
Temperature-Controlled Expression of a Recombinant Human-like Collagen I Peptide in Escherichia coli

Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Recombinant pBV220-rhLCOL-I Expression Vector
2.2. Analysis of rhLCOL-I Expression
2.3. Solubility Analysis
2.4. Plasmid Stability
2.5. Fermentation of rhLCOL-I
2.6. Purification and Identification of rhLCOL-I
2.7. Bioactivity Assay of rhLCol-I
2.8. Minimally Invasive Skin Recovery of rhLCol-I
2.9. Statistical Analysis
3. Results
3.1. Plasmid Construction and Expression of rhLCOL-I
3.2. Plasmid and Thermal Stability of rhLCOL-I
3.3. Fermentation, Purification, and Identification of rhLCOL-I
3.4. rhLCOL-I Promoted HaCaT Cell Migration and Adhesion
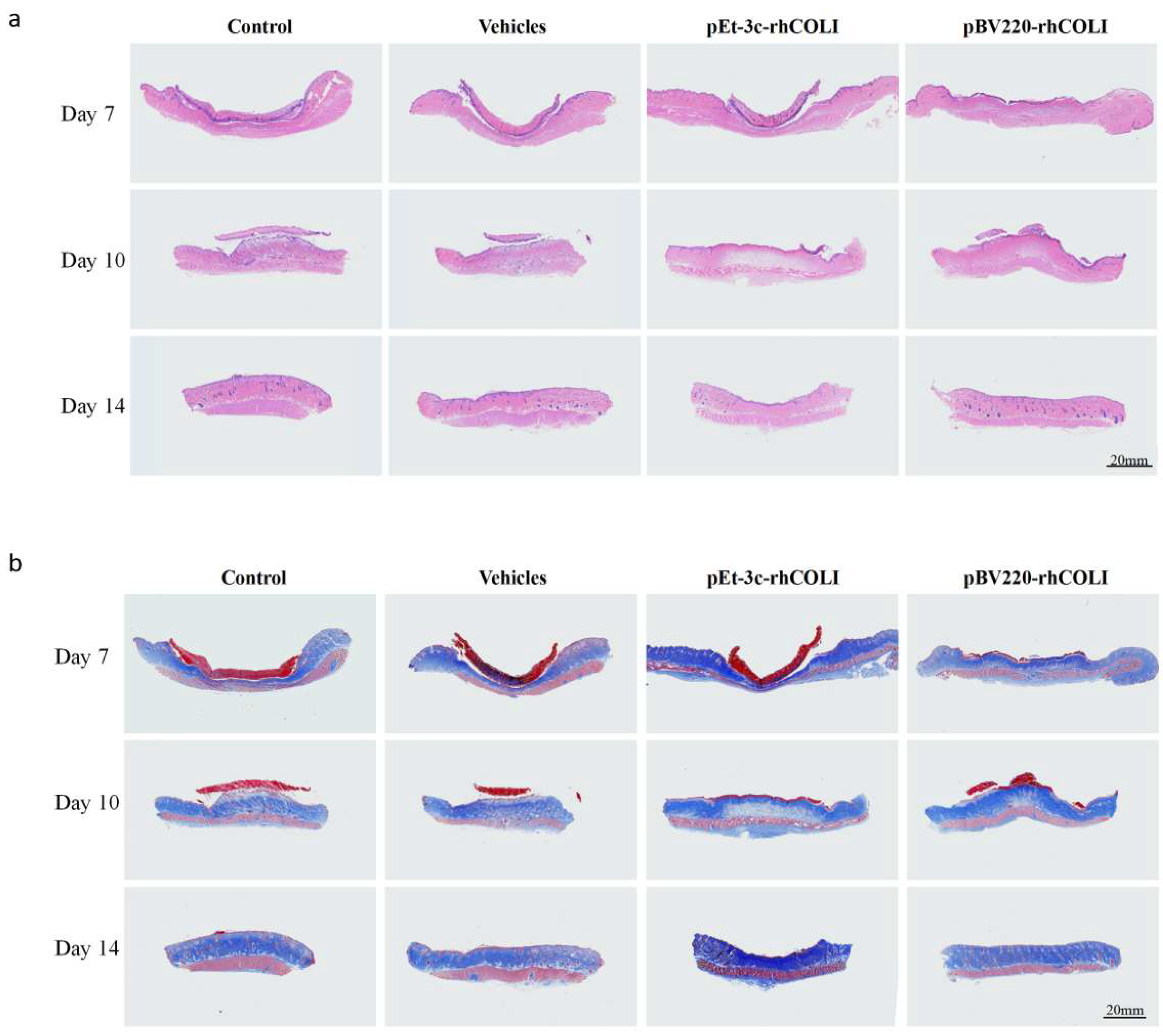
3.5. rhLCOL-I Improved the Wound-Healing Process
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avila Rodríguez, M.I.; Rodríguez Barroso, L.G.; Sánchez, M.L. Collagen: A review on its sources and potential cosmetic applications. J. Cosmet. Dermatol. 2018, 17, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Owczarzy, A.; Kurasiński, R.; Kulig, K.; Rogóż, W.; Szkudlarek, A.; Maciążek-Jurczyk, M. Collagen—Structure, properties and application. Eng. Biomater. 2020, 23, 17–23. [Google Scholar]
- Meyer, M. Processing of collagen based biomaterials and the resulting materials properties. Biomed. Eng. Online 2019, 18, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzi, A.; Gallo, N.; Bettini, S.; Sibillano, T.; Altamura, D.; Madaghiele, M.; De Caro, L.; Valli, L.; Salvatore, L.; Sannino, A.; et al. Sub- and Supramolecular X-Ray Characterization of Engineered Tissues from Equine Tendon, Bovine Dermis, and Fish Skin Type-I Collagen. Macromol. Biosci. 2020, 20, e2000017. [Google Scholar] [CrossRef]
- Kuivaniemi, H.; Tromp, G. Type III collagen (COL3A1): Gene and protein structure, tissue distribution, and associated diseases. Gene 2019, 707, 151–171. [Google Scholar] [CrossRef]
- Senadheera, T.R.; Dave, D.; Shahidi, F. Sea Cucumber Derived Type I Collagen: A Comprehensive Review. Mar. Drugs 2020, 18, 471. [Google Scholar] [CrossRef]
- Zhang, Y.; Stefanovic, B. LARP6 Meets Collagen mRNA: Specific Regulation of Type I Collagen Expression. Int. J. Mol. Sci. 2016, 17, 419. [Google Scholar] [CrossRef] [Green Version]
- Gistelinck, C.; Gioia, R.; Gagliardi, A.; Tonelli, F.; Marchese, L.; Bianchi, L.; Landi, C.; Bini, L.; Huysseune, A.; Witten, P.E.; et al. Zebrafish Collagen Type I: Molecular and Biochemical Characterization of the Major Structural Protein in Bone and Skin. Sci. Rep. 2016, 6, 21540. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Nikoo, M.; Boran, G.; Zhou, P.; Regenstein, J.M. Collagen and Gelatin. Annu. Rev. Food Sci. Technol. 2015, 6, 527–557. [Google Scholar] [CrossRef]
- Kou, Z.; Li, B.; Aierken, A.; Tan, N.; Li, C.; Han, M.; Jing, Y.; Li, N.; Zhang, S.; Peng, S.; et al. Mesenchymal Stem Cells Pretreated with Collagen Promote Skin Wound-Healing. Int. J. Mol. Sci. 2023, 24, 8688. [Google Scholar] [CrossRef]
- Hwang, S.J.; Ha, G.-H.; Seo, W.-Y.; Kim, C.K.; Kim, K.; Lee, S.B. Human collagen alpha-2 type I stimulates collagen synthesis, wound healing, and elastin production in normal human dermal fibroblasts (HDFs). BMB Rep. 2020, 53, 539–544. [Google Scholar]
- Zhao, Y.; Bai, L.; Zhang, Y.; Yao, R.; Sun, Y.; Hang, R.; Chen, X.; Wang, H.; Yao, X.; Xiao, Y.; et al. Type I collagen decorated nanoporous network on titanium implant surface promotes osseointegration through mediating immunomodulation, angiogenesis, and osteogenesis. Biomaterials 2022, 288, 121684. [Google Scholar] [CrossRef]
- Zhou, N.; Liu, Y.-D.; Zhang, Y.; Gu, T.-W.; Peng, L.-H. Pharmacological Functions, Synthesis, and Delivery Progress for Collagen as Biodrug and Biomaterial. Pharmaceutics 2023, 15, 1443. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Fan, D.; Shang, L. Exploring the potential of the recombinant human collagens for biomedical and clinical applications: A short review. Biomed. Mater. 2020, 16, 012001. [Google Scholar] [CrossRef]
- Rutschmann, C.; Baumann, S.; Cabalzar, J.; Luther, K.B.; Hennet, T. Recombinant expression of hydroxylated human collagen in Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 4445–4455. [Google Scholar] [CrossRef] [Green Version]
- Nokelainen, M.; Tu, H.; Vuorela, A.; Notbohm, H.; Kivirikko, K.I.; Myllyharju, J. High-level production of human type I collagen in the yeastPichia pastoris. Yeast 2001, 18, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Shoseyov, O.; Posen, Y.; Grynspan, F. Human collagen produced in plants: More than just another molecule. Bioengineered 2014, 5, 49–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fertala, A. Three Decades of Research on Recombinant Collagens: Reinventing the Wheel or Developing New Biomedical Products? Bioengineering 2020, 7, 155. [Google Scholar] [CrossRef] [PubMed]
- Browne, S.; Zeugolis, D.I.; Pandit, A.; Hackethal, J.; Mühleder, S.; Hofer, A.; Schneider, K.H.; Prüller, J.; Hennerbichler, S.; Redl, H.; et al. Collagen: Finding a Solution for the Source. Tissue Eng. Part A 2013, 19, 1491–1494. [Google Scholar] [CrossRef] [Green Version]
- Sheng, J.; An, K.; Chunsheng, D.; Li, W.; Bao, X.; Qiu, D. Cloning a Cuticle-Degrading Serine Protease Gene with Bio-logic Control Function from Beauveria Brongniartii and Its Expression in Escherichia Coli. Curr. Microbiol. 2006, 53, 124–128. [Google Scholar] [CrossRef]
- Guo–xi, Z.; Kang, Z.; Yang, J.; Jun–rong, W.; Hong–liang, Z. Construction of Prokaryotic Expression Vector for Pbv220/NT4–ADNF–9. J. Otol. 2008, 3, 103–107. [Google Scholar] [CrossRef]
- Jin, J.; Tan, T.; Wang, H.; Su, G. The Expression of Spinach Glycolate Oxidase (GO) in E. Coli and the Application of GO in the Production of Glyoxylic Acid. Mol. Biotechnol. 2003, 25, 207–214. [Google Scholar] [CrossRef]
- He, K.; Wei, F.; Zhang, X.; Li, J.; Zhang, D.; Wei, D. Production and characterization of a single-chain Fv antibody–alkaline phosphatase fusion protein specific for ampicillin. Food Agric. Immunol. 2018, 30, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Salvatore, L.; Gallo, N.; Natali, M.L.; Terzi, A.; Sannino, A.; Madaghiele, M. Mimicking the Hierarchical Organization of Natural Collagen: Toward the Development of Ideal Scaffolding Material for Tissue Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 644595. [Google Scholar] [CrossRef] [PubMed]
- Mccarty, N.S.; Ledesma-Amaro, R. Synthetic biology tools to engineer microbial communities for biotechnology. Trends Biotechnol. 2019, 37, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Ledesma-Fernandez, A.; Velasco-Lozano, S.; Santiago-Arcos, J.; López-Gallego, F.; Cortajarena, A.L. Engineered repeat proteins as scaffolds to assemble multi-enzyme systems for efficient cell-free biosynthesis. Nat. Commun. 2023, 14, 2587. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, B.; Ramshaw, J.A. Bioengineered Collagens. Subcell Biochem. 2017, 82, 601–629. [Google Scholar] [PubMed]
- Pakkanen, O.; Pirskanen, A.; Myllyharju, J. Selective expression of nonsecreted triple-helical and secreted single-chain recombinant collagen fragments in the yeast Pichia pastoris. J. Biotechnol. 2006, 123, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Ma, X.; Gao, Y.; Fan, D.; Zhu, C.; Mi, Y.; Xue, W. Hydroxylation of Human Type III Collagen Alpha Chain by Recombinant Coexpression with a Viral Prolyl 4-Hydroxylase in Escherichia coli. Protein J. 2017, 36, 322–331. [Google Scholar] [CrossRef]
- Li, L.; Fan, D.; Ma, X.; Deng, J.; He, J. High-level secretory expression and purification of unhydroxylated human collagen α1(III) chain in Pichia pastorisGS115. Biotechnol. Appl. Biochem. 2014, 62, 467–475. [Google Scholar] [CrossRef]
- Tang, Y.; Yang, X.; Hang, B.; Li, J.; Huang, L.; Huang, F.; Xu, Z. Efficient Production of Hydroxylated Human-Like Collagen Via the Co-Expression of Three Key Genes in Escherichia coli Origami (DE3). Appl. Biochem. Biotechnol. 2015, 178, 1458–1470. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.K.; Taga, Y.; Takagi, Y.K.; Masuda, R.; Hattori, S.; Koide, T. The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature. Int. J. Mol. Sci. 2022, 23, 2040. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′–3′) |
|---|---|
| pBV220-SELF-F | CTAGGATCCGTCGACCTGCAGCCAAGCTTC |
| pBV220-SELF-R | GATGATGCATATGGAATTCCTCCTTA |
| rhLCOL-I-SELF-F | GGAGGAATTCCATATATGCATCATCATCATCATCATA CTAGTGGCGAACGTG |
| rhLCOL-I-SELF-R | TGGCTGCAGGTCGACGGATCCTTAATGATGATGATGATGA TGACTA |
| pBV220-TEST-F | CTGAGCACATCAGCAGGAC |
| pBV220-TEST-R | ACAGAAGCTTGGCTGCAGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, W.; Wu, Q.; Kuang, Z.; Cong, J.; Zhang, Q.; Huang, Y.; Su, Z.; Xiang, Q. Temperature-Controlled Expression of a Recombinant Human-like Collagen I Peptide in Escherichia coli. Bioengineering 2023, 10, 926. https://doi.org/10.3390/bioengineering10080926
Xie W, Wu Q, Kuang Z, Cong J, Zhang Q, Huang Y, Su Z, Xiang Q. Temperature-Controlled Expression of a Recombinant Human-like Collagen I Peptide in Escherichia coli. Bioengineering. 2023; 10(8):926. https://doi.org/10.3390/bioengineering10080926
Chicago/Turabian StyleXie, Wenjie, Qiqi Wu, Zhanpeng Kuang, Jianhang Cong, Qirong Zhang, Yadong Huang, Zhijian Su, and Qi Xiang. 2023. "Temperature-Controlled Expression of a Recombinant Human-like Collagen I Peptide in Escherichia coli" Bioengineering 10, no. 8: 926. https://doi.org/10.3390/bioengineering10080926