

Induction of Oxidative Stress by Waterborne Copper and Arsenic in Larvae of European Seabass (Dicentrarchus labrax L.): A Comparison with Their Effects as Nanoparticles
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Larvae and Experimental Design
2.2. Physical–Chemical Parameters of Water
2.3. Trace Element Concentrations in Water
2.4. Chemical Analysis in Larvae
2.5. Analysis of Biomarkers
2.5.1. Tissue Preparation
2.5.2. Protein Measurement
2.5.3. Lipid Peroxidation (LPO) Assay
2.5.4. CAT Activity Assay
2.5.5. SOD Activity Assay
2.6. Statistical Analysis
3. Results
3.1. Trace Element Concentrations in Water
- –
- The acid–base behaviour of the trace elements in seawater.
- –
- The salinity and the high ionic strength in seawater also provide rapid aggregation [60].
- –
- The adsorption of the trace elements on the suspended matter with subsequent precipitation.
- –
- The dynamic behaviour of nanoparticles (due to their surface properties) such as sedimentation, flotation, or aggregation and which effects depend on the particle’s size.
- –
- The presence of organic colloids in the aqueous environment can overcome the aggregation process of NPs in real water, such as seawater, due to the presence of counter ions in the aquatic environment, especially for NPs of metal oxides [61].
- –
- The interactions of food with exposure to trace elements in the same way as other suspended matter. However, the solid particles may also be uptaken by the organisms.
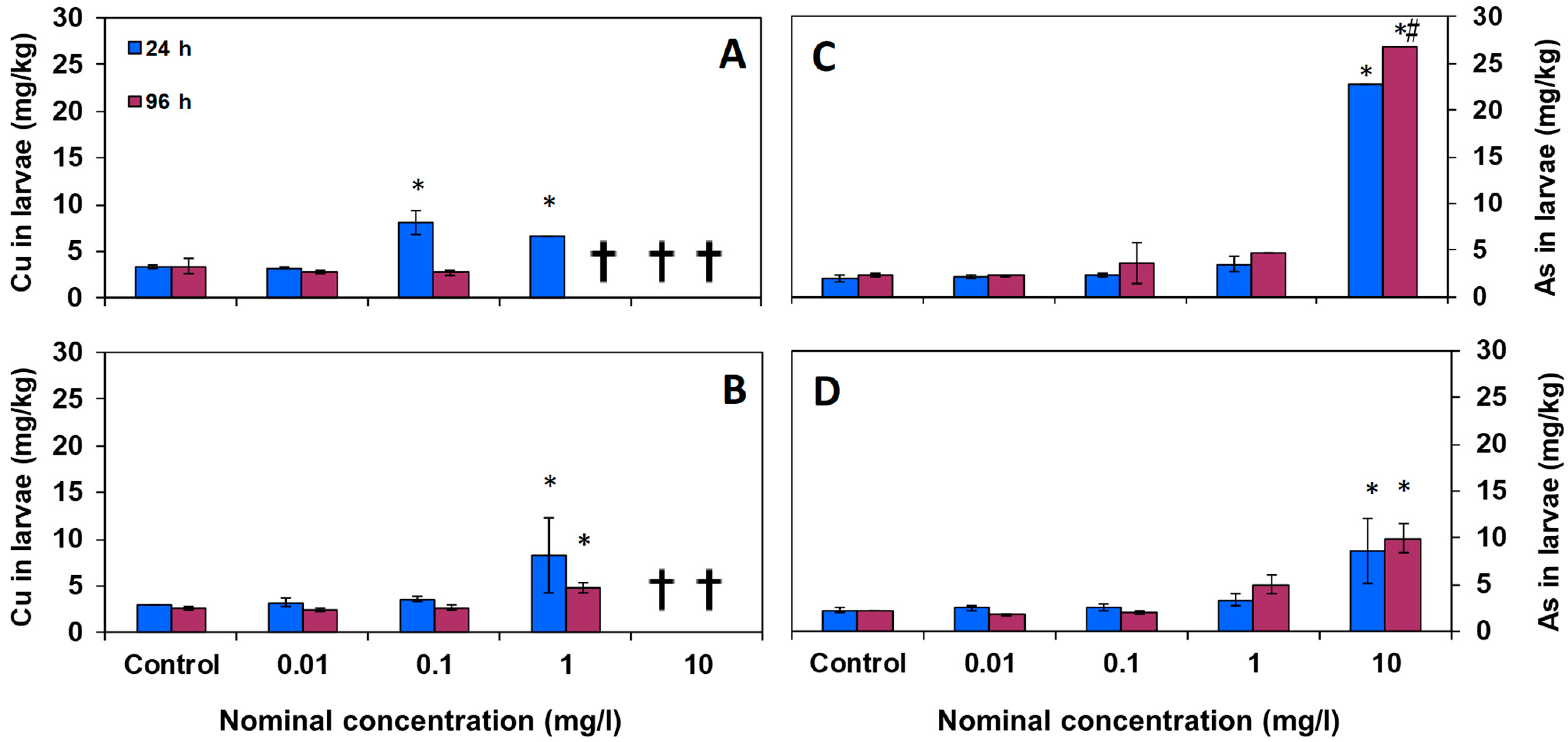
3.2. Chemical Analysis in Larval Tissues
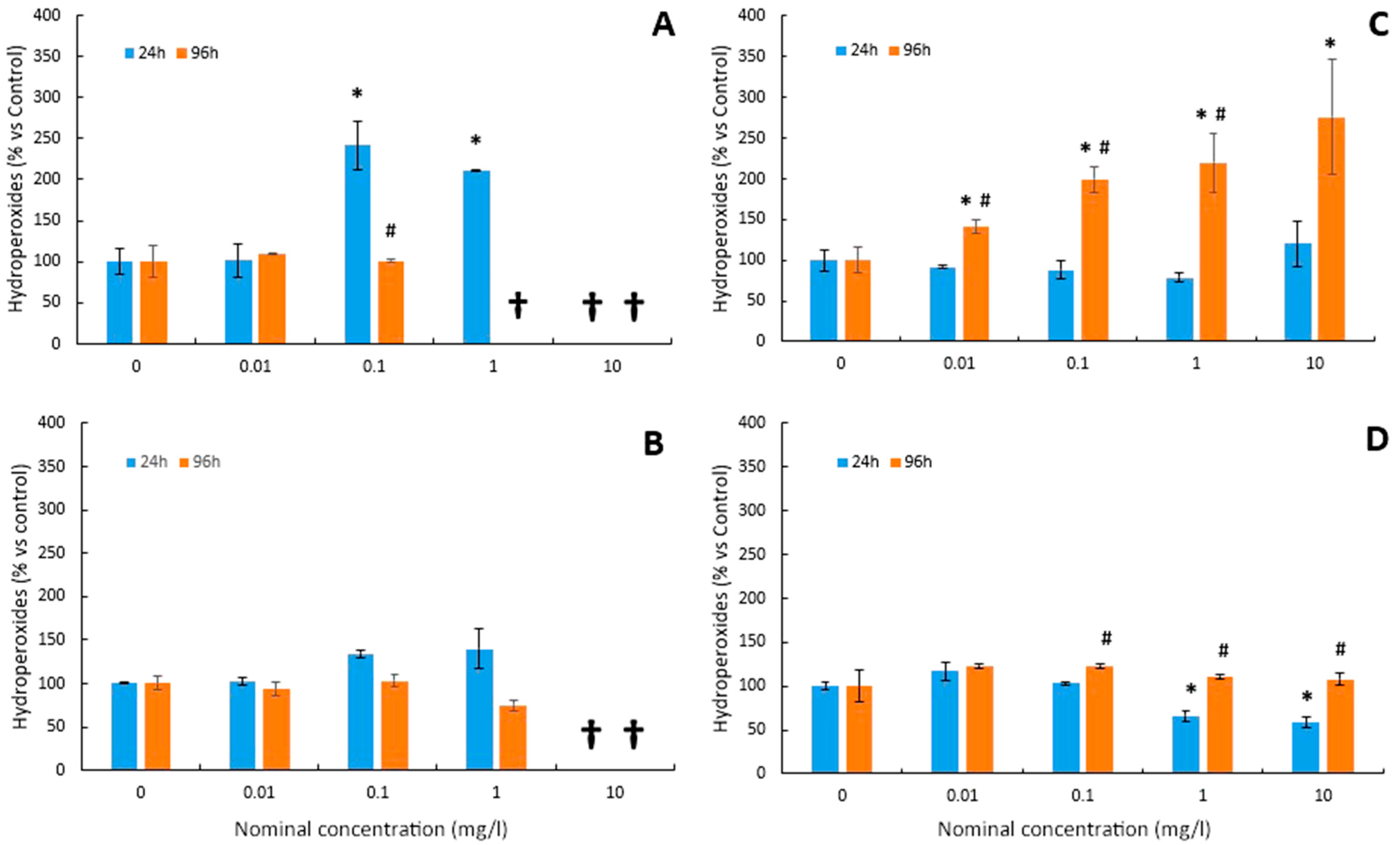
3.3. Lipid Peroxidation
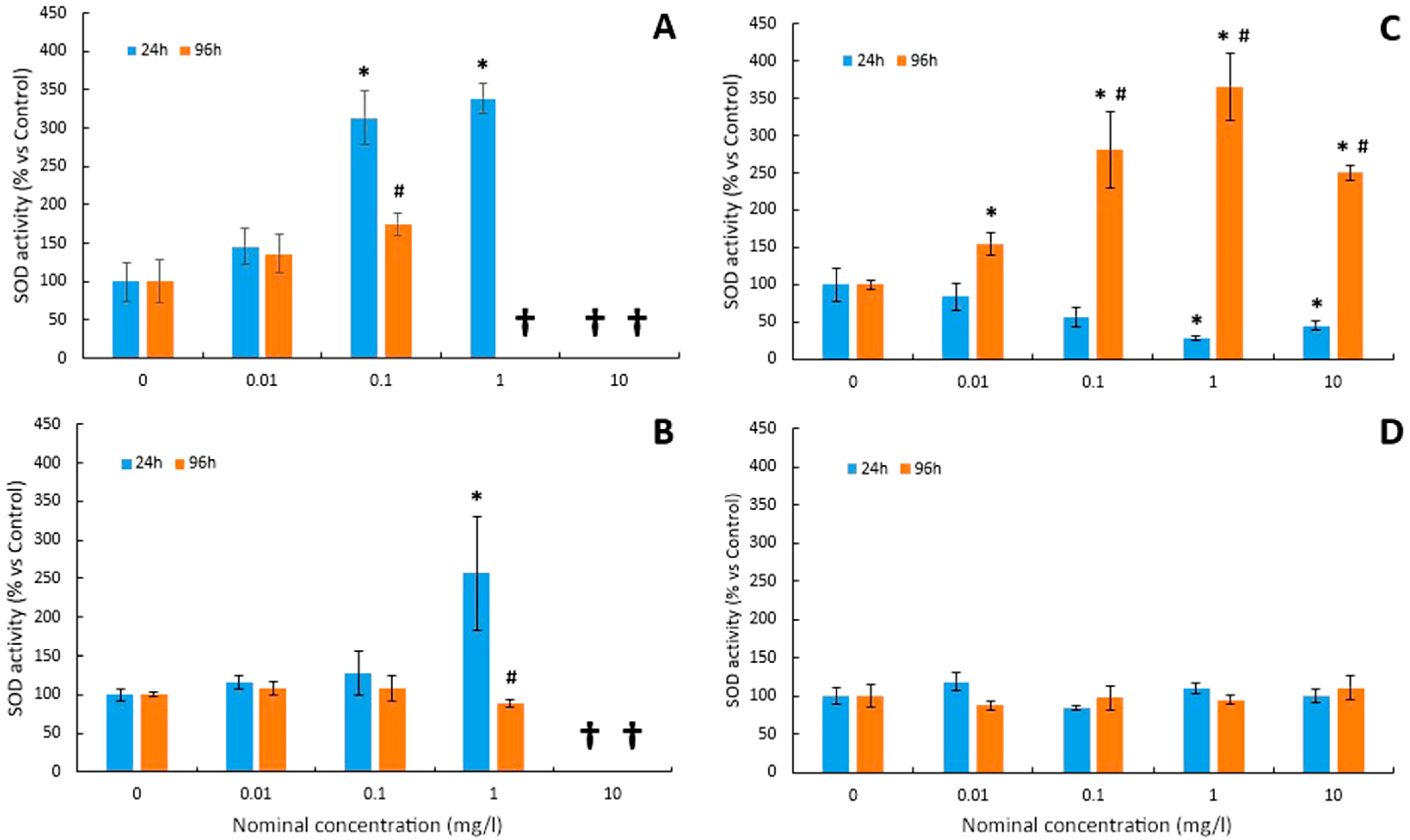
3.4. Superoxide Dismutase Activity
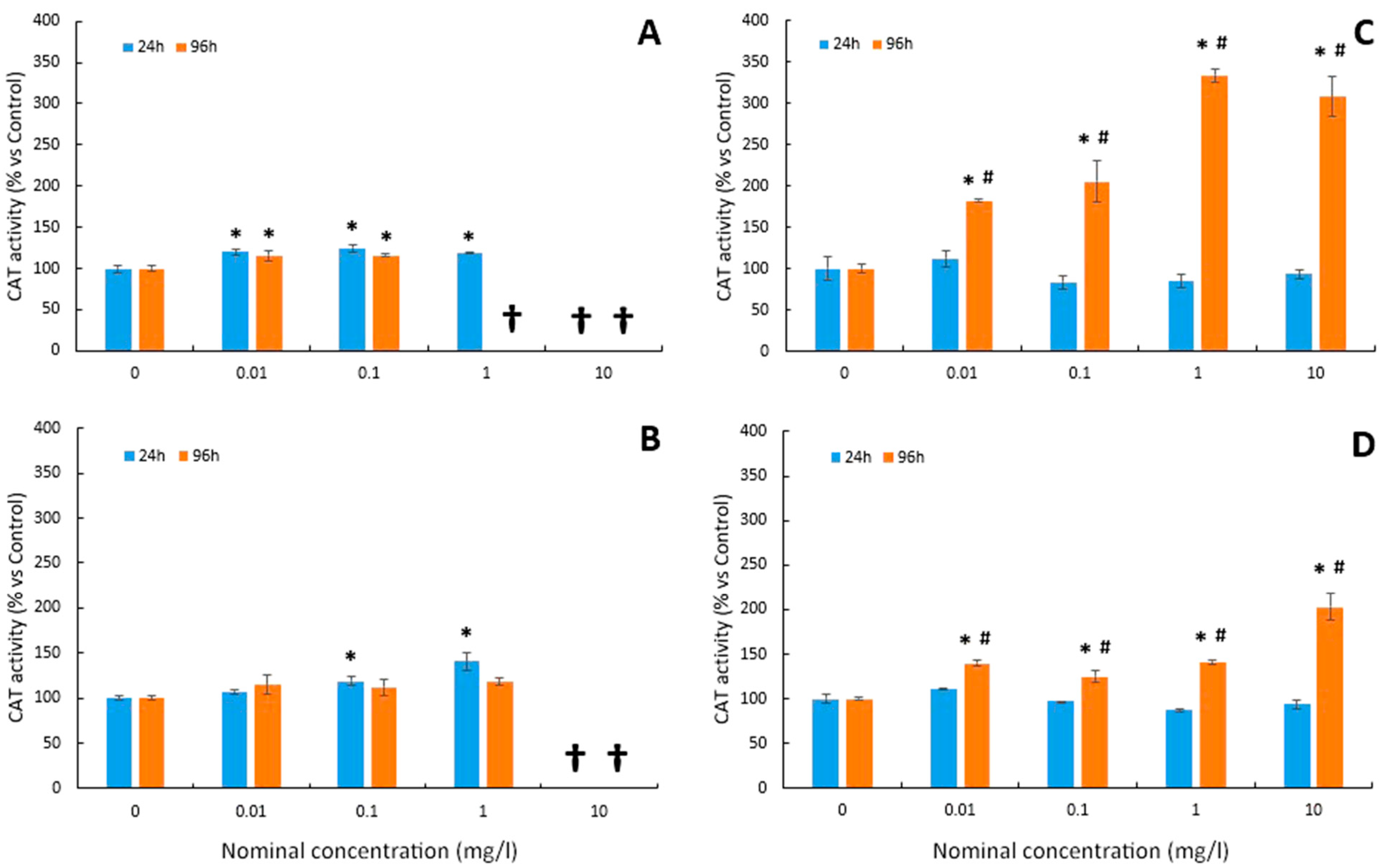
3.5. Catalase Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kadim, M.K.; Risjani, Y. Biomarker for monitoring heavy metal pollution in aquatic environment: An overview toward molecular perspectives. Emerg. Contam. 2022, 8, 195–205. [Google Scholar] [CrossRef]
- Moiseenko, T.I.; Kudryavtseva, L.P. Trace metal accumulation and fish pathologies in areas affected by mining and metallurgical enterprises in the Kola Region, Russia. Environ. Pollut. 2001, 114, 285–297. [Google Scholar] [CrossRef]
- Oliva, M.; Garrido, C.; Sales, D.; González de Canales, M.L. Lindane toxicity on early life stages of gilthead seabream (Sparus aurata) with a note on its histopathological manifestations. Environ. Toxicol. Pharmacol. 2008, 25, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Gashkina, N.A.; Moiseenko, T.I.; Shuma, L.A.; Koroleva, I.M. Biological responses of whitefish (Coregonus lavaretus L.) to reduced toxic impact: Metal accumulation, haematological, immunological, and histopathological alterations. Ecotoxicol. Environ. Saf. 2022, 239, 113659–113671. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Free radicals in biology and medicine. In Free Radicals in Biology and Medicine, 3rd ed.; Halliwell, B., Gutteridge, J.M.C., Eds.; Oxford University Press: Oxford, UK, 1999; pp. 1–25. [Google Scholar]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity, mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Authman, M.M.N.; Zaki, M.S.; Khallaf, E.A.; Abbas, H.H.; Aquac, J. Use of fish as bio-indicator of the effects of heavy metals pollution. Res. Dev. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Mohanty, D.; Samanta, L. Multivariate analysis of potential biomarkers of oxidative stress in Notopterus notopterus tissues from Mahanadi River as a function of concentration of heavy metals. Chemosphere 2016, 155, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.H.; Romma, S.; Garmo, A.; Olsvik, P.A.; Andersen, R.A. Antioxidative stress proteins and their gene expression in brown trout (Salmo trutta) from three rivers with different heavy metal levels. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Hansen, B.H.; Romma, S.; Softeland, L.I.R.; Olsvik, P.A.; Andersen, R.A. Induction and activity of oxidative stress-related proteins during waterborne Cu-exposure in brown trout (Salmo trutta). Chemosphere 2006, 65, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Sevcikova, M.; Modra, H.; Slaninova, A.; Svobodova, Z. Metals as a cause of oxidative stress in fish: A review. Vet. Med. 2011, 56, 537–546. [Google Scholar] [CrossRef]
- WHO International Programme on Chemical Safety (IPCS). Copper. In Environmental Health Criteria 200; World Health Organization: Geneva, Switzerland, 1998. [Google Scholar]
- Hernandez, P.P.; Undurraga, C.; Gallardo, V.E.; Mackenzie, N.; Allende, M.L.; Reyes, A.E. Sublethal concentrations of waterborne copper induce cellular stress and cell death in zebrafish embryos and larvae. Biol. Res. 2011, 44, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Romeo, M.; Bennani, N.; Gnassia-Barelli, M.; Lafaurie, M.; Girard, J.P. Cadmium and copper display different responses towards oxidative stress in the kidney of the seabass Dicentrarchus labrax. Aquat. Toxicol. 2000, 48, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Ates, B.; Orun, I.; Talas, Z.S.; Durmaz, G.; Yilmaz, I. Effects of sodium selenite on some biochemical and hematological parameters of rainbow trout (Oncorhynchus mykiss Walbaum, 1792) exposed to Pb2+ and Cu2+. Fish Physiol. Biochem. 2008, 34, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.A.; Hoff, M.L.; Klein, R.D.; Cardozo, J.G.; Giacomin, M.M.; Pinho, G.L.; Bianchini, A. Biomarkers of waterborne copper exposure in the guppy Poecilia vivipara acclimated to salt water. Aquat. Toxicol. 2013, 138–139, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Leite, C.E.; de Oliveira Maboni, L.; Fernandez Cruz, F.; Rosemberg, D.B.; Zimmermann, F.F.; Pereira, T.C.; Bogo, M.R.; Bonan, C.D.; Campos, M.M.; Morrone, F.B.; et al. Involvement of purinergic system in inflammation and toxicity induced by copper in zebrafish larvae. Toxicol. Appl. Pharmacol. 2013, 272, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.M.; Wood, C.M.; McClelland, G.B. Oxidative stress response and gene expression with acute copper exposure in Zebrafish (Danio rerio). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Atli, G.; Alptekin, Ö.; Tükel, S.; Canli, M. Response of catalase activity to Ag+, Cd2+, Cu2+ and Zn2+ in five tissues of freshwater fish Oreochromis niloticus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 218–224. [Google Scholar] [CrossRef] [PubMed]
- US-EPA (Environmental Protection Agency). National Primary Drinking Water Regulations; Arsenic and Clarifications to Compliance and New Source Contaminants Monitoring. 2001. Available online: https://www.federalregister.gov/documents/2001/01/22/01-1668/national-primary-drinking-water-regulations-arsenic-and-clarifications-to-compliance-and-new-source (accessed on 7 November 2022).
- Wang, Z.; Guo, H.; Liu, H.; Zhang, W. Source, migration, distribution, toxicological effects and remediation technologies of arsenic in groundwater in China. China Geol. 2023, 6, 476–493. [Google Scholar]
- Usese, A.; Chukwu, O.L.; Rahman, M.M.; Naidu, R.; Islam, S.; Oyewo, E.O. Concentrations of arsenic in water and fish in a tropical open lagoon, Southwest-Nigeria: Health risk assessment. Environ. Technol. Innov. 2017, 8, 164–170. [Google Scholar] [CrossRef]
- Das, S.; Unni, B.; Bhattacharjee, M.; Wann, S.B.; Rao, P.G. Toxicological effects of arsenic exposure in a freshwater teleost fish. Channa punctatus. Afr. J. Biotechnol. 2012, 11, 4447–4454. [Google Scholar]
- Sarkar, S.; Mukherjee, S.; Chattopadhyay, A.; Bhattacharya, S. Low dose of arsenic trioxide triggers oxidative stress in zebrafish brain: Expression of antioxidant genes. Ecotoxicol. Environ. Saf. 2014, 107, 1–8. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Lou, B.; Wu, R.; Wang, G.; Lu, C.; Wang, H.; Pi, J.; Xu, Y. The role of reactive oxygen species in arsenic toxicity. Biomolecules 2020, 10, 240. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Bhattacharya, S. Induction of oxidative stress by arsenic in Clarias batrachus: Involvement of peroxisomes. Ecotoxicol. Environ. Saf. 2007, 66, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Díaz-de-Alba, M.; Granado-Castro, M.D.; Galindo-Riaño, M.D.; Casanueva-Marenco, M.J. Comprehensive assessment and potential ecological risk of trace element pollution (As, Ni, Co and Cr) in aquatic environmental samples from an industrialized area. Int. J. Environ. Res. Public Health 2021, 18, 7348. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Chidambaram, S. Assessment of trace inorganic contaminates in water and sediment to address its impact on common fish varieties along Kuwait Bay. Environ. Geochem. Health 2021, 43, 855–883. [Google Scholar] [CrossRef]
- Durán, I.; Nieto, O. Water Characterization in Three Industrialized Harbours (Vigo, Bilbao and Pasajes) in North Coast of Spain. Mar. Pollut. Bull. 2012, 64, 410–415. [Google Scholar] [CrossRef]
- Avvari, L.; Basuri, C.K.; Hari Krishna Chari, N.V.; Tirukkovalluri, S.R.; Gollapalli, N.R. Assessment of Heavy Metal Distribution in Seawater of Kakinada Bay, a Tropical Mangrove-Rich Coastal Environment. Mar. Pollut. Bull. 2022, 181, 113877–113884. [Google Scholar] [CrossRef]
- Sarmiento, A.M.; Nieto, J.M.; Olías, M. The contaminant load transported by the river Odiel to the Gulf of Cádiz (SW Spain). Appl. Earth Sci. 2004, 113, 117–122. [Google Scholar] [CrossRef]
- Nieboer, E.; Richardson, D.H.S. The replacement of the nondescript term ‘heavy metals’ by a biologically and chemically significant classification of metal ions. Environ. Poll. Ser. B Chem. Phys. 1980, 1, 3–26. [Google Scholar] [CrossRef]
- McCully, M.; Sánchez-Navarro, M.; Teixidó, M.; Giralt, E. Peptide Mediated Brain Delivery of Nano- and Submicroparticles: A Synergistic Approach. Curr. Pharm. Des. 2017, 23, 1–11. [Google Scholar] [CrossRef]
- Rastgar, S.; Ardeshir, R.A.; Segner, H.; Tyler, C.R.; Peijnenburg, W.J.G.M.; Wang, Y.J.; Movahedinia, A. Immunotoxic effects of metal-based nanoparticles in fish and bivalves. Nanotoxicology 2022, 16, 88–113. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Malinowski, C.R.; Sepúlveda, M.S. Emerging trends in nanoparticle toxicity and the significance of using Daphnia as a model organism. Chemosphere 2022, 291, 132941–132959. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Xiong, G.; Liu, Z. Toxicity of metal-based nanoparticles: Challenges in the nano era. Front. Bioeng. Biotechnol. 2022, 10, 1001572–1001587. [Google Scholar] [CrossRef] [PubMed]
- Najahi-Missaoui, W.; Arnold, R.D.; Cummings, B.S. Safe Nanoparticles: Are We There Yet? Int. J. Mol. Sci. 2021, 22, 385. [Google Scholar] [CrossRef] [PubMed]
- Windell, D.L.; Mourabit, S.; Moger, J.; Owen, S.F.; Winter, M.J.; Tyler, C.R. The influence of size and surface chemistry on the bioavailability, tissue distribution and toxicity of gold nanoparticles in zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2023, 260, 115019–115030. [Google Scholar] [CrossRef] [PubMed]
- Caixeta, M.B.; Araújo, P.S.; Gonçalves, B.B.; Silva, L.D.; Grano-Maldonado, M.I.; Rocha, T.L. Toxicity of engineered nanomaterials to aquatic and land snails: A scientometric and systematic review. Chemosphere 2020, 260, 127654. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Prasad, S.M.; Bashri, G. Fate and toxicity of nanoparticles in aquatic systems. Acta Geochim. 2023, 42, 63–76. [Google Scholar] [CrossRef]
- Gottschalk, F.; Sun, T.; Nowack, B. Environmental concentrations of engineered nanomaterials: Review of modeling and analytical studies. Environ. Pollut. 2013, 181, 287–300. [Google Scholar] [CrossRef]
- Luo, P.; Roca, A.; Tiede, K.; Privett, K.; Jiang, J.; Pinkstone, J.; Ma, G.; Veinot, J.; Boxall, A. Application of nanoparticle tracking analysis for characterising the fate of engineered nanoparticles in sediment-water systems. J. Environ. Sci. 2018, 64, 62–71. [Google Scholar] [CrossRef]
- Forero-López, A.D.; Fabiani, M.; Lassalle, V.L.; Spetter, C.V.; Severini, M.D.F. Critical review of the characteristics, interactions, and toxicity of micro/nanomaterials pollutants in aquatic environments. Mar. Pollut. Bull. 2022, 174, 113276. [Google Scholar] [CrossRef]
- Yu, Z.; Li, Q.; Wang, J.; Yu, Y.; Wang, Y.; Zhou, Q.; Li, P. Reactive oxygen species-related nanoparticle toxicity in the biomedical field. Nanoscale Res. Lett. 2020, 15, 115–128. [Google Scholar] [CrossRef]
- Ahmad, I.; Maria, V.L.; Pacheco, M.M.A. Juvenile seabass (Dicentrarchus labrax L.) enzymatic and non-enzymatic antioxidant responses following 17β-estradiol exposure. Ecotoxicology 2009, 18, 974–982. [Google Scholar] [CrossRef]
- Fernandes, D.; Bebianno, M.J.; Porte, C. Assessing pollutant exposure in cultured and wild seabass (Dicentrarchus labrax) from the Iberian Peninsula. Ecotoxicology 2009, 18, 1043–1050. [Google Scholar] [CrossRef]
- Maria, V.L.; Ahmad, I.; Oliveira, M.; Serafim, A.; Bebianno, M.J.; Pacheco, M.; Santos, M.A. Wild juvenile Dicentrarchus labrax L. liver antioxidant and damage responses at Aveiro Lagoon. Portugal. Ecotoxicol. Environ. Saf. 2009, 72, 1861–1870. [Google Scholar] [CrossRef]
- Barboza, L.G.A.; Vieira, L.R.; Branco, V.; Figueiredo, N.; Carvalho, F.; Carvalho, C.; Guilhermino, L. Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchus labrax (Linnaeus, 1758). Aquat. Toxicol. 2018, 195, 49–57. [Google Scholar] [CrossRef]
- Malhotra, N.; Ger, T.R.; Uapipatanakul, B.; Huang, J.C.; Chen, K.H.C.; Hsiao, C.D. Review of copper and copper nanoparticle toxicity in fish. Nanomaterials 2020, 10, 1126. [Google Scholar] [CrossRef]
- Byeon, E.; Kang, H.M.; Yoon, C.; Lee, J.S. Toxicity mechanisms of arsenic compounds in aquatic organisms. Aquat. Toxicol. 2021, 237, 105901–105905. [Google Scholar] [CrossRef]
- Naz, S.; Gul, A.; Zia, M. Toxicity of copper oxide nanoparticles: A review study. IET Nanobiotechnol. 2020, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Baláž, P.; Sedlák, J. Arsenic in cancer treatment: Challenges for application of Realgar nanoparticles (A minireview). Toxins 2010, 2, 1568. [Google Scholar] [CrossRef] [PubMed]
- Hai, Y.; Song, P.; Wang, X.; Zhao, L.; Xie, Q.; Li, J.; Li, Y.; Li, H. Realgar transforming solution as a novel arsenic agent with a lower risk of cardiotoxicity. J. Pharmacol. Sci. 2019, 140, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, Z.; Zhang, M.; Yang, X.; Liu, C.; Qiu, G. Mechanism of arsenic release from realgar oxidation in the presence of dissolved oxygen: Effect of reactive oxygen species and light-induced transformation. Geochim. Cosmochim. Acta 2022, 339, 58–69. [Google Scholar] [CrossRef]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.; Diniz, M. Effect of temperature on oxidative stress in fish: Lipid peroxidation and catalase activity in the muscle of juvenile seabass, Dicentrarchus labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Gay, C.; Gebicki, J.M. A critical evaluation of the effect of sorbitol on the ferric-xylenol orange hydroperoxide assay. Anal. Biochem. 2000, 284, 217–220. [Google Scholar] [CrossRef]
- Claiborne, A. Catalase activity. In Handbook of Methods for Oxygen Radical Lethal Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Crouch, R.K.; Gandy, S.E.; Kimsey, G.; Galbraith, R.A.; Galbraith, G.M.; Buse, M.G. The inhibition of islet superoxide dismutase by diabetogenic drugs. Diabetes 1981, 30, 235–241. [Google Scholar] [CrossRef]
- Gondikas, A.; Gallego-Urrea, J.; Halbach, M.; Derrien, N.; Hassellöv, M. Nanomaterial fate in seawater: A rapid sink or intermittent stabilization? Front. Environ. Sci. 2020, 8, 151. [Google Scholar] [CrossRef]
- Tso, C.; Zhung, C.; Shih, Y.; Tseng, Y.; Wu, S.; Doong, R. Stability of metal oxide nanoparticles in aqueous solutions. Water Sci. Technol. 2010, 61, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Wu, H. Reconciling the actual and nominal exposure concentrations of microplastics in aqueous phase: Implications for risk assessment and deviation control. J. Hazard. Mater. 2023, 443, 130246. [Google Scholar] [CrossRef]
- OECD. Test No. 203: Fish, Acute Toxicity Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2019. [Google Scholar] [CrossRef]
- Brad, M.A.; Apte, S.C.; Batley, G.E.; Mark, M. Geochemical factors affecting the solubility of copper in seawater. Environ. Chem. 2021, 18, 1–11. [Google Scholar]
- Yuan, S.; Wang, Y.; Liu, R.; Zheng, W.; Chong, X. Behaviour and distribution of arsenic in seawater and suspended particulate matter in the adjacent area of the Changjiang Estuary during summer and autumn. Ecotoxicol. Environ. Saf. 2021, 227, 112884–112892. [Google Scholar] [CrossRef]
- Durán-Toro, V.M.; Price, R.E.; Maas, M.; Brombach, C.-C.; Pichler, T.; Rezwan, K.; Bühring, S.I. Amorphous arsenic sulfide nanoparticles in a shallow water hydrothermal system. Mar. Chem. 2019, 211, 25–36. [Google Scholar] [CrossRef]
- Hansen, J.A.; Lipton, J.; Welsh, P.G.; Morris, J.; Cacela, D.; Suedkamp, M.J. Relationship between exposure duration, tissue residues, growth and mortality in rainbow trout (Oncorhynchus mykiss) juveniles sub-chronically exposed to copper. Aquat. Toxicol. 2002, 58, 175–188. [Google Scholar] [CrossRef]
- Monteiro, D.A.; Rantin, F.T.; Kalinin, A.L. Inorganic mercury exposure: Toxicological effects, oxidative stress biomarkers and bioaccumulation in the tropical freshwater fish matrinxa. Brycon amazonicus (Spix and Agassiz. 1829). Ecotoxicology 2010, 19, 105–123. [Google Scholar] [CrossRef]
- Sánchez, W.; Palluel, O.; Meunier, L.; Coquery, M.; Porcher, J.M.; Aït-Aïssa, S. Copper-induced oxidative stress in three-spined stickleback: Relationship with hepatic metal levels. Environ. Toxicol. Pharmacol. 2005, 19, 177–183. [Google Scholar] [CrossRef]
- Velma, V.; Tchounwou, P.B. Chromium-induced biochemical, genotoxic and histopathologic effects in liver and kidney of goldfish, Carassius auratus. Mutat. Res. 2010, 698, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.C.; Torronteras, R.; Córdoba, F.; Canalejo, A. Acute toxicity of manganese in goldfish Carassius auratus is associated with oxidative stress and organ specific antioxidant responses. Ecotoxicol. Environ. Saf. 2012, 78, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Handy, R.D.; von der Kammer, F.; Lead, J.R.; Hassellöv, M.; Owen, R.; Crane, M. The ecotoxicology and chemistry of manufactured nanoparticles. Ecotoxicology 2008, 17, 287–314. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Vijver, M.G.; Ahmad, F.; Richardson, M.K.; Peijnenburg, W.J. Toxicity of different-sized copper nano- and submicron particles and their shed copper ions to zebrafish embryos. Environ. Toxicol. Chem. 2014, 33, 1774–1782. [Google Scholar] [CrossRef] [PubMed]
- Shaw, B.J.; Handy, R.D. Physiological effects of nanoparticles on fish: A comparison of nanometals versus metal ions. Environ. Int. 2011, 37, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Foroozandeh, P.; Aziz, A.A. Insight into cellular uptake and intracellular trafficking of nanoparticles. Nanoscale Res. Lett. 2018, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Grosell, M.H.; Hogstrand, C.; Wood, C.M. Renal Cu and Na excretion and hepatic Cu metabolism in both Cu acclimated and non-acclimated rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 1998, 40, 275–291. [Google Scholar] [CrossRef]
- Shaw, B.J.; Al-Bairuty, G.; Handy, R.D. Effects of waterborne copper nanoparticles and copper sulphate on rainbow trout. (Oncorhynchus mykiss): Physiology and accumulation. Aquat. Toxicol. 2012, 116–117, 90–101. [Google Scholar] [CrossRef]
- Hao, L.; Wang, Z.; Xing, B. Effect of sub-acute exposure to TiO2 nanoparticles on oxidative stress and histopathological changes in Juvenile Carp (Cyprinus carpio). J. Environ. Sci. 2009, 21, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Chen, L. Oxidative stress responses in different organs of carp (Cyprinus carpio) with exposure to ZnO nanoparticles. Ecotoxicol. Environ. Saf. 2012, 80, 103–110. [Google Scholar] [CrossRef]
- Ventura-Lima, J.; de Castro, M.R.; Acosta, D.; Fattorini, D.; Regoli, F.; de Carvalho, L.M.; Bohrer, D.; Geracitano, L.A.; Barros, D.M.; Marins, L.F.; et al. Effects of arsenic (As) exposure on the antioxidant status of gills of the zebrafish Danio rerio (Cyprinidae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kang, J.C. The arsenic accumulation and its effect on oxidative stress response in juvenile rockfish, Sebastes schlegelii exposed to waterborne arsenic (As3+). Environ. Toxicol. Pharmacol. 2015, 39, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Altikat, S.; Uysal, K.; Kuru, H.I.; Kavasoglu, M.; Ozturk, G.N.; Kucuk, A. The effect of arsenic on some antioxidant enzyme activities and lipid peroxidation in various tissues of mirror carp (Cyprinus carpio carpio). Environ. Sci. Pollut. Res. Int. 2015, 22, 3212–3218. [Google Scholar] [CrossRef] [PubMed]
- Shankar, S.; Shanker, U. Arsenic contamination of groundwater: A review of sources, prevalence, health risks, and strategies for mitigation. Sci. World J. 2014, 2014, 304524. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | T (°C) | pH | O2 dissolved | Salinity | Suspended Solids | DOC |
|---|---|---|---|---|---|---|
| (mg/L) | (g/L) | (mg/L) | (mg/L) | |||
| Cu | 17.4 ± 1.0 | 8.8 ± 0.4 | 8.0 ± 0.8 | 38.8 ± 0.7 | 29.0 ± 0.5 | 6.8 ± 3.1 |
| Cu-NP | 17.6 ± 0.8 | 8.8 ± 0.2 | 7.9 ± 0.6 | 38.3 ± 1.5 | 28.7 ± 1.0 | 7.9 ± 3.6 |
| As | 17.5 ± 0.6 | 8.3 ± 0.5 | 7.6 ± 0.6 | 38.6 ± 0.7 | 28.9 ± 0.4 | 8.0 ± 2.5 |
| As-NP | 17.6 ± 0.5 | 8.5 ± 0.3 | 7.7 ± 0.7 | 38.4 ± 0.8 | 28.7 ± 0.5 | 6.7 ± 2.4 |
| Element | Reference Material | Method | LOD of Method (µg/L) | Blank Concentration (µg/L) | Found Concentration (µg/L) | Certified Concentration (µg/L) |
|---|---|---|---|---|---|---|
| Cu | LGC 6016 | DPASV | 0.069 | 1.944 | 189.3 ± 3.2 | 190.0 ± 2.0 |
| Cu | LGC 6016 | ICP-AES | 6.300 | <LD | 189.3 ± 3.8 | 190.0 ± 2.0 |
| As | BCR-610 | DPASV | 0.098 | <LD | 9.61 ± 0.92 | 10.8 ± 0.4 |
| As | TMDA-64.2 | ICP-AES | 65.000 | <LD | 158.4 ± 24.0 | 162.0 ± 7.7 |
| Element | LOD of Method (µg/L) | Blank Concentration (µg/L) | Found Concentration (mg/kg) | Certified Concentration (mg/kg) |
|---|---|---|---|---|
| Cu | 0.004 | 0.023 | 31.7 ± 0.6 | 31.2 ± 1 |
| As | 0.293 | <LD | 10.6 ± 0.1 | 10.2 ± 0.5 |
| Nominal Concentration | Copper Test | Exposure Time | Arsenic Test | Exposure Time | ||||
|---|---|---|---|---|---|---|---|---|
| 0 h | 24 h | 96 h | 0 h | 24 h | 96 h | |||
| Control | Cu | 0.001 ± 0.001 | 0.001 ± 0.002 | 0.002 ± 0.001 | As | 0.002 ± 0.001 | 0.002 ± 0.001 | 0.004 ± 0.003 |
| Cu-NP | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.002 ± 0.002 | As-NP | 0.002 ± 0.000 | 0.002 ± 0.000 | 0.002 ± 0.000 | |
| 0.01 | Cu | 0.008 ± 0.001 (−24.7%) | 0.006 ± 0.000 (−36.2%) | 0.004 ± 0.001 (−59.6%) | As | 0.009 ± 0.001 (−13.5%) | 0.009 ± 0.000 (−13.5%) | 0.009 ± 0.001 (−14.5%) |
| Cu-NP | 0.005 ± 0.001 (−51.8%) | 0.003 ± 0.000 (−67.5%) | 0.002 ± 0.001 (−80.2%) | As-NP | 0.007 ± 0.001 (−25.4%) | 0.008 ± 0.001 (−25%) | 0.007 ± 0.001 (−26%) | |
| 0.1 | Cu | 0.131 ± 0.009 (31.3%) | 0.117 ± 0.003 (17%) | 0.080 ± 0.000 (−20%) | As | 0.058 ± 0.037 (−42.4%) | 0.078 ± 0.003 (−22.4%) | 0.110 ± 0.021 (10.5%) |
| Cu-NP | 0.106 ± 0.008 (5.5%) | 0.081 ± 0.006 (−19.5%) | 0.068 ± 0.004 (−31.7%) | As-NP | 0.084 ± 0.008 (−16.2%) | 0.077 ± 0.020 (−23.3%) | 0.082 ± 0.006 (−17.9%) | |
| 1 | Cu | 0.905 ± 0.070 (−9.5%) | 0.643 ± 0.000 (−35.7%) | † | As | 0.865 ± 0.017 (−13.5%) | 0.730 ± 0.047 (−27%) | 0.595 ± 0.023 (−40.6%) |
| Cu-NP | 0.712 ± 0.041 (−28.9%) | 0.335 ± 0.065 (−66.5%) | 0.222 ± 0.045 (−77.8%) | As-NP | 0.769 ± 0.020 (−23.1%) | 0.563 ± 0.096 (−43.8%) | 0.733 ± 0.010 (−26.8%) | |
| 10 | Cu | 8.513 ± 1.213 (−14.9%) | † | † | As | 5.640 ± 1.006 (−43.6%) | 6.415 ± 1.902 (−35.9%) | 4.681 ± 0.283 (−53.2%) |
| Cu-NP | 7.910 ± 0.561 (−20.9%) | † | † | As-NP | 7.628 ± 0.276 (−23.7%) | 6.232 ± 1.844 (−37.7%) | 6.911 ± 0.691 (−30.9%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torronteras, R.; Díaz-de-Alba, M.; Granado-Castro, M.D.; Espada-Bellido, E.; Córdoba García, F.; Canalejo, A.; Galindo-Riaño, M.D. Induction of Oxidative Stress by Waterborne Copper and Arsenic in Larvae of European Seabass (Dicentrarchus labrax L.): A Comparison with Their Effects as Nanoparticles. Toxics 2024, 12, 141. https://doi.org/10.3390/toxics12020141
Torronteras R, Díaz-de-Alba M, Granado-Castro MD, Espada-Bellido E, Córdoba García F, Canalejo A, Galindo-Riaño MD. Induction of Oxidative Stress by Waterborne Copper and Arsenic in Larvae of European Seabass (Dicentrarchus labrax L.): A Comparison with Their Effects as Nanoparticles. Toxics. 2024; 12(2):141. https://doi.org/10.3390/toxics12020141
Chicago/Turabian StyleTorronteras, Rafael, Margarita Díaz-de-Alba, María Dolores Granado-Castro, Estrella Espada-Bellido, Francisco Córdoba García, Antonio Canalejo, and María Dolores Galindo-Riaño. 2024. "Induction of Oxidative Stress by Waterborne Copper and Arsenic in Larvae of European Seabass (Dicentrarchus labrax L.): A Comparison with Their Effects as Nanoparticles" Toxics 12, no. 2: 141. https://doi.org/10.3390/toxics12020141