The Macro- and Micro-Mechanics of the Colon and Rectum II: Theoretical and Computational Methods



Abstract
:1. Introduction
2. Modeling the Colorectum: State of the Art
2.1. Macro-Scale: Biomechanics of the Colorectum
2.2. Micro-Scale: Mechanotransduction of the Colorectum
3. Towards Coupled Multi-Scale Modeling of the Colorectum
3.1. Constitutive Models of the Biomechanics
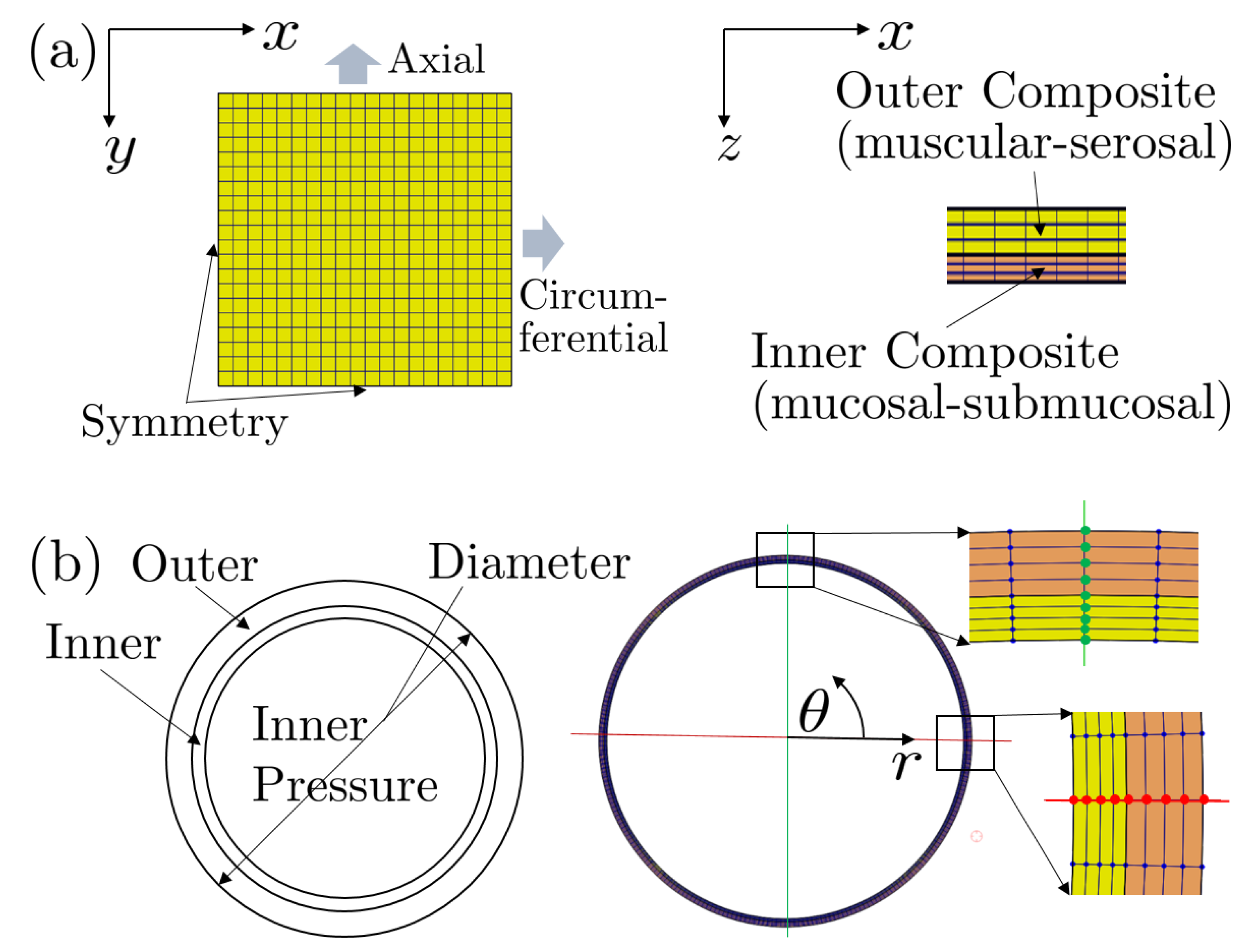
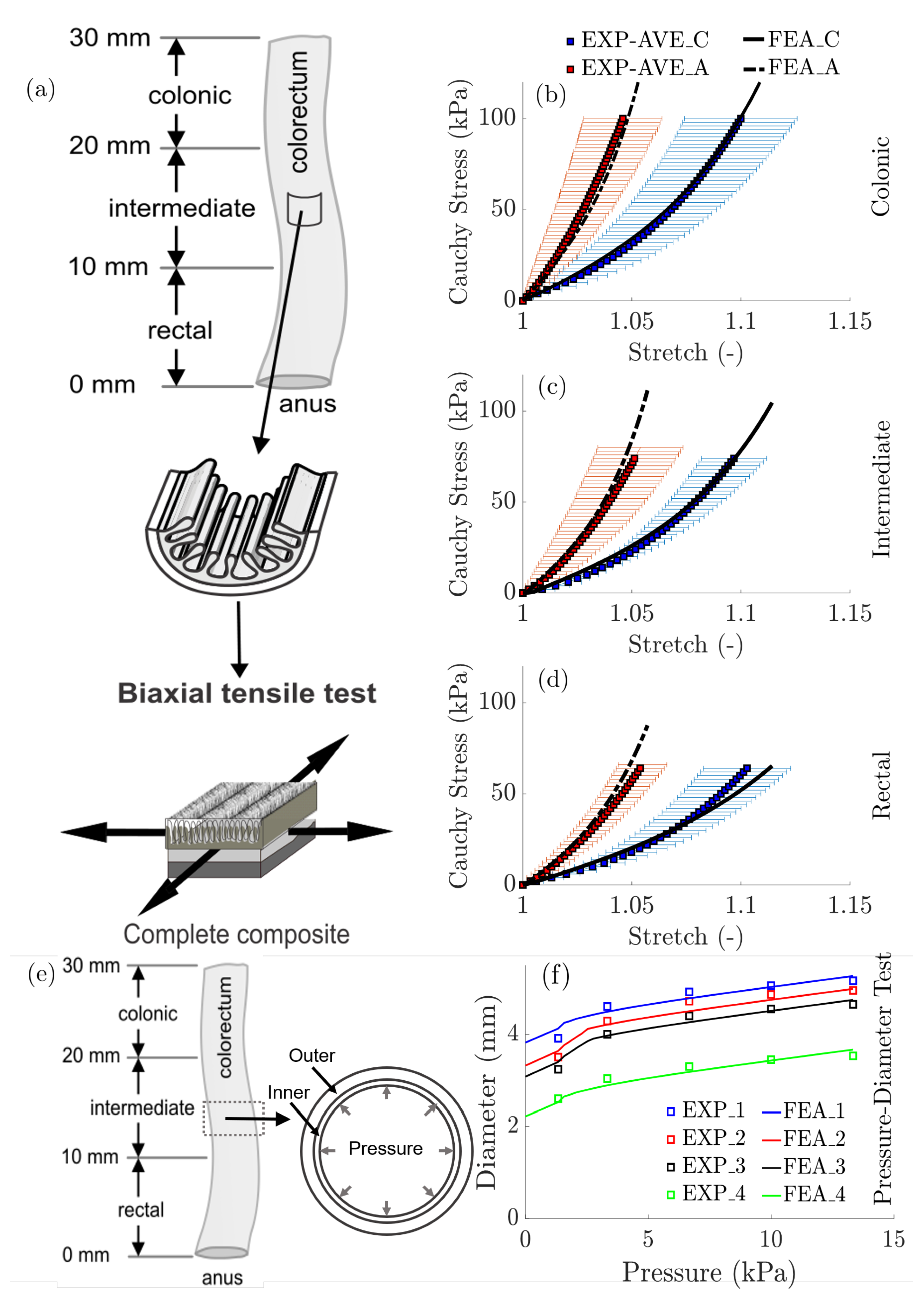
3.2. Biomechanics: Computational Modeling and Results
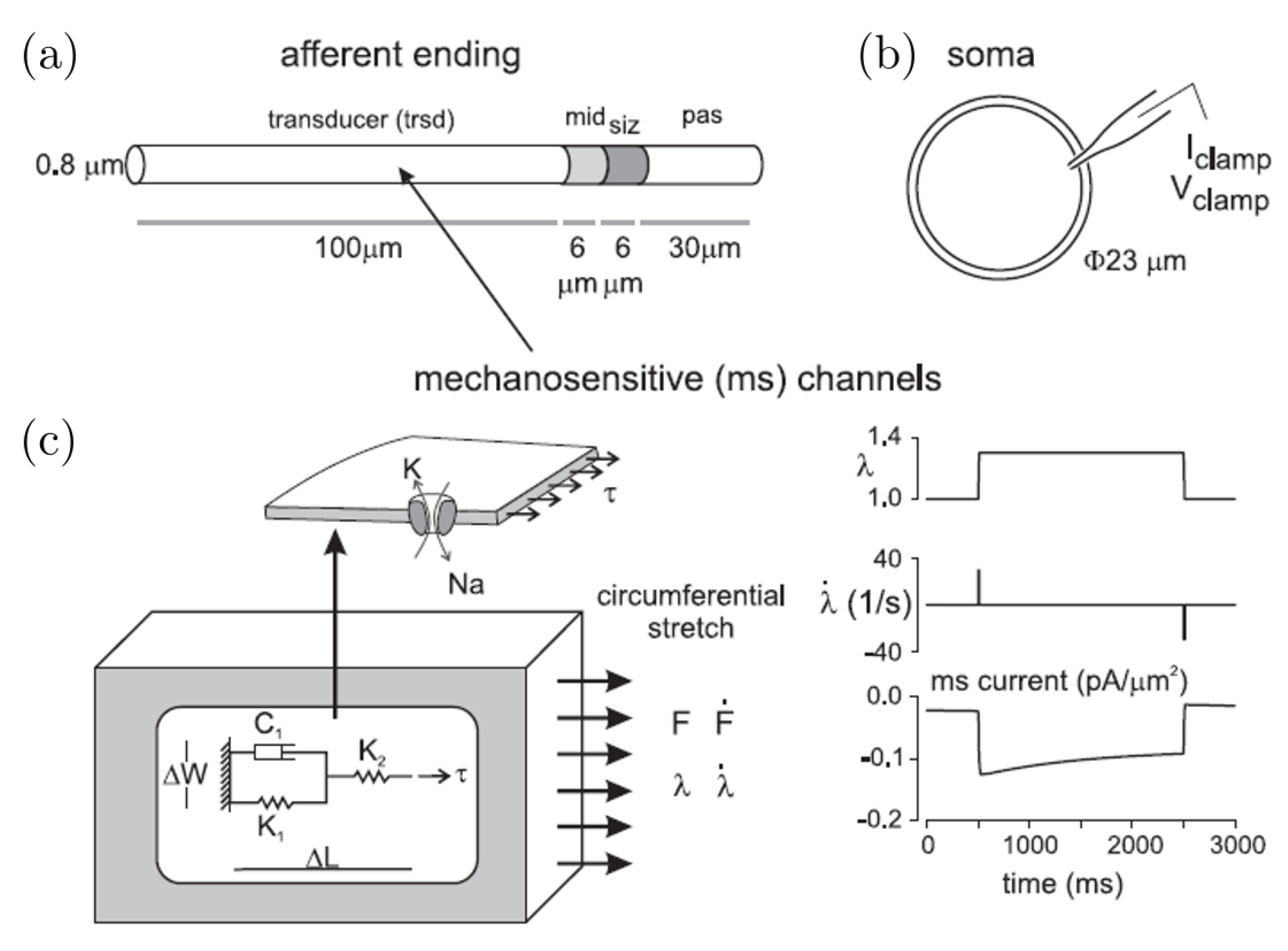
3.3. Theoretical Models of Stretch-Sensitive Afferent Endings
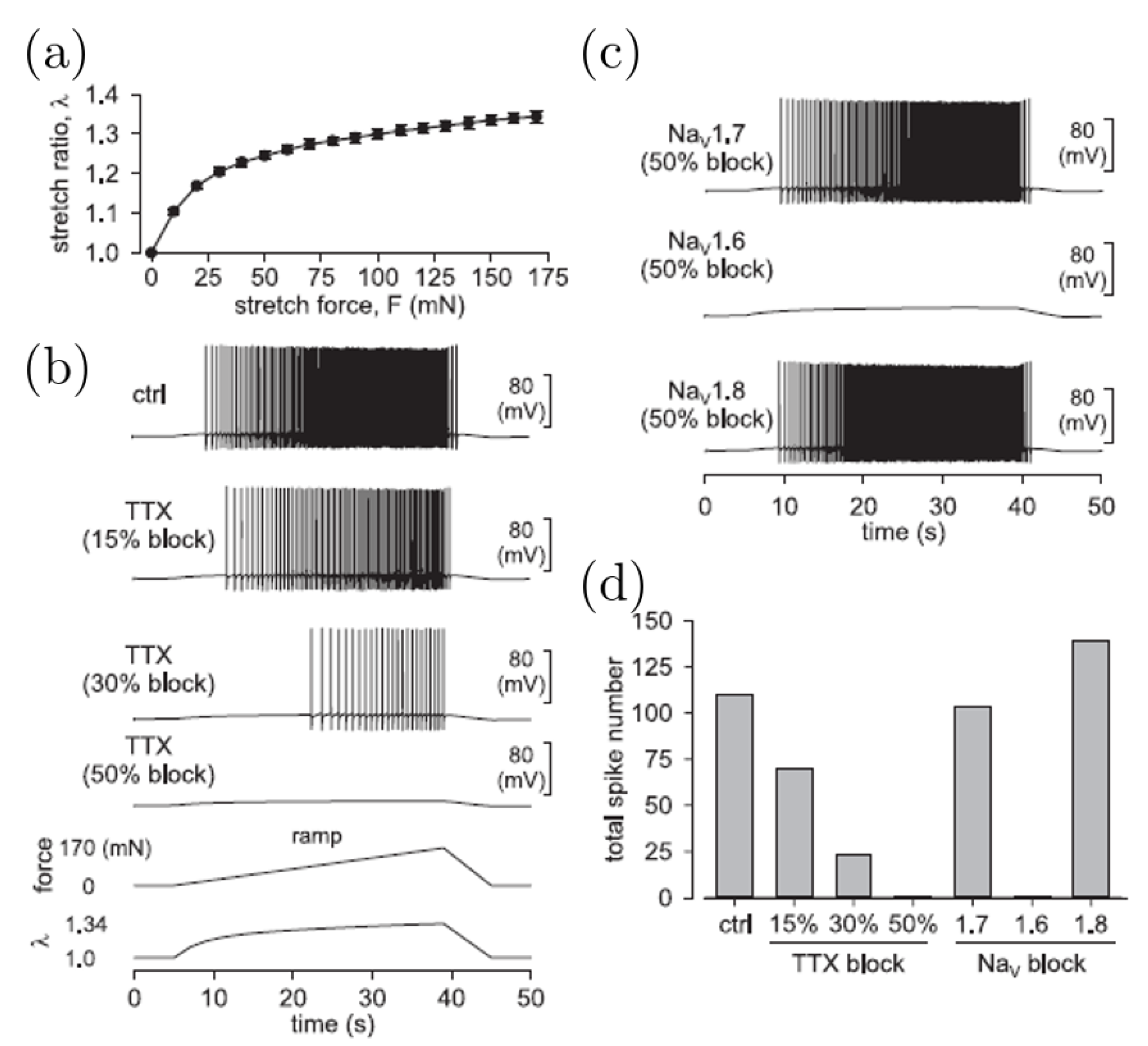
3.4. Mechanotransduction: Computational Models and Results on Stretch-Sensitive Afferent Endings
4. Discussion and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Xavier, R.; Podolsky, D. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liao, D.; Gregersen, H. Mechanical analysis of intestinal contractility in a neonatal maternal deprivation irritable bowel syndrome rat model. J. Biomech. 2019, 93, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Stollman, N.; Raskin, J.B. Diverticular disease of the colon. Lancet 2004, 363, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Bharucha, A.E.; Rao, S.S. An update on anorectal disorders for gastroenterologists. Gastroenterology 2014, 146, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, N.T.; Kalff, J.C.; Türler, A.; Engel, B.M.; Watkins, S.C.; Billiar, T.R.; Bauer, A.J. Prostanoid production via COX-2 as a causative mechanism of rodent postoperative ileus. Gastroenterology 2001, 121, 1354–1371. [Google Scholar] [CrossRef]
- Hughes, P.; Brierley, S.; Blackshaw, L. Post-inflammatory modification of colonic afferent mechanosensitivity. Clin. Exp. Pharmacol. Physiol. 2009, 36, 1034–1040. [Google Scholar] [CrossRef]
- Siri, S.; Maier, F.; Santos, S.; Pierce, D.M.; Feng, B. Load-bearing function of the colorectal submucosa and its relevance to visceral nociception elicited by mechanical stretch. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G349–G358. [Google Scholar] [CrossRef]
- Siri, S.; Zhao, Y.; Maier, F.; Pierce, D.M.; Feng, B. The Macro-and Micro-Mechanics of the Colon and Rectum I: Experimental Evidence. Bioengineering 2020, 7, 130. [Google Scholar] [CrossRef]
- Carniel, E.; Gramigna, V.; Fontanella, C.; Frigo, A.; Stefanini, C.; Rubini, A.; Natali, A. Characterization of the anisotropic mechanical behaviour of colonic tissues: Experimental activity and constitutive formulation. Exp. Physiol. 2014, 99, 759–771. [Google Scholar] [CrossRef]
- Siri, S.; Maier, F.; Chen, L.; Santos, S.; Pierce, D.M.; Feng, B. Differential biomechanical properties of mouse distal colon and rectum innervated by the splanchnic and pelvic afferents. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G473–G481. [Google Scholar] [CrossRef]
- Bellini, C.; Glass, P.; Sitti, M.; Di Martino, E.S. Biaxial mechanical modeling of the small intestine. J. Mech. Behav. Biomed. Mater. 2011, 4, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Sokolis, D.P.; Orfanidis, I.K.; Peroulis, M. Biomechanical testing and material characterization for the rat large intestine: Regional dependence of material parameters. Physiol. Meas. 2011, 32, 1969–1982. [Google Scholar] [CrossRef] [PubMed]
- Sokolis, D.P.; Sassani, S.G. Microstructure-based constitutive modeling for the large intestine validated by histological observations. J. Mech. Behav. Biomed. Mater. 2013, 21, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Xu, X.; Lin, S.; Cheng, Y.; Tong, J.; Li, Y. Alterations in biomechanical properties and microstructure of colon wall in early-stage experimental colitis. Exp. Ther. Med. 2017, 14, 995–1000. [Google Scholar] [CrossRef]
- Patel, B.; Chen, H.; Ahuja, A.; Krieger, J.F.; Noblet, J.; Chambers, S.; Kassab, G.S. Constitutive modeling of the passive inflation-extension behavior of the swine colon. J. Mech. Behav. Biomed. Mater. 2018, 77, 176–186. [Google Scholar] [CrossRef]
- Puértolas, S.; Peña, E.; Herrera, A.; Ibarz, E.; Gracia, L. A comparative study of hyperelastic constitutive models for colonic tissue fitted to multiaxial experimental testing. J. Mech. Behav. Biomed. Mater. 2020, 102, 103507. [Google Scholar] [CrossRef]
- Feng, B.; Gebhart, G.F. Characterization of silent afferents in the pelvic and splanchnic innervations of the mouse colorectum. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G170–G180. [Google Scholar] [CrossRef]
- La, J.H.; Gebhart, G.F. Colitis decreases mechanosensitive K2P channel expression and function in mouse colon sensory neurons. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G165–G174. [Google Scholar] [CrossRef] [Green Version]
- Lanir, Y. A structural theory for the homogeneous biaxial stress-strain relationships in flat collagenous tissues. J. Biomech. 1979, 12, 423–436. [Google Scholar] [CrossRef]
- Lanir, Y. Constitutive equations for fibrous connective tissues. J. Biomech. 1983, 16, 1–12. [Google Scholar] [CrossRef]
- Fung, Y.; Fronek, K.; Patitucci, P. Pseudoelasticity of arteries and the choice of its mathematical expression. Am. J. Physiol. Heart Circ. Physiol. 1979, 237, H620–H631. [Google Scholar] [CrossRef] [PubMed]
- Chuong, C.; Fung, Y. Three-dimensional stress distribution in arteries. J. Biomech. Eng. 1983, 105, 268. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, G.; Gasser, T.; Ogden, R. A new constitutive framework for arterial wall mechanics and a comparative study of material models. J. Elasticity 2000, 61, 1–48. [Google Scholar] [CrossRef]
- Ferruzzi, J.; Vorp, D.A.; Humphrey, J. On constitutive descriptors of the biaxial mechanical behaviour of human abdominal aorta and aneurysms. J. R. Soc. Interface 2011, 8, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Liao, D.; Drewes, A.M.; Gregersen, H. Modelling the elastin, collagen and smooth muscle contribution to the duodenal mechanical behaviour in patients with systemic sclerosis. Neurogastroenterol. Motil. 2009, 21, 914-e68. [Google Scholar] [CrossRef]
- Natali, A.N.; Carniel, E.L.; Gregersen, H. Biomechanical behaviour of oesophageal tissues: Material and structural configuration, experimental data and constitutive analysis. Med. Eng. Phys. 2009, 31, 1056–1062. [Google Scholar] [CrossRef]
- Sokolis, D.P. Structurally-motivated characterization of the passive pseudo-elastic response of esophagus and its layers. Comput. Biol. Med. 2013, 43, 1273–1285. [Google Scholar] [CrossRef]
- Higa, M.; Luo, Y.; Okuyama, T.; Shiraishi, Y.; Liu, H.; Yambe, T.; Takagi, T. In vivo measurements and constitutive modeling of colon tissue. In World Congress on Medical Physics and Biomedical Engineering 2006; Springer: Berlin/Heidelberg, Germany, 2007; pp. 3186–3189. [Google Scholar]
- Sokolis, D.P. Experimental study and biomechanical characterization for the passive small intestine: Identification of regional differences. J. Mech. Behav. Biomed. Mater. 2017, 74, 93–105. [Google Scholar] [CrossRef]
- Liao, D.; Zhao, J.; Fan, Y.; Gregersen, H. Two-layered quasi-3D finite element model of the oesophagus. Med. Eng. Phys. 2004, 26, 535–543. [Google Scholar] [CrossRef]
- Ciarletta, P.; Dario, P.; Tendick, F.; Micera, S. Hyperelastic model of anisotropic fiber reinforcements within intestinal walls for applications in medical robotics. Int. J. Rob. Res. 2009, 28, 1279–1288. [Google Scholar] [CrossRef]
- Gasser, T.C.; Ogden, R.W.; Holzapfel, G.A. Hyperelastic modelling of arterial layers with distributed collagen fibre orientations. J. R. Soc. Interface 2006, 3, 15–35. [Google Scholar] [CrossRef] [PubMed]
- Alastrué, V.; Martínez, M.; Doblaré, M.; Menzel, A. Anisotropic micro-sphere-based finite elasticity applied to blood vessel modelling. J. Mech. Phys. Solids 2009, 57, 178–203. [Google Scholar] [CrossRef]
- Alastrué, V.; Sáez, P.; Martínez, M.; Doblaré, M. On the use of the Bingham statistical distribution in microsphere-based constitutive models for arterial tissue. Mech. Res. Commun. 2010, 37, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Pierce, D.M.; Unterberger, M.J.; Trobin, W.; Ricken, T.; Holzapfel, G.A. A microstructurally based continuum model of cartilage viscoelasticity and permeability incorporating measured statistical fiber orientations. Biomech. Model. Mechanobiol. 2016, 15, 229–244. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F.; Katz, B. Measurement of current-voltage relations in the membrane of the giant axon of Loligo. J. Physiol. 1952, 116, 424. [Google Scholar] [CrossRef]
- Raman, I.M.; Bean, B.P. Inactivation and recovery of sodium currents in cerebellar Purkinje neurons: Evidence for two mechanisms. Biophys. J. 2001, 80, 729–737. [Google Scholar] [CrossRef] [Green Version]
- Gurkiewicz, M.; Korngreen, A.; Waxman, S.G.; Lampert, A. Kinetic modeling of Nav1. 7 provides insight into erythromelalgia-associated F1449V mutation. J. Neurophysiol. 2011, 105, 1546–1557. [Google Scholar] [CrossRef] [Green Version]
- Ranade, S.S.; Syeda, R.; Patapoutian, A. Mechanically activated ion channels. Neuron 2015, 87, 1162–1179. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.H.; Ranade, S.; Weyer, A.D.; Dubin, A.E.; Baba, Y.; Qiu, Z.; Petrus, M.; Miyamoto, T.; Reddy, K.; Lumpkin, E.A.; et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature 2014, 509, 622–626. [Google Scholar] [CrossRef] [Green Version]
- Lesniak, D.R.; Marshall, K.L.; Wellnitz, S.A.; Jenkins, B.A.; Baba, Y.; Rasband, M.N.; Gerling, G.J.; Lumpkin, E.A. Computation identifies structural features that govern neuronal firing properties in slowly adapting touch receptors. Elife 2014, 3, e01488. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Zhu, Y.; La, J.H.; Wills, Z.P.; Gebhart, G.F. Experimental and computational evidence for an essential role of NaV1. 6 in spike initiation at stretch-sensitive colorectal afferent endings. J. Neurophysiol. 2015, 113, 2618–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Siri, S.; Feng, B.; Pierce, D. Computational Modeling of Mouse Colorectum Capturing Longitudinal and Through-thickness Biomechanical Heterogeneity. J. Mech. Behav. Biomed. Mater. 2021, 113, 104127. [Google Scholar] [CrossRef]
- Holzapfel, G.A.; Unterberger, M.J.; Ogden, R.W. An affine continuum mechanical model for cross-linked F-actin networks with compliant linker proteins. J. Mech. Behav. Biomed. Mater. 2014, 38, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Maas, S.A.; Ellis, B.J.; Ateshian, G.A.; Weiss, J.A. FEBio: Finite Elements for Biomechanics. J. Biomech. Eng. 2012, 134, 011005. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.A.; Erdemir, A.; Halloran, J.P.; Weiss, J.A. A general framework for application of prestrain to computational models of biological materials. J. Mech. Behav. Biomed. Mater. 2016, 61, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, N.T.; Hines, M.L. The NEURON Book; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Khaliq, Z.M.; Gouwens, N.W.; Raman, I.M. The contribution of resurgent sodium current to high-frequency firing in Purkinje neurons: An experimental and modeling study. J. Neurosci. 2003, 23, 4899–4912. [Google Scholar] [CrossRef]
- Baker, M.D. Protein kinase C mediates up-regulation of tetrodotoxin-resistant, persistent Na+ current in rat and mouse sensory neurones. J. Physiol. 2005, 567, 851–867. [Google Scholar] [CrossRef]
- Schild, J.; Clark, J.; Hay, M.; Mendelowitz, D.; Andresen, M.; Kunze, D. A-and C-type rat nodose sensory neurons: Model interpretations of dynamic discharge characteristics. J. Neurophysiol. 1994, 71, 2338–2358. [Google Scholar] [CrossRef]
- Jérusalem, A.; García-Grajales, J.A.; Merchán-Pérez, A.; Peña, J.M. A computational model coupling mechanics and electrophysiology in spinal cord injury. Biomech. Model. Mechanobiol. 2014, 13, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Cinelli, I.; Destrade, M.; Duffy, M.; McHugh, P. Electro-mechanical response of a 3D nerve bundle model to mechanical loads leading to axonal injury. Int. J. Numer. Method. Biomed. Eng. 2018, 34, e2942. [Google Scholar] [CrossRef]
- Kwong, M.T.; Bianchi, F.; Malboubi, M.; García-Grajales, J.A.; Homsi, L.; Thompson, M.; Ye, H.; Noels, L.; Jérusalem, A. 3D finite element formulation for mechanical–electrophysiological coupling in axonopathy. Comput. Meth. Appl. Mech. Eng. 2019, 346, 1025–1050. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Model | Parameters | Experimental Data |
|---|---|---|---|
| Higa et al. (2007) | M-R + visco | 4 | in-vivo compression (goat colon) |
| Ciarletta et al. (2009) | N-H + 4 fiber | 8 | uniaxial extension + shear (pig colon) |
| Bellini et al. (2011) | Fung | 4 | biaxial extension (pig small intestine) |
| Sokolis et al. (2011) | Fung | 4 | inflation-extension (mouse intestine) |
| Sokolis + Sassani (2013) | N-H + 2 fiber | 5 | inflation-extension (rat intestine) |
| N-H + 3 fiber | 6 | ||
| N-H + 4 fiber | 8 | ||
| N-H + 5 fiber | 10 | ||
| Carniel et al. (2014) | DEM + 3 fiber | 11 | uniaxial extension (pig colon) |
| Carniel et al. (2015) | DEM + 2 visco | 6 | inflation (pig colon) |
| Sokolis (2017) | Fung | 4 | inflation-extension (rat small intestine) |
| Chuong and Fung | 7 | ||
| Tong and Fung | 7 | ||
| Patel et al. (2018) | N-H + 4 fiber | 9 | inflation-extension (pig colon) |
| Puértolas et al. (2020) | N-H + 2 fiber | 4 | biaxial extension (pig colon) |
| N-H + 2 dis fib | 5 | ||
| N-H + 4 fiber | 8 | ||
| N-H + ODF (Von Mises) | 5 | ||
| N-H + ODF (Bingham) | 5 | ||
| Zhao et al. (2020) | N-H + ODF (diff. tensor) | 4 * | biaxial extension + inflation (mouse colon) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Siri, S.; Feng, B.; Pierce, D.M. The Macro- and Micro-Mechanics of the Colon and Rectum II: Theoretical and Computational Methods. Bioengineering 2020, 7, 152. https://doi.org/10.3390/bioengineering7040152
Zhao Y, Siri S, Feng B, Pierce DM. The Macro- and Micro-Mechanics of the Colon and Rectum II: Theoretical and Computational Methods. Bioengineering. 2020; 7(4):152. https://doi.org/10.3390/bioengineering7040152
Chicago/Turabian StyleZhao, Yunmei, Saeed Siri, Bin Feng, and David M. Pierce. 2020. "The Macro- and Micro-Mechanics of the Colon and Rectum II: Theoretical and Computational Methods" Bioengineering 7, no. 4: 152. https://doi.org/10.3390/bioengineering7040152