
Mercury Bioaccumulation in Benthic Invertebrates: From Riverine Sediments to Higher Trophic Levels


Abstract
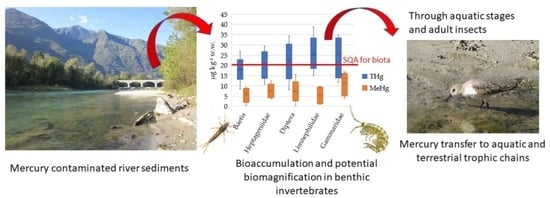
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sediment and Benthic Invertebrate Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Genus/Species | Feeding Habits | FFG |
|---|---|---|---|---|
| Ephemeroptera | Ephemerellidae | Serratella ignita | grazer/gatherer | grazers |
| Ephemeroptera | Baetidae | Baetis rhodani + Baetis alpinus/lutheri gr. | grazer/gatherer | |
| Plecoptera | Leuctridae | Leuctra sp. | grazer/gatherer/shredder | |
| Ephemeroptera | Heptageniidae | Ecdyonurus venosus | grazer/gatherer/omnivore a | gatherers-predators |
| Plecoptera | Perlodidae | Dictyogenus + Isoperla | predator (grazer/gatherer/shredder) | |
| Diptera | Limoniidae | Hexatoma sp. | predator (shredder) | |
| Diptera | Tipulidae b | Tipula lateralis | gatherer (shredder) | |
| Diptera | Tabanidae b | Chrysops sp. + Haematopota sp. | predator (gatherer) | |
| Trichoptera | Limnephilidae b | Allogamus sp. | shredder (grazer/predator) | shredders |
| Amphipoda | Gammaridae | Echinogammarus sp. | shredder | |
| Ephemeroptera | Ephemeridae | Ephemera danica | active filter feeder (gatherer) | active filter feeders |
| Oligochaeta | Lumbricidae | Eisenia tetraedra | gatherer | gatherers |
| Trichoptera | Rhyacophilidae | Rhyacophila sp. | predator | small sized predators |
| Plecoptera | Perlidae | Perla sp. | predator (grazer) | large sized predators |
2.3. THg and MeHg Analysis
2.4. Data Analysis
3. Results and Discussion
3.1. THg in Sediments and Biota
3.2. MeHg in Sediments and Biota
3.3. Relation between BSAF and Environmental Variables
3.4. Environmental Risk
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsui, M.T.K.; Finlay, J.C.; Nater, E.A. Mercury Bioaccumulation in a Stream Network. Environ. Sci. Technol. 2009, 43, 7016–7022. [Google Scholar] [CrossRef] [PubMed]
- Ward, D.M.; Nislow, K.H.; Folt, C.L. Bioaccumulation Syndrome: Identifying Factors That Make Some Stream Food Webs Prone to Elevated Mercury Bioaccumulation: Bioaccumulation Syndrome. Ann. N. Y. Acad. Sci. 2010, 1195, 62–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amos, H.M.; Jacob, D.J.; Kocman, D.; Horowitz, H.M.; Zhang, Y.; Dutkiewicz, S.; Horvat, M.; Corbitt, E.S.; Krabbenhoft, D.P.; Sunderland, E.M. Global Biogeochemical Implications of Mercury Discharges from Rivers and Sediment Burial. Environ. Sci. Technol. 2014, 48, 9514–9522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossa, D.; Garnier, C.; Buscail, R.; Elbaz-Poulichet, F.; Mikac, N.; Patel-Sorrentino, N.; Tessier, E.; Rigaud, S.; Lenoble, V.; Gobeil, C. A Michaelis–Menten Type Equation for Describing Methylmercury Dependence on Inorganic Mercury in Aquatic Sediments. Biogeochemistry 2014, 119, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Bigham, G.N.; Murray, K.J.; Masue-Slowey, Y.; Henry, E.A. Biogeochemical Controls on Methylmercury in Soils and Sediments: Implications for Site Management: Geochemical Controls on Mercury Methylation. Integr. Environ. Assess. Manag. 2017, 13, 249–263. [Google Scholar] [CrossRef]
- Ullrich, S.M.; Tanton, T.W.; Abdrashitova, S.A. Mercury in the Aquatic Environment: A Review of Factors Affecting Methylation. Crit. Rev. Environ. Sci. Technol. 2001, 31, 241–293. [Google Scholar] [CrossRef]
- Amirbahman, A.; Massey, D.I.; Lotufo, G.; Steenhaut, N.; Brown, L.E.; Biedenbach, J.M.; Magar, V.S. Assessment of Mercury Bioavailability to Benthic Macroinvertebrates Using Diffusive Gradients in Thin Films (DGT). Environ. Sci. Process. Impacts 2013, 15, 2104–2114. [Google Scholar] [CrossRef]
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.M. Biomagnification of Mercury in Aquatic Food Webs: A Worldwide Meta-Analysis. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef]
- Coelho, J.P.; Mieiro, C.L.; Pereira, E.; Duarte, A.C.; Pardal, M.A. Mercury Biomagnification in a Contaminated Estuary Food Web: Effects of Age and Trophic Position Using Stable Isotope Analyses. Mar. Pollut. Bull. 2013, 69, 110–115. [Google Scholar] [CrossRef]
- Xu, X.; Bryan, A.L.; Mills, G.L.; Korotasz, A.M. Mercury Speciation, Bioavailability, and Biomagnification in Contaminated Streams on the Savannah River Site (SC, USA). Sci. Total Environ. 2019, 668, 261–270. [Google Scholar] [CrossRef]
- Rennie, M.; Collins, N.; Purchase, C.F.; Tremblay, A. Predictive Models of Benthic Invertebrate Methylmercury in Ontario and Quebec Lakes. Can. J. Fish. Aquat. Sci. 2005, 62, 2770–2783. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.L.; Linehan, J.C.; Murray, D.W.; Prell, W.L. Indicators of Sediment and Biotic Mercury Contamination in a Southern New England Estuary. Mar. Pollut. Bull. 2012, 64, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Lepom, P.; Irmer, U.; Wellmitz, J. Mercury Levels and Trends (1993–2009) in Bream (Abramis Brama L.) and Zebra Mussels (Dreissena Polymorpha) from German Surface Waters. Chemosphere 2012, 86, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Nguetseng, R.; Fliedner, A.; Knopf, B.; Lebreton, B.; Quack, M.; Rüdel, H. Retrospective Monitoring of Mercury in Fish from Selected European Freshwater and Estuary Sites. Chemosphere 2015, 134, 427–434. [Google Scholar] [CrossRef] [PubMed]
- European Environment Agency. European Waters: Assessment of Status and Pressures 2018; Publications Office: Luxembourg, 2018. [Google Scholar]
- Cid, N.; Ibáñez, C.; Palanques, A.; Prat, N. Patterns of Metal Bioaccumulation in Two Filter-Feeding Macroinvertebrates: Exposure Distribution, Inter-Species Differences and Variability across Developmental Stages. Sci. Total Environ. 2010, 408, 2795–2806. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.P.; Laporte, J.-M.; Andres, S. Factors Controlling the Bioaccumulation of Mercury, Methylmercury, Arsenic, Selenium, and Cadmium by Freshwater Invertebrates and Fish. Arch. Environ. Contam. Toxicol. 2000, 38, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.M.; Hall, B.D. Concentrations of Methylmercury in Invertebrates from Wetlands of the Prairie Pothole Region of North America. Environ. Pollut. 2012, 160, 153–160. [Google Scholar] [CrossRef]
- Jardine, T.D.; Kidd, K.A.; O’ Driscoll, N. Food Web Analysis Reveals Effects of PH on Mercury Bioaccumulation at Multiple Trophic Levels in Streams. Aquat. Toxicol. 2013, 132–133, 46–52. [Google Scholar] [CrossRef]
- Edmonds, S.T.; O’Driscoll, N.J.; Hillier, N.K.; Atwood, J.L.; Evers, D.C. Factors Regulating the Bioavailability of Methylmercury to Breeding Rusty Blackbirds in Northeastern Wetlands. Environ. Pollut. 2012, 171, 148–154. [Google Scholar] [CrossRef]
- Tsui, M.T.K.; Wang, W.-X. Uptake and Elimination Routes of Inorganic Mercury and Methylmercury in Daphnia Magna. Environ. Sci. Technol. 2004, 38, 808–816. [Google Scholar] [CrossRef]
- Gimbert, F.; Geffard, A.; Guédron, S.; Dominik, J.; Ferrari, B.J.D. Mercury Tissue Residue Approach in Chironomus riparius: Involvement of Toxicokinetics and Comparison of Subcellular Fractionation Methods. Aquat. Toxicol. 2016, 171, 1–8. [Google Scholar] [CrossRef]
- McGeer, J.C.; Brix, K.V.; Skeaff, J.M.; DeForest, D.K.; Brigham, S.I.; Adams, W.J.; Green, A. Inverse Relationship between Bioconcentration Factor and Exposure Concentration for Metals: Implications for Hazard Assessment of Metals in the Aquatic Environment. Environ. Toxicol. Chem. 2003, 22, 1017–1037. [Google Scholar] [CrossRef] [PubMed]
- Chételat, J.; Amyot, M.; Cloutier, L.; Poulain, A. Metamorphosis in Chironomids, More than Mercury Supply, Controls Methylmercury Transfer to Fish in High Arctic Lakes. Environ. Sci. Technol. 2008, 42, 9110–9115. [Google Scholar] [CrossRef] [PubMed]
- Kraus, J.M.; Walters, D.M.; Wesner, J.S.; Stricker, C.A.; Schmidt, T.S.; Zuellig, R.E. Metamorphosis Alters Contaminants and Chemical Tracers in Insects: Implications for Food Webs. Environ. Sci. Technol. 2014, 48, 10957–10965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marziali, L.; Valsecchi, L. Mercury Bioavailability in Fluvial Sediments Estimated Using Chironomus riparius and Diffusive Gradients in Thin-Films (DGT). Environments 2021, 8, 7. [Google Scholar] [CrossRef]
- Guilizzoni, P.; Levine, S.N.; Manca, M.; Marchetto, A.; Lami, A.; Ambrosetti, W.; Brauer, A.; Gerli, S.; Carrara, E.A.; Rolla, A.; et al. Ecological Effects of Multiple Stressors on a Deep Lake (Lago Maggiore, Italy) Integrating Neo and Palaeolimnological Approaches. J. Limnol. 2012, 71, 1. [Google Scholar] [CrossRef]
- Guzzella, L.M.; Novati, S.; Casatta, N.; Roscioli, C.; Valsecchi, L.; Binelli, A.; Parolini, M.; Solcà, N.; Bettinetti, R.; Manca, M.; et al. Spatial and Temporal Trends of Target Organic and Inorganic Micropollutants in Lake Maggiore and Lake Lugano (Italian-Swiss Water Bodies): Contamination in Sediments and Biota. Hydrobiologia 2018, 824, 271–290. [Google Scholar] [CrossRef]
- Marziali, L.; Guzzella, L.; Salerno, F.; Marchetto, A.; Valsecchi, L.; Tasselli, S.; Roscioli, C.; Schiavon, A. Twenty-Year Sediment Contamination Trends in Some Tributaries of Lake Maggiore (Northern Italy): Relation with Anthropogenic Factors. Environ. Sci. Pollut. Res. 2021, 28, 38193–38208. [Google Scholar] [CrossRef]
- MacDonald, D.D.; Ingersoll, C.G.; Berger, T.A. Development and Evaluation of Consensus-Based Sediment Quality Guidelines for Freshwater Ecosystems. Arch. Environ. Contam. Toxicol. 2000, 39, 20–31. [Google Scholar] [CrossRef]
- Pisanello, F.; Marziali, L.; Rosignoli, F.; Poma, G.; Roscioli, C.; Pozzoni, F.; Guzzella, L. In Situ Bioavailability of DDT and Hg in Sediments of the Toce River (Lake Maggiore Basin, Northern Italy): Accumulation in Benthic Invertebrates and Passive Samplers. Environ. Sci. Pollut. Res. 2016, 23, 10542–10555. [Google Scholar] [CrossRef]
- Marziali, L.; Rosignoli, F.; Drago, A.; Pascariello, S.; Valsecchi, L.; Rossaro, B.; Guzzella, L. Toxicity Risk Assessment of Mercury, DDT and Arsenic Legacy Pollution in Sediments: A Triad Approach under Low Concentration Conditions. Sci. Total Environ. 2017, 593–594, 809–821. [Google Scholar] [CrossRef] [PubMed]
- Belfiore, C. Efemerotteri (Ephemeroptera). In Guide Per Il Riconoscimento Delle Specie Animali Delle Acque Interne Italiane; Consiglio Nazionale Delle Ricerche (CNR) : Rome, Italy, 1983; Volume 24. [Google Scholar]
- Rivosecchi, L. Ditteri (Diptera). In Guide Per Il Riconoscimento Delle Specie Animali Delle Acque Interne Italiane; Consiglio Nazionale Delle Ricerche (CNR): Rome, Italy, 1984; Volume 28. [Google Scholar]
- Campaioli, S.; Ghetti, P.F.; Minelli, A.; Ruffo, S. Manuale per Il Riconoscimento Dei Macroinvertebrati Delle Acque Dolci Italiane; Provincia Autonoma di Trento: Trento, Italy, 1994; Volume 1. [Google Scholar]
- Campaioli, S.; Ghetti, P.F.; Minelli, A.; Ruffo, S. Manuale per Il Riconoscimento Dei Macroinvertebrati Delle Acque Dolci Italiane; Provincia Autonoma di Trento: Trento, Italy, 1999; Volume 2, ISBN 88-7702-089-X. [Google Scholar]
- Vane, C.H.; Kim, A.W.; Emmings, J.F.; Turner, G.H.; Moss-Hayes, V.; Lort, J.A.; Williams, P.J. Grain Size and Organic Carbon Controls Polyaromatic Hydrocarbons (PAH), Mercury (Hg) and Toxicity of Surface Sediments in the River Conwy Estuary, Wales, UK. Mar. Pollut. Bull. 2020, 158, 111412. [Google Scholar] [CrossRef]
- Kersten, M.; Smedes, F. Normalization Procedures for Sediment Contaminants in Spatial and Temporal Trend Monitoring. J. Environ. Monitor. 2002, 4, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding Type and Development Drive the Ingestion of Microplastics by Freshwater Invertebrates. Sci. Rep. 2017, 7, 17006. [Google Scholar] [CrossRef] [Green Version]
- Schmidt-Kloiber, A.; Hering, D. Www.freshwaterecology.info—An Online Tool That Unifies, Standardises and Codifies More than 20,000 European Freshwater Organisms and Their Ecological Preferences. Ecol. Indic. 2015, 53, 271–282. Available online: https://www.freshwaterecology.info/ (accessed on 3 May 2021). [CrossRef]
- Riva-Murray, K.; Chasar, L.C.; Bradley, P.M.; Burns, D.A.; Brigham, M.E.; Smith, M.J.; Abrahamsen, T.A. Spatial Patterns of Mercury in Macroinvertebrates and Fishes from Streams of Two Contrasting Forested Landscapes in the Eastern United States. Ecotoxicology 2011, 20, 1530–1542. [Google Scholar] [CrossRef] [Green Version]
- US-EPA. Method 7473—Mercury in Solids and Solutions by Thermal Decomposition, Amalgamation, and Atomic Absorption Spectrophotometry. Revision 0; US Environmental Protection Agency: Washington, DC, USA, 1998.
- US-EPA. Methods for the Determination of Total Organic Carbon (TOC) in Soils and Sediments; US Environmental Protection Agency, Ecological Risk Assessment Support Center, Office of Research and Development NCEA-C-1282, EMASC-001: Washington, DC, USA, 2002.
- De Vos, B.; Lettens, S.; Muys, B.; Deckers, J.A. Walkley-Black Analysis of Forest Soil Organic Carbon: Recovery, Limitations and Uncertainty. Soil Use Manag. 2007, 23, 221–229. [Google Scholar] [CrossRef]
- Jones, H.J.; Swadling, K.M.; Butler, E.C.V.; Macleod, C.K. Complex Patterns in Fish—Sediment Mercury Concentrations in a Contaminated Estuary: The Influence of Selenium Co-Contamination? Estuar. Coast. Shelf Sci. 2014, 137, 14–22. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Xue, W.; Kwon, S.Y.; Grasby, S.E.; Sunderland, E.M.; Pan, X.; Sun, R.; Zhou, T.; Yan, H.; Yin, R. Anthropogenic Influences on Mercury in Chinese Soil and Sediment Revealed by Relationships with Total Organic Carbon. Environ. Pollut. 2019, 255, 113186. [Google Scholar] [CrossRef] [PubMed]
- Chibunda, R.T. Chronic Toxicity of Mercury (HgCl2) to the Benthic Midge Chironomus riparius. Int. J. Environ. Res. 2009, 3, 455–462. [Google Scholar] [CrossRef]
- Žižek, S.; Horvat, M.; Gibičar, D.; Fajon, V.; Toman, M.J. Bioaccumulation of Mercury in Benthic Communities of a River Ecosystem Affected by Mercury Mining. Sci. Total Environ. 2007, 377, 407–415. [Google Scholar] [CrossRef]
- Roig, N.; Sierra, J.; Ortiz, J.D.; Merseburger, G.; Schuhmacher, M.; Domingo, J.L.; Nadal, M. Integrated Study of Metal Behavior in Mediterranean Stream Ecosystems: A Case-Study. J. Hazard. Mater. 2013, 263, 122–130. [Google Scholar] [CrossRef]
- George, B.M.; Batzer, D. Spatial and Temporal Variations of Mercury Levels in Okefenokee Invertebrates: Southeast Georgia. Environ. Pollut. 2008, 152, 484–490. [Google Scholar] [CrossRef] [PubMed]
- St. Louis, V.L.; Rudd, J.W.M.; Kelly, C.A.; Hall, B.D.; Rolfhus, K.R.; Scott, K.J.; Lindberg, S.E.; Dong, W. Importance of the Forest Canopy to Fluxes of Methyl Mercury and Total Mercury to Boreal Ecosystems. Environ. Sci. Technol. 2001, 35, 3089–3098. [Google Scholar] [CrossRef] [PubMed]
- Fantozzi, L.; Guerrieri, N.; Manca, G.; Orrù, A.; Marziali, L. An Integrated Investigation of Atmospheric Gaseous Elemental Mercury Transport and Dispersion Around a Chlor-Alkali Plant in the Ossola Valley (Italian Central Alps). Toxics 2021, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Razavi, N.R.; Ridal, J.J.; de Wit, W.; Hickey, M.B.C.; Campbell, L.M.; Hodson, P.V. Ebullition Rates and Mercury Concentrations in St. Lawrence River Sediments and a Benthic Invertebrate. Environ. Toxicol. Chem. 2013, 32, 857–865. [Google Scholar] [CrossRef]
- Covelli, S.; Faganeli, J.; Horvat, M.; Brambati, A. Mercury Contamination of Coastal Sediments as the Result of Long-Term Cinnabar Mining Activity (Gulf of Trieste, Northern Adriatic Sea). Appl. Geochem. 2001, 16, 541–558. [Google Scholar] [CrossRef]
- Díaz-Jaramillo, M.; Muñoz, C.; Rudolph, I.; Servos, M.; Barra, R. Seasonal Mercury Concentrations and Δ15N and Δ13C Values of Benthic Macroinvertebrates and Sediments from a Historically Polluted Estuary in South Central Chile. Sci. Total Environ. 2013, 442, 198–206. [Google Scholar] [CrossRef]
- DeForest, D.K.; Brix, K.V.; Adams, W.J. Assessing Metal Bioaccumulation in Aquatic Environments: The Inverse Relationship between Bioaccumulation Factors, Trophic Transfer Factors and Exposure Concentration. Aquat. Toxicol. 2007, 84, 236–246. [Google Scholar] [CrossRef]
- International Commission for the Protection of the Italian-Swiss Waters (CIPAIS). Indagini Sulle Sostanze Pericolose Nell’ecosistema Del Lago Maggiore. Programma 2016–2018. Rapporto Annuale 2017; CIPAIS: Verbania, Italy, 2018; Available online: http://www.cipais.org/ (accessed on 19 May 2021).
- International Commission for the Protection of the Italian-Swiss Waters (CIPAIS). Indagini Sulle Sostanze Pericolose Nell’ecosistema Del Lago Maggiore. Programma 2019–2021. Rapporto Annuale 2019; CIPAIS: Verbania, Italy, 2020; Available online: http://www.cipais.org/ (accessed on 19 May 2021).
- Rossaro, B.; Gaggino, G.F.; Marchetti, F. Accumulation of Mercury in Larvae and Adults, Chironomus riparius (Meigen). Bull. Environ. Contam. Toxicol. 1986, 37, 402–406. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marziali, L.; Roscioli, C.; Valsecchi, L. Mercury Bioaccumulation in Benthic Invertebrates: From Riverine Sediments to Higher Trophic Levels. Toxics 2021, 9, 197. https://doi.org/10.3390/toxics9090197
Marziali L, Roscioli C, Valsecchi L. Mercury Bioaccumulation in Benthic Invertebrates: From Riverine Sediments to Higher Trophic Levels. Toxics. 2021; 9(9):197. https://doi.org/10.3390/toxics9090197
Chicago/Turabian StyleMarziali, Laura, Claudio Roscioli, and Lucia Valsecchi. 2021. "Mercury Bioaccumulation in Benthic Invertebrates: From Riverine Sediments to Higher Trophic Levels" Toxics 9, no. 9: 197. https://doi.org/10.3390/toxics9090197