The Macro- and Micro-Mechanics of the Colon and Rectum I: Experimental Evidence



Abstract
:1. Introduction
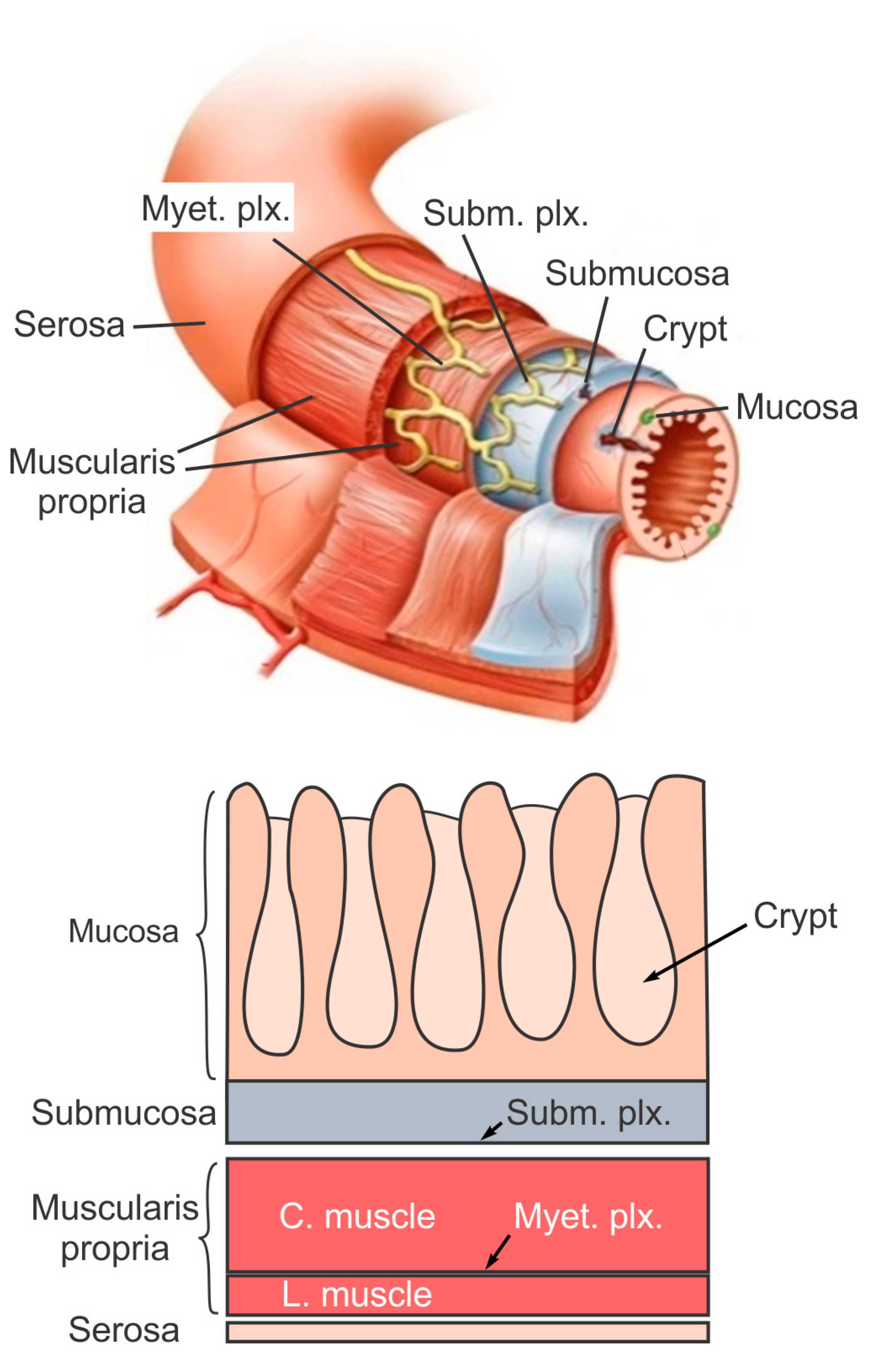
2. The Layered Structure and Function of Large Intestine
3. Macroscale Biomechanics of the Colon and Rectum
3.1. Biomechanical Heterogeneity along the Longitudinal Direction, from Colon to Rectum
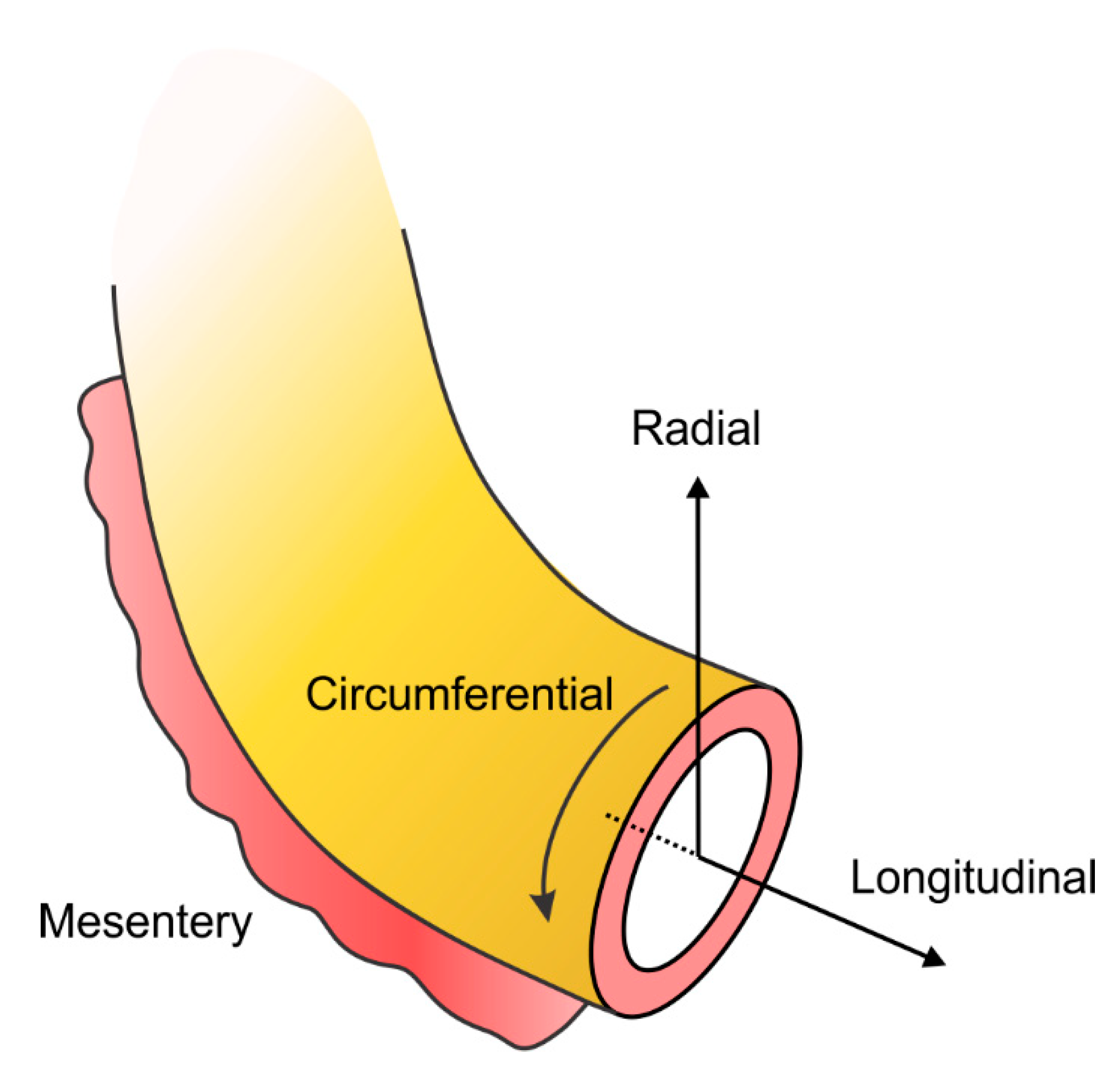
3.2. Biomechanical Heterogeneity along the Radial Direction
3.3. Biomechanical Heterogeneity along the Circumferential Direction
3.4. In-Plane Biomechanical Anisotropy
3.5. Macroscopic Mechanical Tests on Large Intestines with Lower GI Disorders
4. Microscale Experimental Evidence (Large and Small Intestine)
Altered Collagen Morphology and Contents in Lower GI Disorders
5. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Feng, B.; La, J.H.; Schwartz, E.S.; Gebhart, G.F. Irritable bowel syndrome: Methods, mechanisms, and pathophysiology. Neural and neuro-immune mechanisms of visceral hypersensitivity in irritable bowel syndrome. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1085–G1098. [Google Scholar] [CrossRef] [PubMed]
- Lembo, A.; Camilleri, M. Chronic Constipation. N. Engl. J. Med. 2003, 349, 1360–1368. [Google Scholar] [CrossRef]
- Hillemeier, C.; Biancani, P. Mechanical properties of obstructed colon in a Hirschsprung’s model. Gastroenterology 1990, 99, 995–1000. [Google Scholar] [CrossRef]
- Martucciello, G.; Torre, M.; Prato, A.P.; Lerone, M.; Campus, R.; Leggio, S.; Jasonni, V. Associated anomalies in intestinal neuronal dysplasia. J. Pediatr. Surg. 2002, 37, 219–223. [Google Scholar] [CrossRef]
- Pucilowska, J.B.; Williams, K.L.; Lund, P.K. Fibrogenesis IV. Fibrosis and inflammatory bowel disease: Cellular mediators and animal models. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G653–G659. [Google Scholar] [CrossRef]
- Watters, D.; Smith, A. Strength of the colon wall in diverticular disease. Br. J. Surg. 1990, 77, 257–259. [Google Scholar] [CrossRef]
- Almadi, M.A.; Ghosh, S.; Aljebreen, A.M. Differentiating intestinal tuberculosis from Crohn’s disease: A diagnostic challenge. Am. J. Gastroenterol. 2009, 104, 1003–1012. [Google Scholar] [CrossRef]
- Afzalpurkar, R.G.; Schiller, L.R.; Little, K.H.; Santangelo, W.C.; Fordtran, J.S. The Self-Limited Nature of Chronic Idiopathic Diarrhea. N. Engl. J. Med. 1992, 327, 1849–1852. [Google Scholar] [CrossRef]
- Rao, S.S.C. Pathophysiology of adult fecal incontinence. Gastroenterology 2004, 126, S14–S22. [Google Scholar] [CrossRef]
- Feng, B.; Guo, T. Visceral pain from colon and rectum: The mechanotransduction and biomechanics. J. Neural Transm. 2020, 127, 415–429. [Google Scholar] [CrossRef]
- Parks, T. Rectal and colonic studies after resection of the sigmoid for diverticular disease. Gut 1970, 11, 121. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Evans, F.G. Strength of Biological Materials; Evans, F.G., Ed.; Lippincott Williams Wilkins: Philadelphia, PA, USA, 1970. [Google Scholar]
- Arhan, P.; Faverdin, C.; Persoz, B.; Devroede, G.; Dubois, F.; Dornic, C.; Pellerin, D. Relationship between viscoelastic properties of the rectum and anal pressure in man. J. Appl. Physiol. 1976, 41, 677–682. [Google Scholar] [CrossRef]
- Watters, D.; Smith, A.; Eastwood, M.; Anderson, K.; Elton, R. Mechanical properties of the rat colon: The effect of age, sex and different conditions of storage. Q. J. Exp. Physiol. Transl. Integr. 1985, 70, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Watters, D.; Smith, A.; Eastwood, M.; Anderson, K.; Elton, R.; Mugerwa, J. Mechanical properties of the colon: Comparison of the features of the African and European colon in vitro. Gut 1985, 26, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Carniel, E.L.; Gramigna, V.; Fontanella, C.G.; Frigo, A.; Stefanini, C.; Rubini, A.; Natali, A.N. Characterization of the anisotropic mechanical behaviour of colonic tissues: Experimental activity and constitutive formulation. Exp. Physiol. 2014, 99, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Xu, X.; Lin, S.; Cheng, Y.; Tong, J.; Li, Y. Alterations in biomechanical properties and microstructure of colon wall in early-stage experimental colitis. Exp. Ther. Med. 2017, 14, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.; Andreassen, T.; Oxlund, H. Age-related alterations in the strength and collagen content of left colon in rats. Int. J. Colorectal Dis. 1992, 7, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Siri, S.; Maier, F.; Chen, L.; Santos, S.; Pierce, D.M.; Feng, B. Differential biomechanical properties of mouse distal colon and rectum innervated by the splanchnic and pelvic afferents. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G473–G481. [Google Scholar] [CrossRef]
- Siri, S.; Maier, F.; Santos, S.; Pierce, D.M.; Feng, B. Load-bearing function of the colorectal submucosa and its relevance to visceral nociception elicited by mechanical stretch. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G349–G358. [Google Scholar] [CrossRef]
- Puértolas, S.; Peña, E.; Herrera, A.; Ibarz, E.; Gracia, L. A comparative study of hyperelastic constitutive models for colonic tissue fitted to multiaxial experimental testing. J. Mech. Behav. Biomed. Mater. 2020, 102, 103507. [Google Scholar] [CrossRef]
- Qiao, Y.; Pan, E.; Chakravarthula, S.; Han, F.; Liang, J.; Gudlavalleti, S. Measurement of mechanical properties of rectal wall. J. Mater. Sci. Mater. Med. 2005, 16, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Higa, M.; Luo, Y.; Okuyama, T.; Takagi, T.; Shiraishi, Y.; Yambe, T. Passive mechanical properties of large intestine under in vivo and in vitro compression. Med. Eng. Phys. 2007, 29, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.C.; Rubiano, A.; Santisteban, M.M.; Shenoy, V.; Qi, Y.; Pepine, C.J.; Raizada, M.K.; Simmons, C.S. Hypertension-linked mechanical changes of rat gut. Acta Biomater. 2016, 45, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brauchle, E.; Kasper, J.; Daum, R.; Schierbaum, N.; Falch, C.; Kirschniak, A.; Schäffer, T.E.; Schenke-Layland, K. Biomechanical and biomolecular characterization of extracellular matrix structures in human colon carcinomas. Matrix Biol. 2018, 68–69, 180–193. [Google Scholar] [CrossRef]
- Sokolis, D.P.; Orfanidis, I.K.; Peroulis, M. Biomechanical testing and material characterization for the rat large intestine: Regional dependence of material parameters. Physiol. Meas. 2011, 32, 1969. [Google Scholar] [CrossRef]
- Sokolis, D.P.; Sassani, S.G. Microstructure-based constitutive modeling for the large intestine validated by histological observations. J. Mech. Behav. Biomed. Mater. 2013, 21, 149–166. [Google Scholar] [CrossRef]
- Drewes, A.M.; Petersen, P.; Rössel, P.; Gao, C.; Hansen, J.B.; Arendt-Nielsen, L. Sensitivity and distensibility of the rectum and sigmoid colon in patients with irritable bowel syndrome. Scand. J. Gastroenterol. 2001, 36, 827–832. [Google Scholar] [CrossRef]
- Patel, B.; Chen, H.; Ahuja, A.; Krieger, J.F.; Noblet, J.; Chambers, S.; Kassab, G.S. Constitutive modeling of the passive inflation-extension behavior of the swine colon. J. Mech. Behav. Biomed. Mater. 2018, 77, 176–186. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Hubmayr, R.D.; Ferber, I.J.; Zinsmeister, A.R. Viscoelastic properties of the human colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G459–G466. [Google Scholar] [CrossRef] [Green Version]
- Bassotti, G.; Crowell, M.D.; Whitehead, W.E. Contractile activity of the human colon: Lessons from 24 hour studies. Gut 1993, 34, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Egorov, V.I.; Schastlivtsev, I.V.; Prut, E.V.; Baranov, A.O.; Turusov, R.A. Mechanical properties of the human gastrointestinal tract. J. Biomech. 2002, 35, 1417–1425. [Google Scholar] [CrossRef]
- Chen, J.; Ahn, T.; Colón-Bernal, I.D.; Kim, J.; Banaszak Holl, M.M. The Relationship of Collagen Structural and Compositional Heterogeneity to Tissue Mechanical Properties: A Chemical Perspective. ACS Nano 2017, 11, 10665–10671. [Google Scholar] [CrossRef] [PubMed]
- Offner, F.A.; Jao, R.V.; Lewin, K.J.; Havelec, L.; Weinstein, W.M. Collagenous colitis: A study of the distribution of morphological abnormalities and their histological detection. Hum. Pathol. 1999, 30, 451–457. [Google Scholar] [CrossRef]
- Bode, M.K.; Karttunen, T.J.; Mäkelä, J.; Risteli, L.; Risteli, J. Type I and III Collagens in Human Colon Cancer and Diverticulosis. Scand. J. Gastroenterol. 2000, 35, 747–752. [Google Scholar] [CrossRef]
- Krarup, P.-M.; Rehn, M.; Sand-Dejmek, J.; Ehrnström, R.; Ågren, M.S.; Syk, I. Rapid morphological changes and loss of collagen following experimental acute colonic obstruction. Int. J. Colorectal Dis. 2013, 28, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Segnani, C.; Ippolito, C.; Antonioli, L.; Pellegrini, C.; Blandizzi, C.; Dolfi, A.; Bernardini, N. Histochemical detection of collagen fibers by sirius red/fast green is more sensitive than van gieson or sirius red alone in normal and inflamed rat colon. PLoS ONE 2015, 10, e0144630. [Google Scholar] [CrossRef]
- Williams, R.M.; Zipfel, W.R.; Webb, W.W. Interpreting second-harmonic generation images of collagen I fibrils. Biophys. J. 2005, 88, 1377–1386. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Zhu, Y.; La, J.H.; Wills, Z.P.; Gebhart, G.F. Experimental and computational evidence for an essential role of NaV1.6 in spike initiation at stretch-sensitive colorectal afferent endings. J. Neurophysiol. 2015, 113, 2618–2634. [Google Scholar] [CrossRef] [Green Version]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286. [Google Scholar] [CrossRef]
- Steadman, C.J.; Phillips, S.F.; Camilleri, M.; Talley, N.J.; Haddad, A.; Hanson, R. Control of muscle tone in the human colon. Gut 1992, 33, 541–546. [Google Scholar] [CrossRef] [Green Version]
- Spencer, N.J.; Kyloh, M.; Duffield, M. Identification of different types of spinal afferent nerve endings that encode noxious and innocuous stimuli in the large intestine using a novel anterograde tracing technique. PLoS ONE 2014, 9, e112466. [Google Scholar] [CrossRef] [Green Version]
- Dean, B.B.; Aguilar, D.; Barghout, V.; Kahler, K.H.; Frech, F.; Groves, D.; Ofman, J.J. Impairment in work productivity and health-related quality of life in patients with IBS. Am. J. Manag. Care 2005, 11, S17–S26. [Google Scholar] [PubMed]
- Maier, F.; Siri, S.; Santos, S.; Chen, L.; Feng, B.; Pierce, D.M. The heterogeneous morphology of networked collagen in distal colon and rectum of mice quantified via nonlinear microscopy. J. Mech. Behav. Biomed. Mater. 2020, 113, 104116. [Google Scholar] [CrossRef]
- Shamsuddin, A.M.; Phelps, P.C.; Trump, B.F. Human large intestinal epithelium: Light microscopy, histochemistry, and ultrastructure. Hum. Pathol. 1982, 13, 790–803. [Google Scholar] [CrossRef]
- Robert, M.E.; Singh, S.K.; Ikuma, M.; Jain, D.; Ardito, T.; Binder, H.J. Morphology of isolated colonic crypts. Cells Tissues Organs 2001, 168, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P.; Michl, T.; Danzer, M.; Jocic, M.; Schicho, R.; Lippe, I.T. Surveillance of the gastrointestinal mucosa by sensory neurons. J. Physiol. Pharmacol. 2001, 52 Pt 1, 505–521. [Google Scholar]
- Smith, A.; Shepherd, J.; Eastwood, M. Pressure changes after balloon distension of the colon wall in diverticular disease. Gut 1981, 22, 841–844. [Google Scholar] [CrossRef]
- Dall, F.; Jørgensen, C.; Houe, D.; Gregersen, H.; Djurhuus, J. Biomechanical wall properties of the human rectum. A study with impedance planimetry. Gut 1993, 34, 1581–1586. [Google Scholar] [CrossRef] [Green Version]
- Petersen, P.; Gao, C.; Rössel, P.; Qvist, P.; Arendt-Nielsen, L.; Gregersen, H.; Drewes, A.M. Sensory and biomechanical responses to distension of the normal human rectum and sigmoid colon. Digestion 2001, 64, 191–199. [Google Scholar] [CrossRef]
- Drewes, A.M.; Frφkjær, J.B.; Larsen, E.; Reddy, H.; Arendt-Nielsen, L.; Gregersen, H. Pain and mechanical properties of the rectum in patients with active ulcerative colitis. Inflamm. Bowel Dis. 2006, 12, 294–303. [Google Scholar] [CrossRef]
- Glavind, E.; Forman, A.; Madsen, G.; Svane, D.; Andersson, K.; Tottrup, A. Mechanical properties of isolated smooth muscle from human rectum and internal anal sphincter. Am. J. Physiol. Gastrointest. Liver Physiol. 1993, 265, G792–G798. [Google Scholar] [CrossRef] [PubMed]
- Massalou, D.; Masson, C.; Foti, P.; Afquir, S.; Baqué, P.; Berdah, S.-V.; Bège, T. Dynamic biomechanical characterization of colon tissue according to anatomical factors. J. Biomech. 2016, 49, 3861–3867. [Google Scholar] [CrossRef] [PubMed]
- Massalou, D.; Masson, C.; Afquir, S.; Baqué, P.; Arnoux, P.-J.; Bège, T. Mechanical effects of load speed on the human colon. J. Biomech. 2019, 91, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Massalou, D.; Masson, C.; Afquir, S.; Baqué, P.; Arnoux, P.-J.; Bège, T. Influence of gender, age, shelf-life, and conservation method on the biomechanical behavior of colon tissue under dynamic solicitation. Clin. Biomech. 2019, 65, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.K.; Hardy, W.N. Material properties of the post-mortem colon in high-rate equibiaxial elongation. Biomed. Sci. Instrum. 2012, 48, 171–178. [Google Scholar] [PubMed]
- Carniel, E.L.; Mencattelli, M.; Bonsignori, G.; Fontanella, C.G.; Frigo, A.; Rubini, A.; Stefanini, C.; Natali, A.N. Analysis of the structural behaviour of colonic segments by inflation tests: Experimental activity and physio-mechanical model. Proc. Inst. Mech. Eng. Part H J. Eng. Med. 2015, 229, 794–803. [Google Scholar] [CrossRef]
- Ciarletta, P.; Dario, P.; Tendick, F.; Micera, S. Hyperelastic model of anisotropic fiber reinforcements within intestinal walls for applications in medical robotics. Int. J. Robot. Res. 2009, 28, 1279–1288. [Google Scholar] [CrossRef]
- Christensen, M.B.; Oberg, K.; Wolchok, J.C. Tensile properties of the rectal and sigmoid colon: A comparative analysis of human and porcine tissue. Springerplus 2015, 4, 142. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Gregersen, H. Biomechanical and morphological properties in rat large intestine. J. Biomech. 2000, 33, 1089–1097. [Google Scholar] [CrossRef]
- Feng, B.; Brumovsky, P.R.; Gebhart, G.F. Differential roles of stretch-sensitive pelvic nerve afferents innervating mouse distal colon and rectum. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G402–G409. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Gebhart, G.F. Characterization of silent afferents in the pelvic and splanchnic innervations of the mouse colorectum. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G170–G180. [Google Scholar] [CrossRef]
- Matrana, M.R.; Margolin, D.A. Epidemiology and pathophysiology of diverticular disease. Clin. Colon Rectal Surg. 2009, 22, 141–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, L. Postmortem survey of diverticular disease of the colon. I. Diverticulosis and diverticulitis. Gut 1969, 10, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reihsner, R.; Menzel, E. Two-dimensional stress-relaxation behavior of human skin as influenced by non-enzymatic glycation and the inhibitory agent aminoguanidine. J. Biomech. 1998, 31, 985–993. [Google Scholar] [CrossRef]
- Ker, R.F. The design of soft collagenous load-bearing tissues. J. Exp. Biol. 1999, 202, 3315–3324. [Google Scholar]
- Hariton, I.; Debotton, G.; Gasser, T.C.; Holzapfel, G.A. Stress-driven collagen fiber remodeling in arterial walls. Biomech. Model. Mechanobiol. 2007, 6, 163–175. [Google Scholar] [CrossRef]
- Hulmes, D.J. Building collagen molecules, fibrils, and suprafibrillar structures. J. Struct. Biol. 2002, 137, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Orberg, J.; Klein, L.; Hiltner, A. Scanning electron microscopy of collagen fibers in intestine. Connect. Tissue Res. 1982, 9, 187–193. [Google Scholar] [CrossRef]
- Orberg, J.; Baer, E.; Hiltner, A. Organization of collagen fibers in the intestine. Connect. Tissue Res. 1983, 11, 285–297. [Google Scholar] [CrossRef]
- Gabella, G. The collagen fibrils in the collapsed and the chronically stretched intestinal wall. J. Ultrastruct. Res. 1983, 85, 127–138. [Google Scholar] [CrossRef]
- Storkholm, J.; Villadsen, G.; Jensen, S.; Gregersen, H. Mechanical properties and collagen content differ between isolated guinea pig duodenum, jejunum, and distal ileum. Dig. Dis. Sci. 1998, 43, 2034–2041. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.-J.; Qiao, A.-K.; Yu, J.-D.; Zhao, J.-B.; Liao, D.-H.; Xu, X.-H.; Hans, G. Collagen fiber angle in the submucosa of small intestine and its application in gastroenterology. World J. Gastroenterol. WJG 2003, 9, 804. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zeng, Y.; Zhao, J.; Liao, D.; Gregersen, H. Quantitative analysis of collagen fiber angle in the submucosa of small intestine. Comput. Biol. Med. 2004, 34, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhong, J.; Liu, Y.; Yu, H.; Zhuo, S.; Chen, J. Two-photon fluorescence and second-harmonic generation imaging of collagen in human tissue based on multiphoton microscopy. Scanning 2011, 33, 53–56. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, S.; Zhu, X.; Chen, J.; Xie, S.; Wu, G. Quantitative biomarkers of colonic dysplasia based on intrinsic second-harmonic generation signal. J. Biomed. Opt. 2011, 16, 120501. [Google Scholar] [CrossRef]
- Zhuo, S.; Yan, J.; Chen, G.; Shi, H.; Zhu, X.; Lu, J.; Chen, J.; Xie, S. Label-free imaging of basement membranes differentiates normal, precancerous, and cancerous colonic tissues by second-harmonic generation microscopy. PLoS ONE 2012, 7, e38655. [Google Scholar] [CrossRef]
- Liu, N.; Chen, J.; Xu, R.; Jiang, S.; Xu, J.; Chen, R. Label-free imaging characteristics of colonic mucinous adenocarcinoma using multiphoton microscopy. Scanning 2013, 35, 277–282. [Google Scholar] [CrossRef]
- Schürmann, S.; Foersch, S.; Atreya, R.; Neumann, H.; Friedrich, O.; Neurath, M.F.; Waldner, M.J. Label-free imaging of inflammatory bowel disease using multiphoton microscopy. Gastroenterology 2013, 145, 514–516. [Google Scholar] [CrossRef]
- Bianchi, M.; Adur, J.; Ruff, S.Y.; Izaguirre, M.F.; Carvalho, H.F.; Cesar, C.L.; Casco, V.H. Mouse colorectal cancer an early detection approach using nonlinear microscopy. Bio-Med Mater. Eng. 2014, 24, 3419–3426. [Google Scholar] [CrossRef] [Green Version]
- Birk, J.W.; Tadros, M.; Moezardalan, K.; Nadyarnykh, O.; Forouhar, F.; Anderson, J.; Campagnola, P. Second harmonic generation imaging distinguishes both high-grade dysplasia and cancer from normal colonic mucosa. Dig. Dis. Sci. 2014, 59, 1529–1534. [Google Scholar] [CrossRef]
- Mao, H.; Su, P.; Qiu, W.; Huang, L.; Yu, H.; Wang, Y. The use of Masson’s trichrome staining, second harmonic imaging and two-photon excited fluorescence of collagen in distinguishing intestinal tuberculosis from Crohn’s disease. Colorectal Dis. 2016, 18, 1172–1178. [Google Scholar] [CrossRef]
- He, K.; Zhao, L.; Chen, Y.; Huang, X.; Ding, Y.; Hua, H.; Liu, L.; Wang, X.; Wang, M.; Zhang, Y. Label-free multiphoton microscopic imaging as a novel real-time approach for discriminating colorectal lesions: A preliminary study. J. Gastroenterol. Hepatol. 2019, 34, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Sarri, B.; Canonge, R.; Audier, X.; Simon, E.; Wojak, J.; Caillol, F.; Cador, C.; Marguet, D.; Poizat, F.; Giovannini, M. Fast stimulated raman and second harmonic generation imaging for intraoperative gastro-intestinal cancer detection. Sci. Rep. 2019, 9, 10052. [Google Scholar] [CrossRef] [PubMed]
- Despotović, S.Z.; Milićević, Đ.N.; Krmpot, A.J.; Pavlović, A.M.; Živanović, V.D.; Krivokapić, Z.; Pavlović, V.B.; Lević, S.; Nikolić, G.; Rabasović, M.D. Altered organization of collagen fibers in the uninvolved human colon mucosa 10 cm and 20 cm away from the malignant tumor. Sci. Rep. 2020, 10, 6359. [Google Scholar] [CrossRef] [PubMed]
- Feng., B.; Maier., F.; Siri., S.; Pierce., D.M. Quantifying the collagen-network morphology in mouse distal colon and rectum via nonlinear microscopy. In Proceedings of the Biomedical Engineering Society 2019 Annual Fall Meeting, Philadelphia, PA, USA, 12–16 October 2019; pp. 16–19. [Google Scholar]
- Zonios, G.I.; Cothren, R.M.; Arendt, J.T.; Wu, J.; Van Dam, J.; Crawford, J.M.; Manoharan, R.; Feld, M.S. Morphological model of human colon tissue fluorescence. IEEE Trans. Biomed. Eng. 1996, 43, 113–122. [Google Scholar] [CrossRef]
- Abraham, G.A.; Murray, J.; Billiar, K.; Sullivan, S.J. Evaluation of the porcine intestinal collagen layer as a biomaterial. J. Biomed. Mater. Res. 2000, 51, 442–452. [Google Scholar] [CrossRef]
- Sacks, M.S.; Gloeckner, D.C. Quantification of the fiber architecture and biaxial mechanical behavior of porcine intestinal submucosa. J. Biomed. Mater. Res. 1999, 46, 1–10. [Google Scholar] [CrossRef]
- Donnellan, W.L. Early histological changes in ulcerative colitis: A light and electron microscopic study. Gastroenterology 1966, 50, 519–540. [Google Scholar] [CrossRef]
- Thomson, H.; Busuttil, A.; Eastwood, M.; Smith, A.; Elton, R. Submucosal collagen changes in the normal colon and in diverticular disease. Int. J. Colorectal Dis. 1987, 2, 208–213. [Google Scholar] [CrossRef]
- Xu, R.a.; Zhu, X.; He, N.; Zhuo, S.; Xu, J.; Jiang, S.; Li, H.; Chen, J. Multiphoton microscopic imaging of mouse intestinal mucosa based on two-photon excited fluorescence and second harmonic generation. J. Innov. Opt. Health Sci. 2013, 6, 1350004. [Google Scholar] [CrossRef] [Green Version]
- Prieto, S.P.; Reed, C.L.; James, H.M.; Quinn, K.P.; Muldoon, T.J. Differences in colonic crypt morphology of spontaneous and colitis-associated murine models via second harmonic generation imaging to quantify colon cancer development. BMC Cancer 2019, 19, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnellan, W.L.; Beal, J.M. Early pathologic changes in ulcerative colitis: Their relation to surgical complications. Am. J. Surg. 1966, 111, 107–113. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Specimen | Condition | Test Methods | References |
|---|---|---|---|
| Mechanical tests on whole intestinal wall | |||
| Human | In vivo | Pressure-volume | Parks, 1970 [11]; Smith et al., 1981 [48]; Bharucha et al., 2001 [30] |
| Pressure-CSA (Cross-section area) | Arhan et al., 1976 [13]; Dall et al., 1993 [49]; Drewes et al., 2001 [28]; Petersen et al., 2001 [50]; Drewes et al., 2006 [51] | ||
| Human | In vitro | Uniaxial stretch | Watters et al., 1985b [15]; Glavind et al., 1993 [52]; Massalou et al., 2016 [53]; Massalou et al., 2019 [54]; Massalou et al., 2019 [55] |
| Biaxial stretch | Howes and Hardy, 2012 [56] | ||
| Porcine | In vitro | Inflation | Carniel et al., 2015 [57]; Patel et al., 2018 [29,57] |
| Compression and shear | Qiao et al., 2005 [22] | ||
| Uniaxial stretch | Qiao et al., 2005 [22]; Ciarletta et al., 2009 [58]; Carniel et al., 2014 [16]; Christensen et al., 2015 [59] | ||
| Biaxial extension | Puértolas et al., 2020 [21] | ||
| Goats | In vitro | Compression | Higa et al., 2007 [23] |
| Rats | In vitro | Pressure-diameter | Gao and Gregersen, 2000 [60]; Sokolis et al., 2011 [26]; Sokolis and Sassani, 2013 [27] |
| Uniaxial stretch | Watters et al., 1985 [14] | ||
| Indentation | Stewart et al., 2016 [24] | ||
| Mice | In vitro | Biaxial stretch | Siri et al., 2019 [19] |
| Mechanical tests on separated intestinal layers | |||
| Mice | In vitro | Biaxial stretch | Siri et al., 2019 [20] |
| Human | In vitro | Uniaxial stretch | Egorov et al., 2002 [32] |
| Specimens | Tissue | Layers | References | ||
|---|---|---|---|---|---|
| Chromatic and fluorescent staining | |||||
| Human | Large intestine | Mucosa | Zonios et al., 1996 [87] | ||
| Rats | Large intestine | Mucosa | Sokolis and Sassani, 2013 [27] | ||
| Submucosa | Sokolis and Sassani, 2013 [27] | ||||
| Muscular layers | Sokolis and Sassani, 2013 [27] | ||||
| Porcine | Small intestine | Submucosa | Abraham et al., 2000 [88] | ||
| Small-angle light scattering | |||||
| Porcine | Small intestine | Submucosa | Sacks and Gloeckner, 1999 [89] | ||
| Polarized light microscopy | |||||
| Rats | Small intestine | Submucosa | Orberg et al., 1983 [70]; Zeng et al., 2003 [73]; Yu et al., 2004 [74] | ||
| Electron microscopy | |||||
| Human | Large intestine | Mucosa | Donnellan et al., 1966 [90]; Shamsuddin et al., 1982 [45] | ||
| Submucosa | Thomson et al., 1987 [91] | ||||
| Rats | Small intestine | Submucosa | Orberg et al., 1982 [69]; Orberg et al., 1983 [70]; Gabella, 1983 [71] | ||
| Porcine | Small intestine | Submucosa | Gabella, 1983 [71] | ||
| Two-photon excited fluorescence and second harmonic generation microscopy | |||||
| Human | Large intestine | Mucosa | Zhuo et al., 2011 [76]; Zhuo et al., 2012 [77]; Liu et al., 2013 [78]; Schürmann et al., 2013 [79]; Bianchi et al., 2014 [80]; Birk et al., 2014 [81]; Mao et al., 2016 [82]; He et al., 2019 [83]; Sarri et al., 2019 [84]; Despotović et al., 2020 [85] | ||
| Submucosa | Jiang et al., 2011 [75]; Bianchi et al., 2014 [80] | ||||
| Mice | Large intestine | Mucosa | Xu et al., 2013 [92]; Prieto et al., 2019 [93]; Maier et al., 2020 [44] | ||
| Submucosa | Maier et al., 2020 [44] | ||||
| Muscular layers | Maier et al., 2020 [44] | ||||
| Serosa | Maier et al., 2020 [44] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siri, S.; Zhao, Y.; Maier, F.; Pierce, D.M.; Feng, B. The Macro- and Micro-Mechanics of the Colon and Rectum I: Experimental Evidence. Bioengineering 2020, 7, 130. https://doi.org/10.3390/bioengineering7040130
Siri S, Zhao Y, Maier F, Pierce DM, Feng B. The Macro- and Micro-Mechanics of the Colon and Rectum I: Experimental Evidence. Bioengineering. 2020; 7(4):130. https://doi.org/10.3390/bioengineering7040130
Chicago/Turabian StyleSiri, Saeed, Yunmei Zhao, Franz Maier, David M. Pierce, and Bin Feng. 2020. "The Macro- and Micro-Mechanics of the Colon and Rectum I: Experimental Evidence" Bioengineering 7, no. 4: 130. https://doi.org/10.3390/bioengineering7040130