
Thyroid Hormone Receptor Agonistic and Antagonistic Activity of Newly Synthesized Dihydroxylated Polybrominated Diphenyl Ethers: An In Vitro and In Silico Coactivator Recruitment Study
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
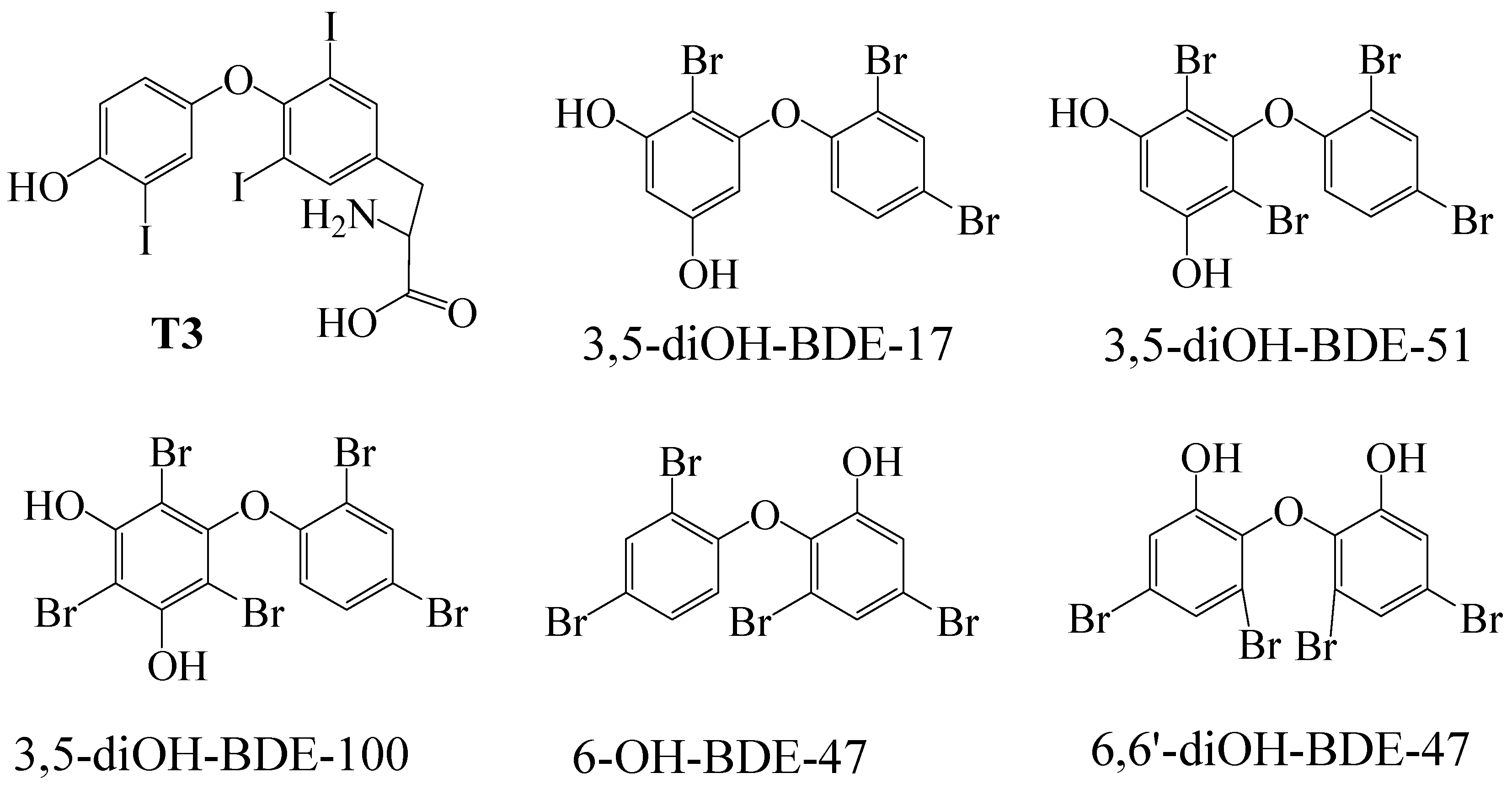
2.1. Chemicals
2.2. Coactivator Recruitment Assay
2.3. In Silico Simulations
3. Results and Discussion
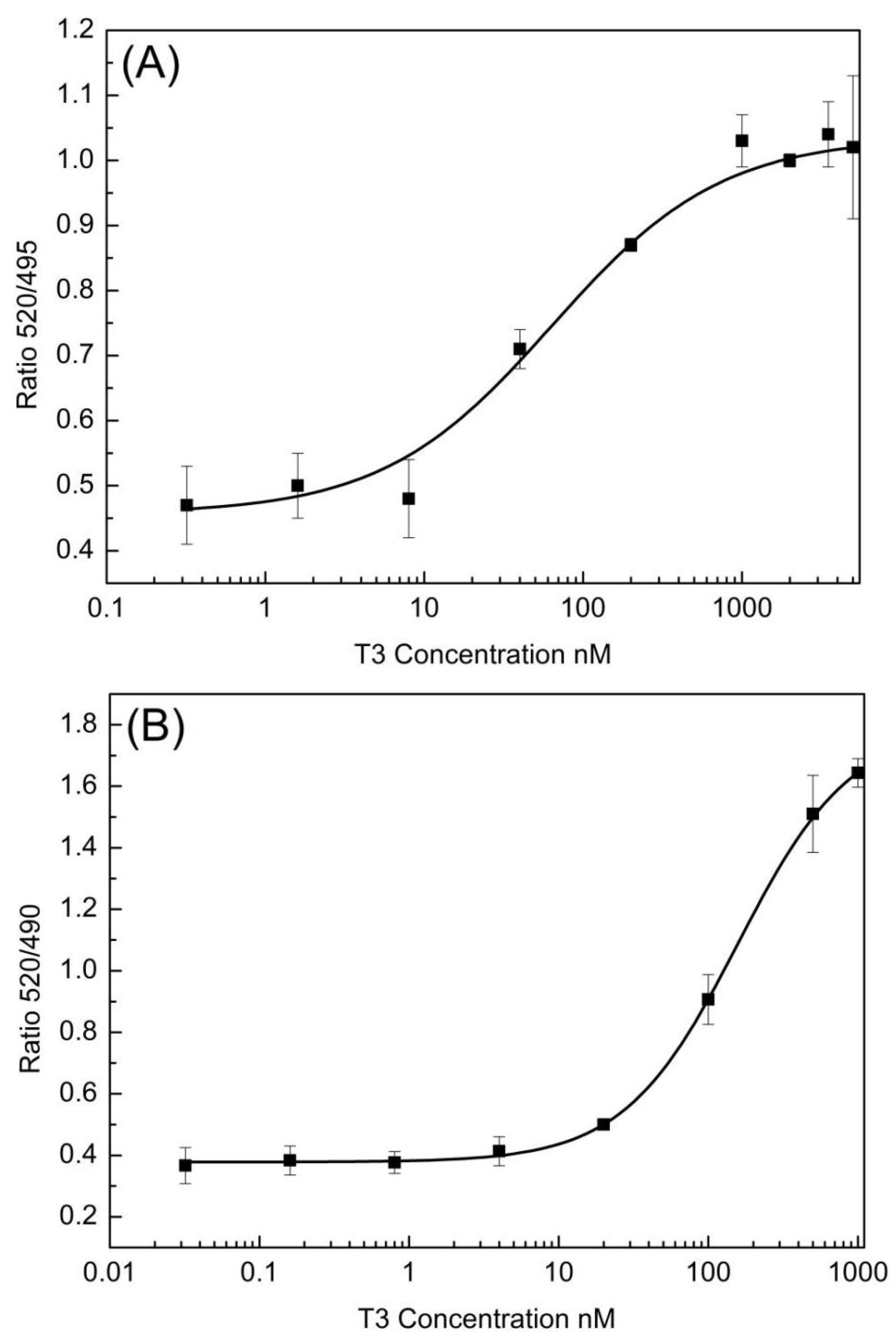
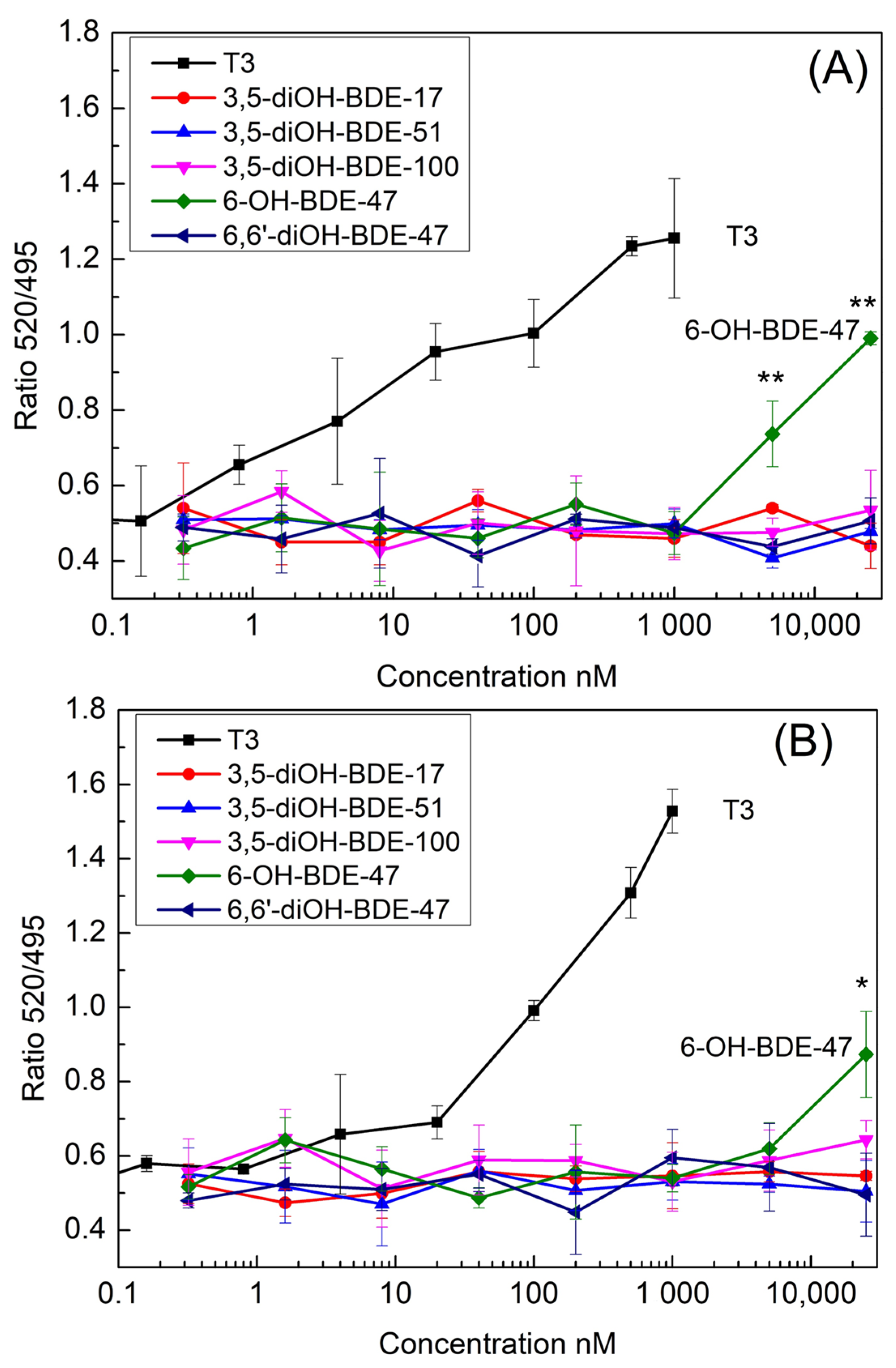
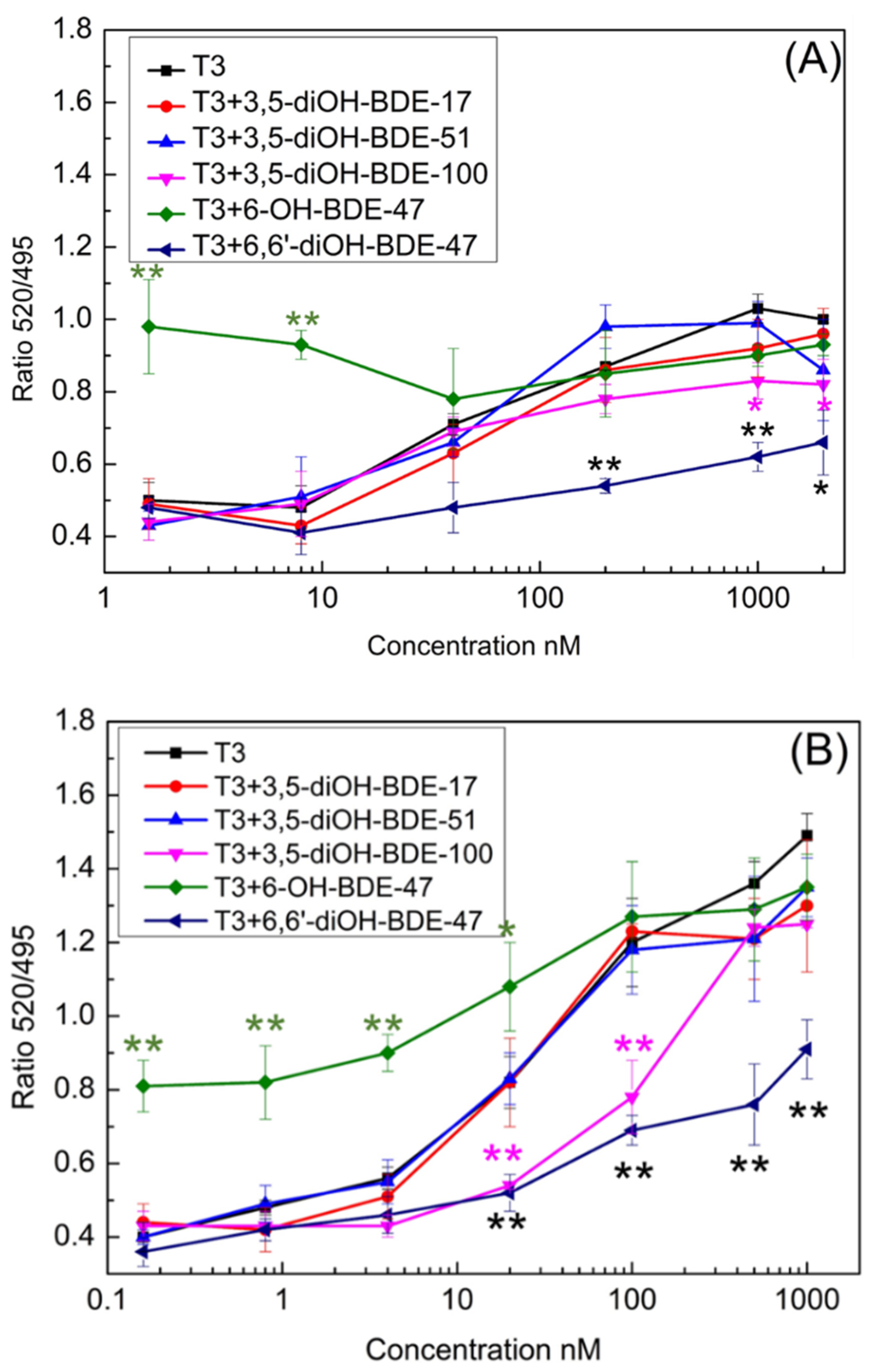
3.1. TR Coactivator Recruitment Assay for OH/diOH-PBDEs TR Agonists Activity
3.2. OH/diOH-PBDEs Competitive Binding with T3 for TR Antagonistic Activity Assay
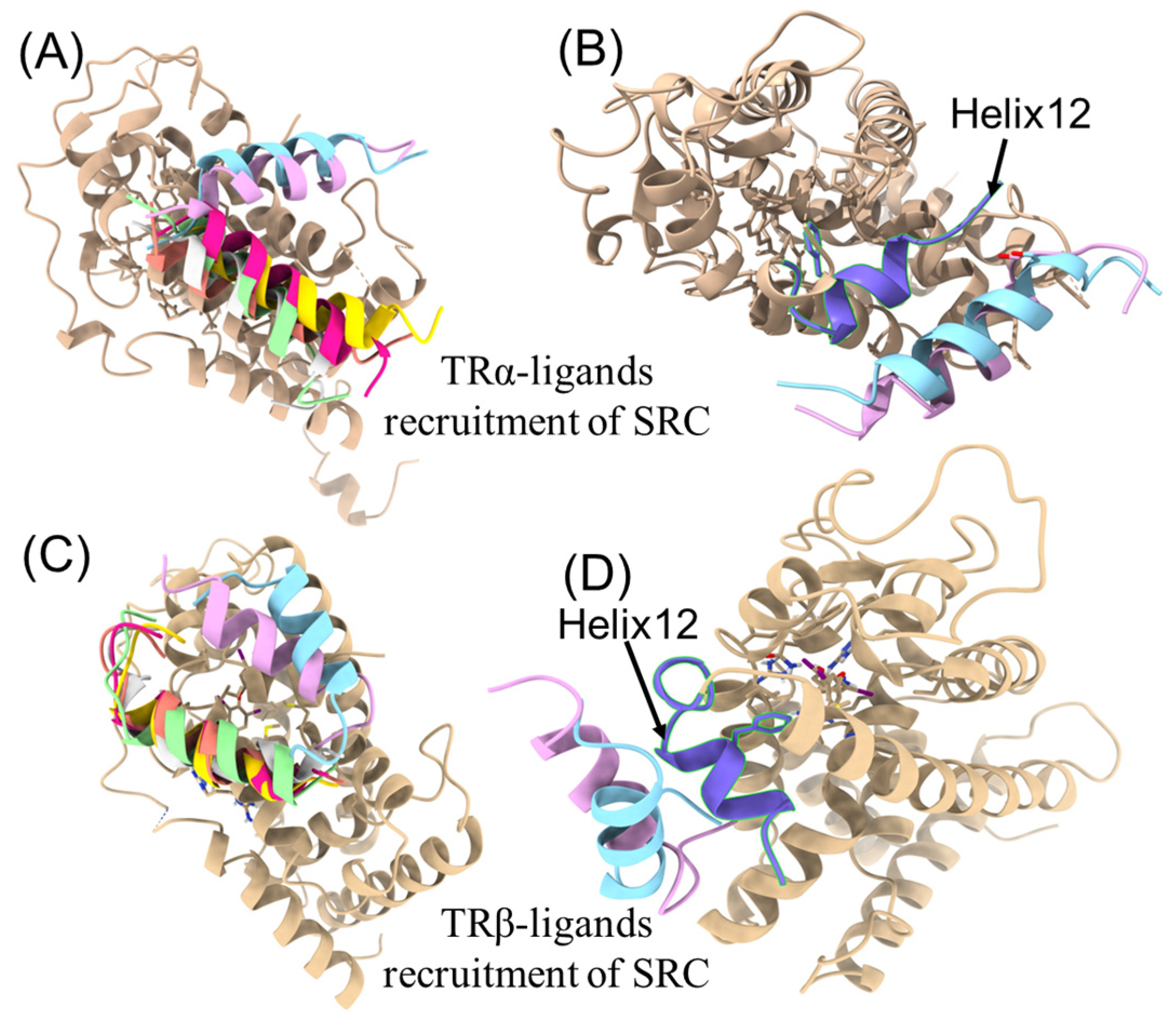
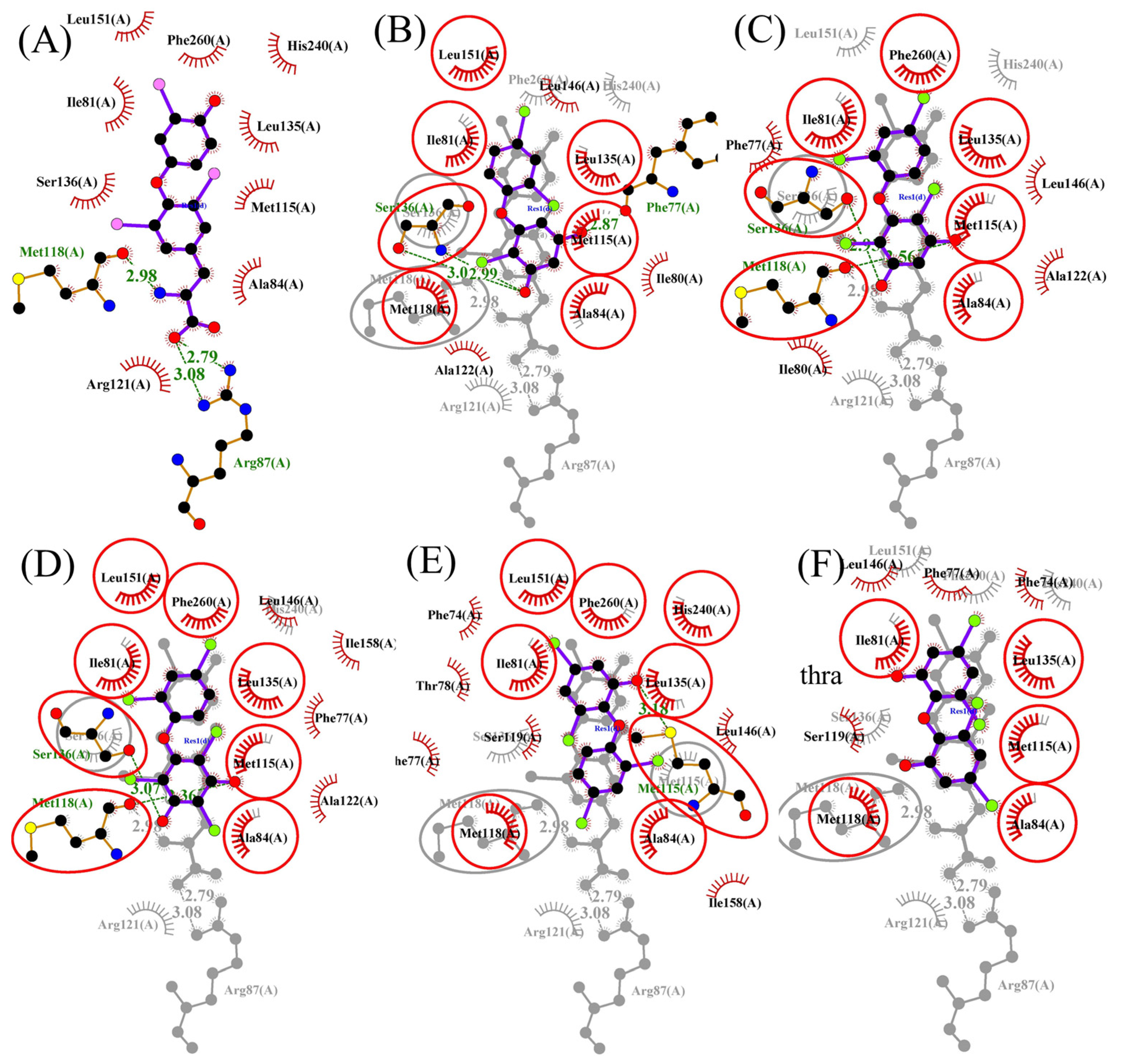
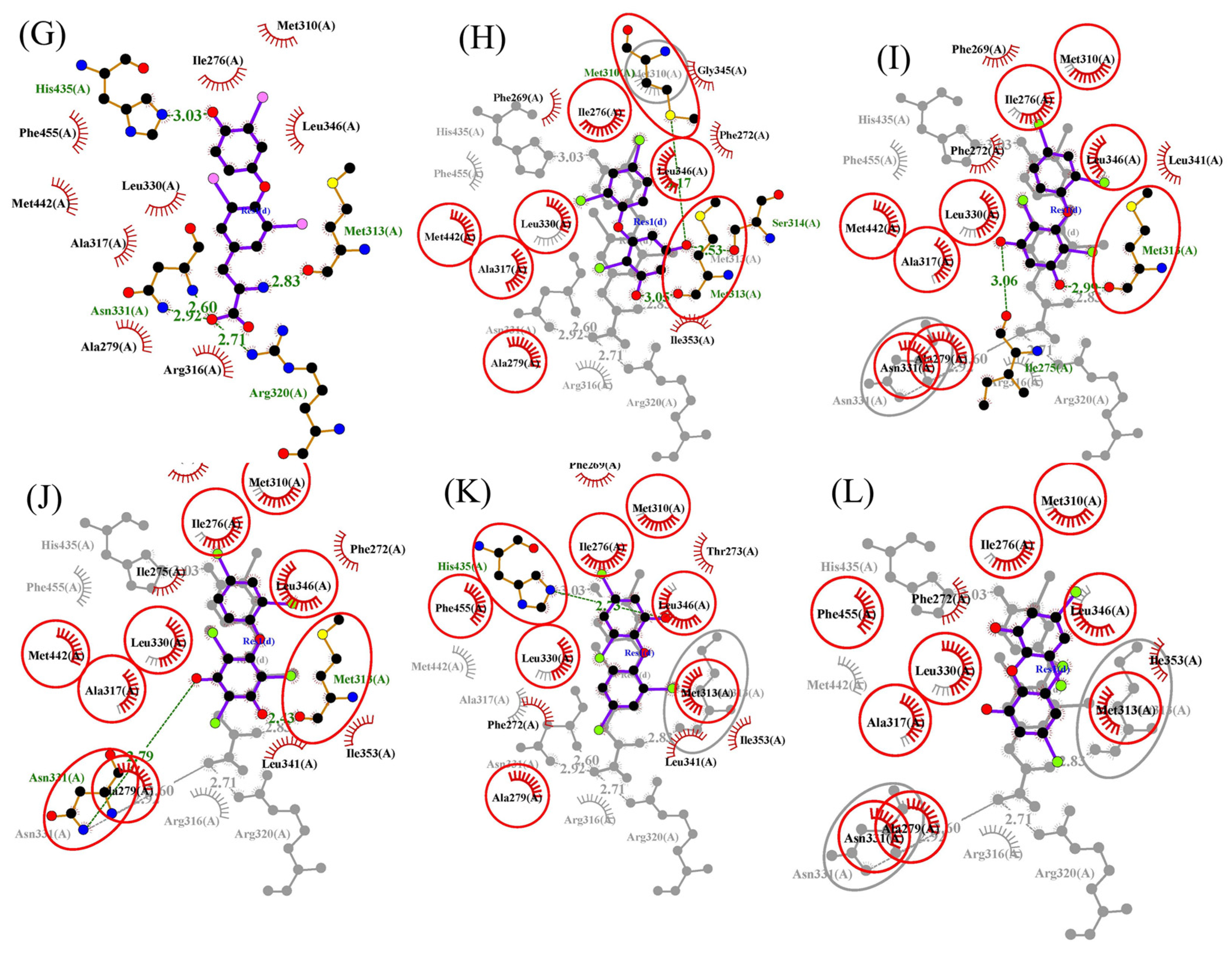
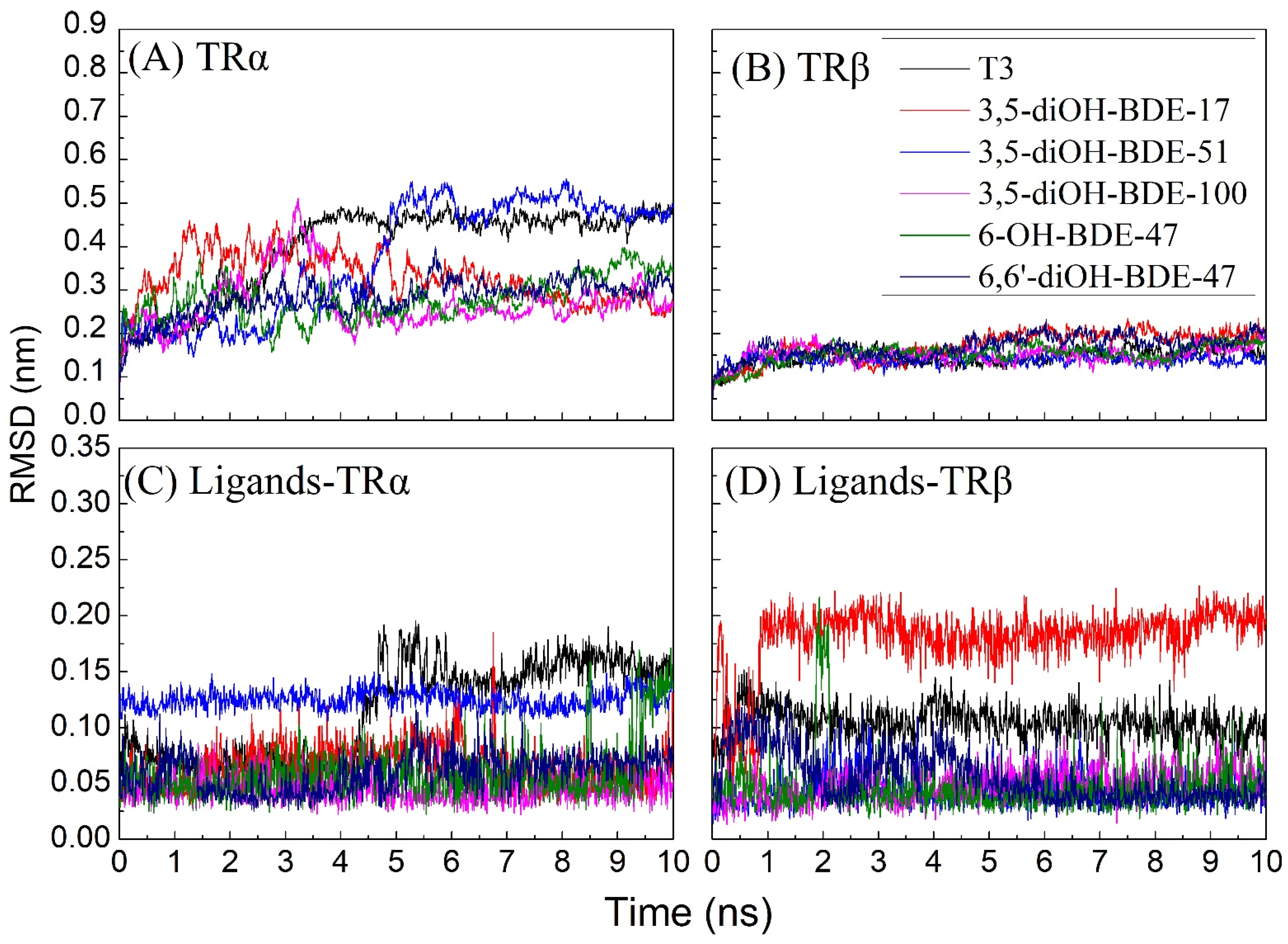
3.3. Molecular Docking and Molecular Dynamic Simulations of OH/diOH-PBDEs with TR
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbasi, G.; Li, L.; Breivik, K. Global Historical Stocks and Emissions of PBDEs. Environ. Sci. Technol. 2019, 53, 6330–6340. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Lai, C.; Xu, F.; Huang, D.; Zhang, M.; Zhou, X.; Xu, M.; Li, Y.; Li, L.; Liu, S.; et al. A review of polybrominated diphenyl ethers and novel brominated flame retardants in Chinese aquatic environment: Source, occurrence, distribution, and ecological risk assessment. Sci. Total Environ. 2023, 904, 166180. [Google Scholar] [CrossRef] [PubMed]
- UNEP. Fourth Meeting of the Conference of the Parties to the Stockholm Convention. 2009. Available online: http://Chm.Pops.Int/Programmes/Newpops/The9newpops/Tabid/672/Language/En-Us/Default.Aspx (accessed on 18 March 2024).
- UNEP. Technical Review of the Implications of Recycling Commercial Penta and Octabromodiphenyl Ethers. Annexes. Stockholm Convention Document for 6th Pop Reviewing Committee Meeting (Unep/Pops/Poprc.6/Inf/6). 2010. Available online: http://Chm.Pops.Int/Portals/0/Repository/Poprc6/Unep-Pops-Poprc.6-2.English.Pdf (accessed on 18 March 2024).
- UNEP. Eighth Meeting of the Conference of the Parties to the Stockholm Convention. 2017. Available online: http://Chm.Pops.Int/Theconvention/Conferenceoftheparties/Meetings/Cop8/Tabid/5309/Default.Aspx (accessed on 18 March 2024).
- Mizukawa, H.; Nomiyama, K.; Kunisue, T.; Watanabe, M.X.; Subramanian, A.; Iwata, H.; Ishizuka, M.; Tanabe, S. Organohalogens and their hydroxylated metabolites in the blood of pigs from an open waste dumping site in south India: Association with hepatic cytochrome P450. Environ. Res. 2015, 138, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.B.; Erratico, C.; Abdallah, M.A.E.; Negreira, N.; Luo, X.J.; Mai, B.X.; Covaci, A. In vitro metabolism of BDE-47, BDE-99, and α-, β-, γ-HBCD isomers by chicken liver microsomes. Environ. Res. 2015, 143, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Blanton, J.M.; Podell, S.; Taton, A.; Schorn, M.A.; Busch, J.; Lin, Z.; Schmidt, E.W.; Jensen, P.R.; Paul, V.J.; et al. Metagenomic discovery of polybrominated diphenyl ether biosynthesis by marine sponges. Nat. Chem. Biol. 2017, 13, 537. [Google Scholar] [CrossRef] [PubMed]
- Teuten, E.L.; Xu, L.; Reddy, C.M. Two Abundant Bioaccumulated Halogenated Compounds Are Natural Products. Science 2005, 307, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Teuten, E.L.; Reddy, C.M. Halogenated organic compounds in archived whale oil: A pre-industrial record. Environ. Pollut. 2007, 145, 668–671. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Moore, B.S. Enzymatic Synthesis of Polybrominated Dioxins from the Marine Environment. ACS Chem. Biol. 2014, 9, 1980–1984. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Li, S.J.; Huang, H.; Chen, Y.Z.; Wang, X.T. A Review of Hydroxylated and Methoxylated Brominated Diphenyl Ethers in Marine Environments. Toxics 2022, 10, 751. [Google Scholar] [CrossRef]
- Zhang, M.T.; Guo, W.; Wei, J.T.; Shi, J.H.; Zhang, J.W.; Ge, H.; Tao, H.Y.; Liu, X.W.; Hu, Q.; Cai, Z.W. Determination of newly synthesized dihydroxylated polybrominated diphenyl ethers in sea fish by gas chromatography-tandem mass spectrometry. Chemosphere 2020, 240, 124878. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, X.; Shi, W.; Yu, H.; Zhang, X.; Giesy, J.P. Identification of Thyroid Hormone Disruptors among HO-PBDEs: In Vitro Investigations and Coregulator Involved Simulations. Environ. Sci. Technol. 2016, 50, 12429–12438. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Xiang, L.; Cai, Z. Emerging Environmental Pollutants Hydroxylated Polybrominated Diphenyl Ethers: From Analytical Methods to Toxicology Research. Mass Spectrom. Rev. 2021, 40, 255–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.T.; Zhao, F.; Zhang, J.W.; Shi, J.H.; Tao, H.Y.; Ge, H.; Guo, W.; Liu, D.; Cai, Z.W. Toxicity and accumulation of 6-OH-BDE-47 and newly synthesized 6,6′-diOH-BDE-47 in early life-stages of Zebrafish (Danio rerio). Sci. Total Environ. 2021, 763, 143036. [Google Scholar] [CrossRef] [PubMed]
- Elsalini, O.A.; Rohr, K.B. Phenylthiourea disrupts thyroid function in developing zebrafish. Dev. Genes Evolut. 2003, 212, 593–598. [Google Scholar] [CrossRef]
- Usenko, C.Y.; Hopkins, D.C.; Trumble, S.J.; Bruce, E.D. Hydroxylated PBDEs induce developmental arrest in Zebrafish. Toxicol. Appl. Pharmacol. 2012, 262, 43–51. [Google Scholar] [CrossRef]
- Kojima, H.; Takeuchi, S.; Uramaru, N.; Sugihara, K.; Yoshida, T.; Kitamura, S. Nuclear Hormone Receptor Activity of Polybrominated Diphenyl Ethers and Their Hydroxylated and Methoxylated Metabolites in Transactivation Assays Using Chinese Hamster Ovary Cells. Environ. Health Perspect. 2009, 117, 1210–1218. [Google Scholar] [CrossRef]
- Ren, X.M.; Guo, L.H.; Gao, Y.; Zhang, B.T.; Wan, B. Hydroxylated polybrominated diphenyl ethers exhibit different activities on thyroid hormone receptors depending on their degree of bromination. Toxicol. Appl. Pharmacol. 2013, 268, 256–263. [Google Scholar] [CrossRef]
- Kitamura, S.; Shinohara, S.; Iwase, E.; Sugihara, K.; Uramaru, N.; Shigematsu, H.; Fujimoto, N.; Ohta, S. Affinity for Thyroid Hormone and Estrogen Receptors of Hydroxylated Polybrominated Diphenyl Ethers. J. Health Sci. 2008, 54, 607–614. [Google Scholar] [CrossRef]
- Zhang, Y.; Sanner, M.F. Docking Flexible Cyclic Peptides with AutoDock CrankPep. J. Chem. Theory Comput. 2019, 15, 5161–5168. [Google Scholar] [CrossRef]
- Zhang, Y.; Sanner, M.F. AutoDock CrankPep: Combining folding and docking to predict protein–peptide complexes. Bioinformatics 2019, 35, 5121–5127. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, A.; Hollenberg, A.N. New insights into thyroid hormone action. Pharmacol. Ther. 2017, 173, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Ocasio, C.A.; Scanlan, T.S. Characterization of thyroid hormone receptor α(TRα)-specific analogs with varying inner- and outer-ring substituents. Bioorg. Med. Chem. 2008, 16, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Levy-Bimbot, M.; Major, G.; Courilleau, D.; Blondeau, J.P.; Levi, Y. Tetrabromobisphenol-A disrupts thyroid hormone receptor alpha function in vitro: Use of fluorescence polarization to assay corepressor and coactivator peptide binding. Chemosphere 2012, 87, 782–788. [Google Scholar] [CrossRef]
- Hofmann, P.J.; Schomburg, L.; Köhrle, J. Interference of Endocrine Disrupters with Thyroid Hormone Receptor–Dependent Transactivation. Toxicol. Sci. 2009, 110, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Shanker, S.; Sanner, M.F. Predicting Protein–Peptide Interactions: Benchmarking Deep Learning Techniques and a Comparison with Focused Docking. J. Chem. Inf. Model. 2023, 63, 3158–3170. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.C.; Apriletti, J.W.; Wagner, R.L.; Feng, W.; Kushner, P.J.; Nilsson, S.; Scanlan, T.S.; West, B.L.; Fletterick, R.J.; Baxter, J.D. X-ray Crystallographic and Functional Studies of Thyroid Hormone Receptor. J. Steroid Biochem. Mol. Biol. 1998, 65, 133–141. [Google Scholar] [CrossRef]
- Khan, M.U.; Besis, A.; Li, J.; Zhang, G.; Malik, R.N. New insight into the distribution pattern, levels, and risk diagnosis of FRs in indoor and outdoor air at low- and high-altitude zones of Pakistan: Implications for sources and exposure. Chemosphere 2017, 184, 1372–1387. [Google Scholar] [CrossRef]
- Heo, S.; Lee, J.; Joo, K.; Shin, H.C.; Lee, J. Protein Loop Structure Prediction Using Conformational Space Annealing. J. Chem. Inf. Model. 2017, 57, 1068–1078. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | TRα | TRβ | ||||
|---|---|---|---|---|---|---|
| Best Binding Affinity (kcal/M) | Cluster Size | Cluster Average Binding Affinity (kcal/M) | Best Binding Affinity (kcal/M) | Cluster Size | Cluster Average Binding Affinity (kcal/M) | |
| T3 | −21.2 | 32 | −19.3 | −24.3 | 28 | −21.2 |
| 3,5-diOH-BDE-17 | −21.1 | 18 | −19.3 | −22.0 | 34 | −19.9 |
| 3,5-diOH-BDE-51 | −20.7 | 29 | −19.0 | −22.3 | 36 | −19.4 |
| 3,5-diOH-BDE-100 | −21.8 | 37 | −19.3 | −22.7 | 13 | −22.2 |
| 6-OH-BDE-47 | −21.5 | 10 | −21.1 | −25.3 | 32 | −23.4 |
| 6,6′-diOH-BDE-47 | −22.1 | 52 | −19.6 | −22.4 | 24 | −21.5 |
| Compound | Length (Å) | TRα | TRβ | ||
|---|---|---|---|---|---|
| Docking Energy (kcal/M) | Hydrogen Bonding | Docking Energy (kcal/M) | Hydrogen Bonding | ||
| T3 | 12.37 | −10.51 | Arg87, Met118 | −11.75 | Met313, Asn331, Arg320, His435 |
| 3,5-diOH-BDE-17 | 10.21 | −8.26 | Ser136, Phe77 | −8.01 | Met313, Ser314, Met310 |
| 3,5-diOH-BDE-51 | 10.41 | −8.73 | Met118, Ser136 | −8.35 | Met313 |
| 3,5-diOH-BDE-100 | 11.20 | −9.55 | Met118, Ser136 | −8.95 | Met313, Asn331 |
| 6-OH-BDE-47 | 10.99 | −8.25 | Met115 | −8.05 | His435 |
| 6,6′-diOH-BDE-47 | 11.29 | −7.50 | no | −7.74 | no |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Shi, J.; Li, B.; Ge, H.; Tao, H.; Zhang, J.; Li, X.; Cai, Z. Thyroid Hormone Receptor Agonistic and Antagonistic Activity of Newly Synthesized Dihydroxylated Polybrominated Diphenyl Ethers: An In Vitro and In Silico Coactivator Recruitment Study. Toxics 2024, 12, 281. https://doi.org/10.3390/toxics12040281
Zhang M, Shi J, Li B, Ge H, Tao H, Zhang J, Li X, Cai Z. Thyroid Hormone Receptor Agonistic and Antagonistic Activity of Newly Synthesized Dihydroxylated Polybrominated Diphenyl Ethers: An In Vitro and In Silico Coactivator Recruitment Study. Toxics. 2024; 12(4):281. https://doi.org/10.3390/toxics12040281
Chicago/Turabian StyleZhang, Mengtao, Jianghong Shi, Bing Li, Hui Ge, Huanyu Tao, Jiawei Zhang, Xiaoyan Li, and Zongwei Cai. 2024. "Thyroid Hormone Receptor Agonistic and Antagonistic Activity of Newly Synthesized Dihydroxylated Polybrominated Diphenyl Ethers: An In Vitro and In Silico Coactivator Recruitment Study" Toxics 12, no. 4: 281. https://doi.org/10.3390/toxics12040281