Acids (PUFA) by the Polar Marine Microalga Chlamydomonas malina RCC2488. Algal Res. 2020, 50, 102016. [Google Scholar] [CrossRef]
- Barta, D.G.; Coman, V.; Vodnar, D.C. Microalgae as Sources of Omega-3 Polyunsaturated Fatty Acids: Biotechnological Aspects. Algal Res. 2021, 58, 102410. [Google Scholar] [CrossRef]
- Sansone, C.; Brunet, C. Promises and Challenges of Microalgal Antioxidant Production. Antioxidants 2019, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, M.; Senhorinho, G.; Scott, J. Microalgae under Environmental Stress as a Source of Antioxidants. Algal Res. 2020, 52, 102104. [Google Scholar] [CrossRef]
- Edelmann, M.; Aalto, S.; Chamlagain, B.; Kariluoto, S.; Piironen, V. Riboflavin, Niacin, Folate and Vitamin B12 in Commercial Microalgae Powders. J. Food Compos. Anal. 2019, 82, 103226. [Google Scholar] [CrossRef]
- Del Mondo, A.; Smerilli, A.; Sané, E.; Sansone, C.; Brunet, C. Challenging Microalgal Vitamins for Human Health. Microb. Cell Fact. 2020, 19, 1–23. [Google Scholar] [CrossRef]
- Ovando, C.A.; Carvalho, J.C.D.; de Melo Pereira, G.V.; Jacques, P.; Soccol, V.T.; Soccol, C.R. Functional Properties and Health Benefits of Bioactive Peptides Derived from Spirulina: A Review. Food Rev. Int. 2018, 34, 34–51. [Google Scholar] [CrossRef]
- Çelekli, A.; Alslibi, Z.A.; Bozkurt, H. Influence of Incorporated Spirulina Platensis on the Growth of Microflora and Physicochemical Properties of Ayran as a Functional Food. Algal Res. 2019, 44, 101710. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Brück, W.M.; Brück, T.; Beyrer, M. Microalgae as Healthy Ingredients for Functional Foods. In The Role of Alternative and Innovative Food Ingredients and Products in Consumer Wellness; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 103–137. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Dubinsky, Z.; Verdelho, V.; Iluz, D. Unconventional High-Value Products from Microalgae: A Review. Bioresour. Technol. 2021, 329, 124895. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, N.K.M.; Ghazy, E.W.; Abdel-Daim, M.M. Pharmacodynamic Interaction of Spirulina Platensis and Deltamethrin in Freshwater Fish Nile Tilapia, Oreochromis niloticus: Impact on Lipid Peroxidation and Oxidative Stress. Environ. Sci. Pollut. Res. 2015, 22, 3023–3031. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Dawood, M.A.; Elbadawy, M.; Aleya, L.; Alkahtani, S. Spirulina Platensis Reduced Oxidative Damage Induced by Chlorpyrifos Toxicity in Nile Tilapia (Oreochromis niloticus). Animals 2020, 10, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vingiani, G.M.; De Luca, P.; Ianora, A.; Dobson, A.D.; Lauritano, C. Microalgal Enzymes with Biotechnological Applications. Mar. Drugs 2019, 17, 459. [Google Scholar] [CrossRef] [Green Version]
- Elleuch, J.; Hadj Kacem, F.; Ben Amor, F.; Hadrich, B.; Michaud, P.; Fendri, I.; Abdelkafi, S. Extracellular Neutral Protease from Arthrospira platensis: Production, Optimization and Partial Characterization. Int. J. Biol. Macromol. 2021, 167, 1491–1498. [Google Scholar] [CrossRef]
- Brasil, B.d.S.A.F.; de Siqueira, F.G.; Salum, T.F.C.; Zanette, C.M.; Spier, M.R. Microalgae and Cyanobacteria as Enzyme Biofactories. Algal Res. 2017, 25, 76–89. [Google Scholar] [CrossRef]
- Sharifian, S.; Homaei, A.; Kim, S.-K.; Satari, M. Production of Newfound Alkaline Phosphatases from Marine Organisms with Potential Functions and Industrial Applications. Process. Biochem. 2018, 64, 103–115. [Google Scholar] [CrossRef]
- Chu, Y.-H.; Yu, X.-X.; Jin, X.; Wang, Y.-T.; Zhao, D.-J.; Zhang, P.; Sun, G.-M.; Zhang, Y.-H. Purification and Characterization of Alkaline Phosphatase from Lactic Acid Bacteria. RSC Adv. 2019, 9, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Shao, W.; Ebaid, R.; El-Sheekh, M.; Abomohra, A.; Eladel, H. Pharmaceutical Applications and Consequent Environmental Impacts of Spirulina (Arthrospira): An Overview. Grasas Aceites 2019, 70, e292. [Google Scholar] [CrossRef] [Green Version]
- Lafarga, T.; Fernández-Sevilla, J.M.; González-López, C.; Acién-Fernández, F.G. Spirulina for the Food and Functional Food Industries. Food Res. Int. 2020, 137, 109356. [Google Scholar] [CrossRef]
- Almeida, L.M.R.; da Silva Cruz, L.F.; Machado, B.A.S.; Nunes, I.L.; Costa, J.A.V.; de Souza Ferreira, E.; Lemos, P.V.F.; Druzian, J.I.; de Souza, C.O. Effect of the Addition of Spirulina Sp. Biomass on the Development and Characterization of Functional Food. Algal Res. 2021, 58, 102387. [Google Scholar] [CrossRef]
- Şahin, O.I. Functional and Sensorial Properties of Cookies Enriched with Spirulina and Dunaliella Biomass. J. Food Sci. Technol. 2020, 57, 3639–3646. [Google Scholar] [CrossRef]
- Casciano, F.; Nissen, L.; Gianotti, A. Effect of Formulations and Fermentation Processes on Volatile Organic Compounds and Prebiotic Potential of Gluten-Free Bread Fortified by Spirulina (Arthrospira platensis). Food Funct. 2021. [Google Scholar] [CrossRef] [PubMed]
- Padalino, L.; Lecce, L.; Chini Zittelli, G.; Lo Grieco, A.; Yorzillo, G. Use of Spirulina to Enhance the Nutritional Value of Durum Wheat Spaghetti. Food Nutr. J. 2017, 2, 122. [Google Scholar]
- Bosnea, L.; Terpou, A.; Pappa, E.; Kondyli, E.; Mataragas, M.; Markou, G.; Katsaros, G. Incorporation of Spirulina Platensis on Traditional Greek Soft Cheese with Respect to Its Nutritional and Sensory Perspectives. Proceedings 2020, 70, 99. [Google Scholar] [CrossRef]
- Galafat, A.; Vizcaíno, A.J.; Sáez, M.I.; Martínez, T.F.; Jérez-Cepa, I.; Mancera, J.M.; Alarcón, F.J. Evaluation of Arthrospira sp. Enzyme Hydrolysate as Dietary Additive in Gilthead Seabream (Sparus aurata) Juveniles. J. Appl. Phycol. 2020, 32, 3089–3100. [Google Scholar] [CrossRef]
- Khan, S.; Mobashar, M.; Mahsood, F.K.; Javaid, S.; Abdel-Wareth, A.; Ammanullah, H.; Mahmood, A. Spirulina Inclusion Levels in a Broiler Ration: Evaluation of Growth Performance, Gut Integrity, and Immunity. Trop. Anim. Health Prod. 2020, 52, 3233–3240. [Google Scholar] [CrossRef]
- Martins, C.F.; Pestana Assuncao, J.; Ribeiro Santos, D.M.; Madeira, M.S.M.d.S.; Alfaia, C.M.R.P.M.; Lopes, P.A.A.B.; Coelho, D.F.M.; Cardoso Lemos, J.P.; de Almeida, A.M.; Mestre Prates, J.A. Effect of Dietary Inclusion of Spirulina on Production Performance, Nutrient Digestibility and Meat Quality Traits in Post-Weaning Piglets. J. Anim. Physiol. Anim. Nutr. 2021, 105, 247–259. [Google Scholar] [CrossRef]
- Markou, G. Alteration of the Biomass Composition of Arthrospira (Spirulina) platensis under Various Amounts of Limited Phosphorus. Bioresour. Technol. 2012, 116, 533–535. [Google Scholar] [CrossRef]
- Quisel, J.D.; Wykoff, D.D.; Grossman, A.R. Biochemical Characterization of the Extracellular Phosphatases Produced by Phosphorus-Deprived Chlamydomonas reinhardtii. Plant Physiol. 1996, 111, 839–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golotin, V.; Balabanova, L.; Likhatskaya, G.; Rasskazov, V. Recombinant Production and Characterization of a Highly Active Alkaline Phosphatase from Marine Bacterium Cobetia marina. Mar. Biotechnol. 2015, 17, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Droop, M.R. Vitamin B12 and Marine Ecology. IV. The Kinetics of Uptake, Growth and Inhibition in Monochrysis lutheri. J. Mar. Biol. Assoc. 1968, 48, 689–733. [Google Scholar] [CrossRef]
- Carvalho, A.; Silva, S.; Baptista, J.; Malcata, F. Light Requirements in Microalgal Photobioreactors: An Overview of Biophotonic Aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef]
- Adams, C.; Godfrey, V.; Wahlen, B.; Seefeldt, L.; Bugbee, B. Understanding Precision Nitrogen Stress to Optimize the Growth and Lipid Content Tradeoff in Oleaginous Green Microalgae. Bioresour. Technol. 2013, 131, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Ghyoot, C.; Gypens, N.; Flynn, K.J.; Lancelot, C. Modelling Alkaline Phosphatase Activity in Microalgae under Orthophosphate Limitation: The Case of Phaeocystis globosa. J. Plankton Res. 2015, 37, 869–885. [Google Scholar] [CrossRef] [Green Version]
- Whitton, B.A.; Al-Shehri, A.M.; Ellwood, N.T.; Turner, B.L. Ecological Aspects of Phosphatase Activity in Cyanobacteria, Eukaryotic Algae and Bryophytes. In Organic Phosphorus in the Environment; CAB International: Wallingford, UK, 2005; pp. 205–241. [Google Scholar]
- Girault, M.; Siano, R.; Labry, C.; Latimier, M.; Jauzein, C.; Beneyton, T.; Buisson, L.; Del Amo, Y.; Baret, J.-C. Variable Inter and Intraspecies Alkaline Phosphatase Activity within Single Cells of Revived Dinoflagellates. ISME J. 2021, 15, 2057–2069. [Google Scholar] [CrossRef]
- Mo, Y.; Ou, L.; Lin, L.; Huang, B. Temporal and Spatial Variations of Alkaline Phosphatase Activity Related to Phosphorus Status of Phytoplankton in the East China Sea. Sci. Total Environ. 2020, 731, 139192. [Google Scholar] [CrossRef]
- Pandey, S.; Banik, R. Optimization of Process Parameters for Alkaline Phosphatase Production by Bacillus licheniformis using Response Surface Methodology. Int. J. Agric. Technol. 2010, 6, 721–732. [Google Scholar]
- Zhou, Y.; Lu, Z.; Wang, X.; Selvaraj, J.N.; Zhang, G. Genetic Engineering Modification and Fermentation Optimization for Extracellular Production of Recombinant Proteins using Escherichia coli. Appl. Microbiol. Biotechnol. 2018, 102, 1545–1556. [Google Scholar] [CrossRef]
- Sebastian, M.; Ammerman, J.W. The Alkaline Phosphatase PhoX is More Widely Distributed in Marine Bacteria Than the Classical PhoA. ISME J. 2009, 3, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.E.; Kantrowitz, E.R. Why are Mammalian Alkaline Phosphatases Much More Active than Bacterial Alkaline Phosphatases? Mol. Microbiol. 1994, 12, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Thengodkar, R.R.M.; Sivakami, S. Degradation of Chlorpyrifos by an Alkaline Phosphatase from the Cyanobacterium Spirulina Platensis. Biodegradation 2010, 21, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Asencio, A.D.; Morte, A.; García-Carmona, F.; Pérez-Gilabert, M. Partial Purification and Characterization of a Calcium-Dependent Alkaline Phosphatase from the Cyanobacterium Arthrospira platensis. J. Phycol. 2012, 48, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, S.; Pandey, V.; Mishra, A. Factors Modulating Alkaline Phosphatase Activity in the Diazotrophic Rice-field Cyanobacterium, Anabaena oryzae. World J. Microbiol. Biotechnol. 2006, 22, 927–935. [Google Scholar] [CrossRef]
- Upadhyay, L.S.B.; Verma, N. A Three Step Approach for the Purification of Alkaline Phosphatase from Non-Pasteurized Milk. J. Food Sci. Technol. 2015, 52, 3140–3146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plocke, D.J.; Levinthal, C.; Vallee, B.L. Alkaline Phosphatase of Escherichia coli: A Zinc Metalloenzyme. Biochemistry 1962, 1, 373–378. [Google Scholar] [CrossRef]
- McComb, R.B.; Bowers, G.N., Jr.; Posen, S. Alkaline Phosphatase; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Lopez, A.; Pique, M.; Boatella, J.; Parcerisa, J.; Romero, A.; Ferrá, A.; Garcí, J. Influence of Drying Conditions on the Hazelnut Quality. II. Enzymatic Activity. Dry Technol. 1997, 15, 979–988. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P concentration (mg-P L−1) | Intracellular P (mg-P g−1) | Proteins %DW) | Carbohydrates (%DW) | Phycocyanin (%DW) | Lipids (%DW) |
|---|---|---|---|---|---|
| 0.75 | 1.81 | 23.55 ± 2.37 a | 65.66 ± 7.55 a | 1.52 ± 0.23 a | 6.53 ± 1.23 abc |
| 1 | 1.74 | 22.67 ± 1.65 a | 64.24 ± 3.44 a | 1.49 ± 0.32 ab | 6.14 ± 0.69 ac |
| 1.5 | 1.85 | 23.88 ± 2.14 a | 66.12 ± 2.78 a | 1.67± 0.12 a | 5.98 ± 1.11 ac |
| 2 | 1.75 | 22.44 ± 2.98 a | 65.46 ± 6.79 a | 1.22 ± 0.23 b | 6.99 ± 1.36 abc |
| 3 | 2.10 | 25.39 ± 2.44 a | 62.99 ± 4.22 a | 2.45 ± 0.55 c | 8.45 ± 0.96 b |
| 4 | 2.73 | 31.49 ± 1.94 b | 42.14 ± 3.47 b | 7.78 ± 1.05 d | 5.44 ± 0.55 c |
| 6 | 3.64 | 45.96 ± 3.59 c | 25.83 ± 2.59 c | 9.98 ± 0.98 e | 7.76 ± 2.01 ab |
| 9 | 5.23 | 55.58 ± 4.17 d | 14.29 ± 1.55 d | 12.32 ± 2.04 f | 7.42 ± 1.87 ab |
| 89 (Control) | 8.55 | 58.48 ± 3.42 d | 12.11 ± 2.84 d | 12.12 ± 1.26 f | 7.29 ± 1.84ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markou, G. Bioprocess Optimization for the Production of Arthrospira (Spirulina) platensis Biomass Enriched in the Enzyme Alkaline Phosphatase. Bioengineering 2021, 8, 142. https://doi.org/10.3390/bioengineering8100142
Markou G. Bioprocess Optimization for the Production of Arthrospira (Spirulina) platensis Biomass Enriched in the Enzyme Alkaline Phosphatase. Bioengineering. 2021; 8(10):142. https://doi.org/10.3390/bioengineering8100142
Chicago/Turabian StyleMarkou, Giorgos. 2021. "Bioprocess Optimization for the Production of Arthrospira (Spirulina) platensis Biomass Enriched in the Enzyme Alkaline Phosphatase" Bioengineering 8, no. 10: 142. https://doi.org/10.3390/bioengineering8100142