
Ultrastructure of a Mechanoreceptor of the Trichoid Sensilla of the Insect Nabis rugosus: Stimulus-Transmitting and Bio-Sensory Architecture

Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Collection
2.1.1. Samples under Scanning Electron Microscope
2.1.2. Samples under Transmission Electron Microscope
2.1.3. Image Processing—AMIRA
2.2. Theoretical Model of the Hair Mechanoreceptor Trichoid Sensilla
2.2.1. Mathematical Model
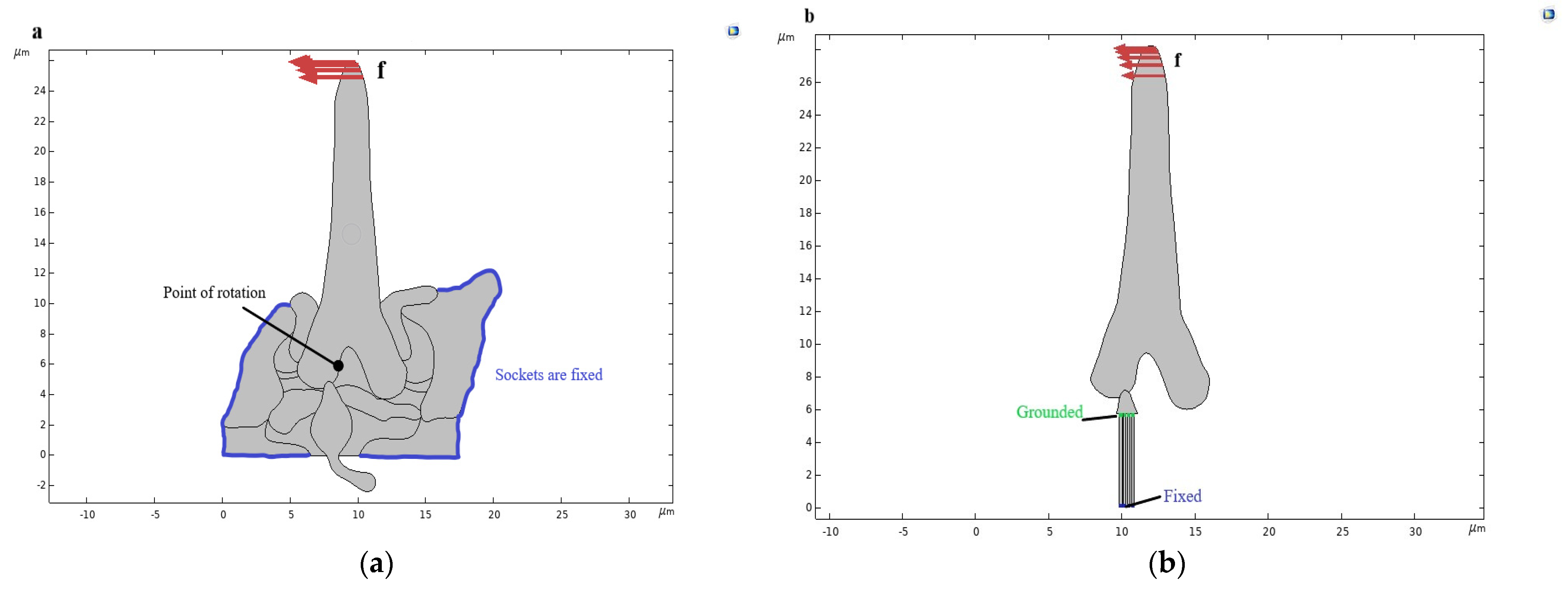
2.2.2. Geometrical Model and Numerical Analysis
2.3. Trichoid Sensilla Mechanical Properties
3. Results
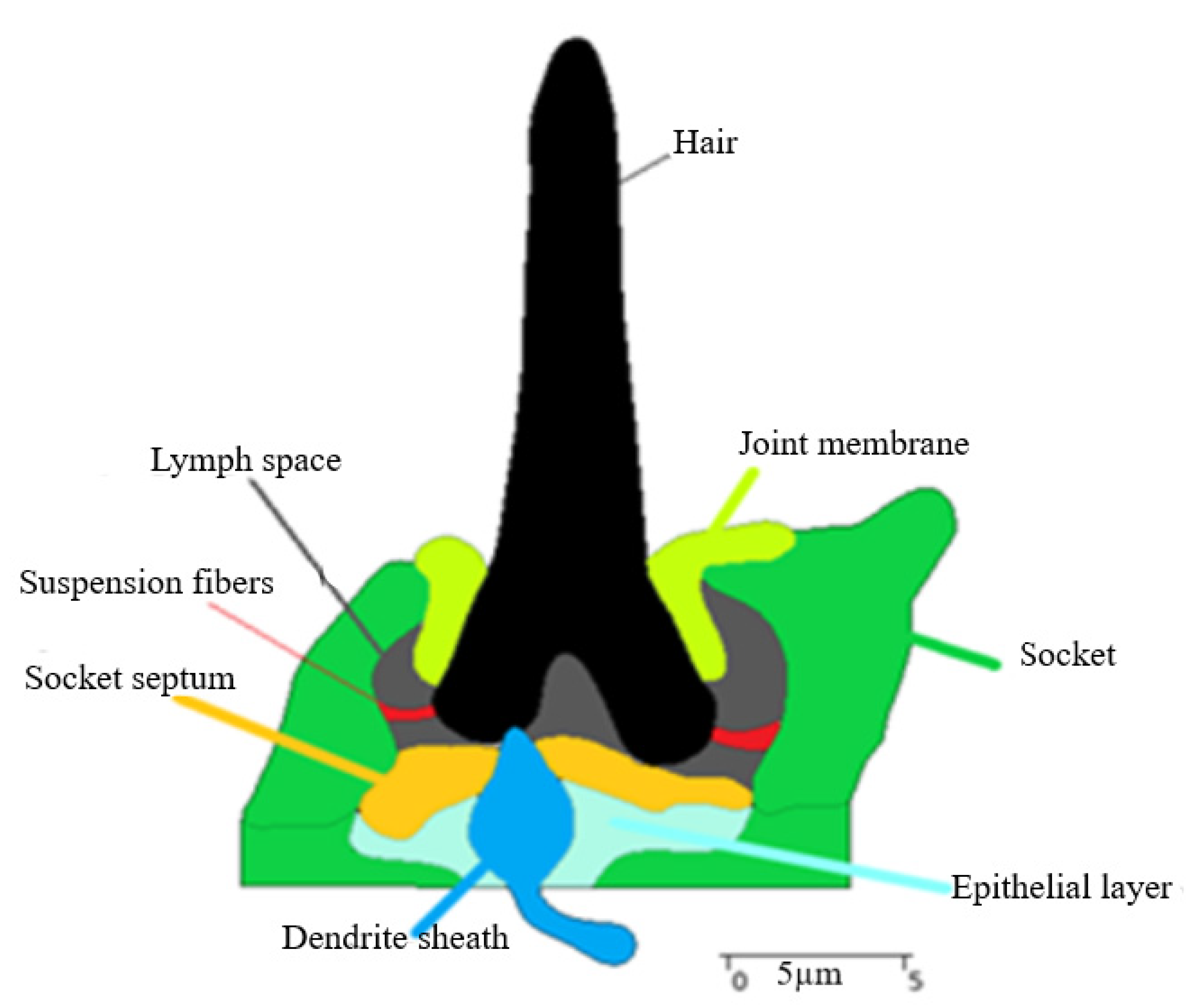
3.1. Trichoid Sensilla Ultrastructure
3.2. Computational Scheme of the Trichoid Sensilla—External Excitation and Boundary Conditions
3.3. Bio-Micro-Electro-Mechanical System of the Hair-Type Mechanoreceptor Trichoid Sensilla
4. Discussion
4.1. Trichoid Mechanosensilla Morphological Parameters
4.2. Hair Deflection and Mechanism of Stimulation Transmission
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kerzhner, I.M. Family Nabidae, A. Costa—Damsel bugs, 1853. In Catalogue of the Heteroptera of the Palaearctic Region, 2nd ed.; Aukema, B., Rieger, C., Eds.; The Netherlands Entomological Society: Wageningen, The Netherlands, 2004; pp. 84–107. [Google Scholar]
- Ghahari, H.; Linnavuori, R.E.; Moulet, P.; Ostovan, H. An annotated catalogue of the Iranian Nabidae (Hemiptera: Heteroptera). Acta Entomol. Musei Natl. Pragae 2010, 50, 33–44. [Google Scholar] [CrossRef]
- Kerzhner, I.M. Poluzhestkokrylye semeystva Nabidae [Heteroptera of the family Zoo morphology]. In Fauna of the USSR. Rhynchota, 2nd ed.; Theodor, O., Ed.; Nauka: Leningrad, Russia, 1981; pp. 1–326. [Google Scholar]
- Chakilam, S.; Brożek, J.; Chajec, Ł.; Poprawa, I.; Gaidys, R. Ultra-Morphology and Mechanical Function of the Trichoideum Sensillum in Nabis rugosus (Linnaeus, 1758) (Insecta: Heteroptera: Cimicomorpha). Insects 2022, 13, 799. [Google Scholar] [CrossRef] [PubMed]
- Taszakowski, A.; Nowińska, A.; Brożek, J. Morphological study of the labial sensilla in Nabidae (Hemiptera: Heteroptera: Cimicomorpha). Zoomorphology 2019, 138, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Chapman, R.F. The Insects: Structure and Function, 5th ed.; Simpson, S.J., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 738–768. [Google Scholar]
- Tautz, J. Reception of particle oscillation in a medium—An unorthodox sensory capacity. Naturwissenschaften 1979, 66, 452–461. [Google Scholar] [CrossRef]
- Fletcher, N.H. Acoustical response of hair receptors in insects. J. Comp. Physiol. 1978, 127, 185–189. [Google Scholar] [CrossRef]
- Thurm, U.; Erler, G.; Gödde, J.; Kastrup, H.; Keil, T.; Volker, W.; Vohwinkel, B. Cilia specialized for mechanoreception. J. Submicrosc. Cytotol. 1983, 15, 151–155. [Google Scholar]
- Barth, F.G. Dynamics of arthropod filiform hairs. V. The response of spider trichobothria to natural stimuli. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 183–192. [Google Scholar] [CrossRef] [Green Version]
- Dechant, H.E. Mechanical Properties and Finite Element Simulation of Spider Tactile Hairs. Ph.D. Thesis, University of Technology, Vienna, Austria, 2001. [Google Scholar]
- Dechan, H.E.; Rammerstorfer, F.G.; Barth, F.G. Arthropod touch reception: Stimulus transformation and finite element model of spider tactile hairs. J. Comp. Physiol. A 2001, 187, b313–b322. [Google Scholar] [CrossRef]
- Albert, J.T.; Friedrich, O.C.; Dechant, H.E.; Barth, F.G. Arthropod touch reception: Spider hair sensilla as rapid touch detectors. J. Comp. Physiol. A 2001, 187, 303–312. [Google Scholar] [CrossRef]
- Schaber, C.F.; Barth, F.G. Spider joint hair sensilla: Adaptation to proprioreceptive stimulation. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2015, 201, 235–248. [Google Scholar] [CrossRef]
- Gierlasiński, G.; Lis, B.; Kaszyca-Taszakowska, N.; Tasza-kowski, A. Damsel bugs (Hemiptera: Heteroptera: Nabidae) of Poland: Identification key, distribution and bionomy. In Monographs of the Upper Silesian Museum 17, 1st ed.; Roland, D., Katarzyna, K., Eds.; Upper Silesian Museum: Bytom, Poland, 2020; pp. 1–100. [Google Scholar]
- Mariusz, K.; Jagna, K.; Karina, W. Morphology of the European species of the aphid genus Eulachnus (Hemiptera: Aphididae: Lachninae)– A SEM comparative and integrative study. Micron 2015, 76, 23–36. [Google Scholar]
- Schneeberg, K.; Bauernfeind, R.; Pohl, H. Comparison of cleaning methods for delicate insect specimens for scanning electron microscopy. Microsc. Res. Tech. 2017, 80, 1199–1204. [Google Scholar] [CrossRef] [PubMed]
- DeGraef, M. The Transmission electron microscopy. In Introduction to Conventional Transmission Electron Microscopy, 4th ed.; Cambridge University Press: New York, NY, USA, 2003; Volume 1, pp. 136–233. [Google Scholar]
- Billingsley, P.F.; Lehane, M.J. Structure and ultrastructure of the insect midgut. In Biology of the insect midgut. Springer 1996, 3, 3–30. [Google Scholar]
- Phillips, P.J.; Brandes, M.C.; Mills, M.J.; DeGraef, M. Diffraction contrast STEM of Dislocations: Imaging and Simulations. Ultramicroscopy 2011, 111, 1483–1487. [Google Scholar] [CrossRef]
- Kanou, M.; Osawa, T.; Shimozawa, T. Mechanical polarization in the air-current sensory hair of a cricket. Experientia 1989, 45, 1082–1083. [Google Scholar] [CrossRef]
- Shimozawa, T.; Kumagai, T.; Baba, Y. Structural Scaling and Functional Design of the Cercal Wind Receptor. J. Comp. Physiol. 1998, 183, 171–186. [Google Scholar] [CrossRef]
- Kumagai, T.; Shimozawa, T.; Baba, Y. Mobilities of the Cercal Wind-Receptor Hairs of the Cricket, Gryllus bimaculatus. J. Comp. Physiol. 1998, a183, 7–21. [Google Scholar] [CrossRef]
- French, A.S.; Torkkeli, P.H. Mechanoreceptors. In Encyclopedia of Neuroscience; Academic Press: Cambridge, MA, USA, 2009; pp. 689–695. [Google Scholar] [CrossRef]
- Singh, S.; Melnik, R. Microtubule Biomechanics and the Effect of Degradation of Elastic Moduli. Comput. Sci. ICCS 2020, 12142, 348–358. [Google Scholar] [CrossRef]
- Vincent, J.F.; Wegst, U.G. Design and Mechanical Properties of Insect Cuticle. Arthropod Struct. Dev. 2004, 33, 187–199. [Google Scholar] [CrossRef]
- Öztürk-Çolak, A.; Moussian, B.; Araújo, S.J.; Casanova, J. A feedback Mechanism Converts Individual Cell Features into A Supracellular ECM Structure in Drosophila Trachea. Elife 2016, 2, 5–8. [Google Scholar] [CrossRef]
- Rutkowski, A.; Evers, J.; Miller, M. A Biologically Inspired Approach to Controlling the Altitude and Groundspeed of an Aerial Vehicle Using Passive Sensors. In Proceedings of the Aerospace Conference and AIAA Unmanned. Unlimited Conference, Seattle, WA, USA, 6–9 April 2009. [Google Scholar]
- Guillet, J.C.; Bernard, J.; Coillot, J.P.; Callec, J.J. Electrical Properties of the Dendrite in An Insect Mechanoreceptor: Effects of Antidromic or Direct Electrical Stimulation. J. Insect Physiol. 1980, 26, 755–762. [Google Scholar] [CrossRef]
- Shimozawa, T.; Kanou, M. Varieties of filiform hairs: Range Fractionation by Sensory Afferents and Cereal Interneurons of A Cricket. J. Comp. Physiol 1984, 155, 485–493. [Google Scholar] [CrossRef]
- Barth, F.G. Mechanics to pre-process information for the fine tuning of mechanoreceptors. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2019, 205, 661–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnatzy, W.; Schmidt, K. Die Feinstruktur der Sinneshaare auf den Cerci von Gryllus bimaculatus Deg. (Saltatoria, Gryllidae). I. Faden- und Keulenhaare [The fine structure of the sensory hairs on the ceroi of Gryllus bimaculatus Deg. (Saltatoria, Gryllidae). I. Filamentous and club-shaped hairs]. Z. Zellforsch. Mikrosk. Anat. 1971, 122, 190–209. [Google Scholar]
- Keil, T.A. Functional morphology of insect mechanoreceptors. Microsc. Res. Tech. 1997, 39, 506–531. [Google Scholar] [CrossRef]
- Theiß, J. Mechanoreceptive bristles on the head of the blowfly: Mechanics and electrophysiology of the macrochaetae. J. Comp. Physiol. 1979, 132, 55–68. [Google Scholar] [CrossRef]
- Palka, J.; Edwards, J.S. The cerci and abdominal giant fibres of the house cricket, Acheta domesticus. II. Regeneration and effects of chronic deprivation. Proc. R Soc. Lond. B Biol. Sci. 1974, 185, 105–121. [Google Scholar] [CrossRef]
- Bischof, H.J. The club-shaped sensilla on the cerci of the cricket Gryllus bimaculatus as gravity receptors. J. Comp. physiol. 1975, 98, 277–288. [Google Scholar] [CrossRef]
- Guo, J.S.; Wang, X.Q.; Li, D.T.; Song, D.D.; Zhang, C.X. Three-dimensional architecture of a mechanoreceptor in the brown planthopper, Nilaparvata lugens, revealed by FIB-SEM. Cell Tissue Res. 2020, 379, 487–495. [Google Scholar] [CrossRef]
- Schmidt-Hieber, C.; Jonas, P.; Bischofberger, J. Action potential initiation and propagation in hippocampal mossy fibre axons. J. Physiol. 2008, 586, 1849–1857. [Google Scholar] [CrossRef]
- Kennedy, M.J.; Ehlers, M.D. Mechanisms and function of dendritic exocytosis. Neuron 2011, 69, 856–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuntz, H.; Forstner, F.; Haag, J.; Borst, A. The morphological identity of insect dendrites. PLoS Comput. Biol. 2008, 4, e1000251. [Google Scholar] [CrossRef] [Green Version]
- Thurm, U. Mechanoreceptors in The Cuticle of The Honeybee: Fine Structure and Stimulus Mechanism. Science 1964, 145, 1063–1065. [Google Scholar] [CrossRef]
- Blickhan, R.; Barth, F.G. Strains in the exoskeleton of spiders. J. Comp. Physiol. 1985, 157, 115–147. [Google Scholar] [CrossRef]
- Moran, D.T.; Varela, F.G. Microtubules and sensory transduction. Proc. Natl. Acad. Sci. USA 1971, 68, 757–760. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, M.M.; Henderson, C.G.; Mackie, J.B.; Lane, E.B.; Garrod, D.R.; Tucker, J.B. Keratin filament deployment and cytoskeletal networking in a sensory epithelium that vibrates during hearing. Cell Motil Cytoskelet. 1998, 41, 138–153. [Google Scholar] [CrossRef]
- Anne, D.; Jaime, G.A.; David, P. Corey, Insect mechanoreception: What a long, strange TRP it’s been. Curr. Biol. 2000, 10, R384–R387. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.P.; Krueger, S.; Heys, J.J.; Gedeon, T. Quantitative characterization of the filiform mechanosensory hair array on the cricket cercus. PLoS ONE 2011, 6, e27873. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Madrid, J.; Howard, J. The microtubule-based cytoskeleton is a component of a mechanical signalling pathway in fly campaniform receptors. Biophys. J. 2014, 107, 2767–2774. [Google Scholar] [CrossRef] [Green Version]
- Keil, T. Die Makrochaeten auf dem Thorax vonCalliphora vicina Robineau-Desvoidy (Calliphoridae, Diptera). Zoomorphologie 1978, 90, 151–180. [Google Scholar] [CrossRef]
- Cameron, M.H.; David, N.P.; Clayton, B.; Aarat, P.K.; Patricia, J.O.; Chenxi, H.; Tuszynski, J.A.; Warkentin, B.J.; Hegmann, F.A. Disassembly of microtubules by intense terahertz pulses. Biomed. Opt. Express 2021, 12, 5812–5828. [Google Scholar]
- Tuthill, J.C.; Wilson, R.I. Mechanosensation and Adaptive Motor Control in Insects. Curr. Biol. 2016, 26, R1022–R1038. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part | Youngs Modulus | Poisson’s Ratio | Density | Ref. |
|---|---|---|---|---|
| Socket and cuticle | 2 Gpa | 0.3 | 1100 kg/m3 | [25] |
| Hair | 1.8 Gpa | 0.4 | 1100 kg/m3 | [26] |
| Joint membrane | 1.6 Gpa | 0.2 | 1100 kg/m3 | [27] |
| Dendrite sheath | 1 gpa | 0.49 | 1100 kg/m3 | [28] |
| Suspension fibres | 500 kpa | 0.1 | 1100 kg/m3 | [29] |
| Lymph space | 10 Kpa | 0.1 | 1100 kg/m3 | [30] |
| Socket septum | 0.01 Gpa | 0.1 | 1100 kg/m3 | [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakilam, S.; Gaidys, R.; Brożek, J. Ultrastructure of a Mechanoreceptor of the Trichoid Sensilla of the Insect Nabis rugosus: Stimulus-Transmitting and Bio-Sensory Architecture. Bioengineering 2023, 10, 97. https://doi.org/10.3390/bioengineering10010097
Chakilam S, Gaidys R, Brożek J. Ultrastructure of a Mechanoreceptor of the Trichoid Sensilla of the Insect Nabis rugosus: Stimulus-Transmitting and Bio-Sensory Architecture. Bioengineering. 2023; 10(1):97. https://doi.org/10.3390/bioengineering10010097
Chicago/Turabian StyleChakilam, Shashikanth, Rimvydas Gaidys, and Jolanta Brożek. 2023. "Ultrastructure of a Mechanoreceptor of the Trichoid Sensilla of the Insect Nabis rugosus: Stimulus-Transmitting and Bio-Sensory Architecture" Bioengineering 10, no. 1: 97. https://doi.org/10.3390/bioengineering10010097