
Subchronic Arsenite Exposure Induced Atrophy and Erythropoietin Sensitivity Reduction in Skeletal Muscle Were Relevant to Declined Serum Melatonin Levels in Middle-Aged Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design of the Animal Study
2.3. Determination of Total Arsenic
2.4. Enzyme-Linked Immunosorbent Assay (ELISA) Assays
2.5. Histology Assessment
2.6. Immunofluorescence Assay
2.7. Genomic DNA Isolation and Telomere Length Analysis
2.8. Western Blot
2.9. Statistical Analysis
3. Results
3.1. Subchronic Arsenite Exposure Reduced Skeletal Muscle Index in Middle-Aged Rats
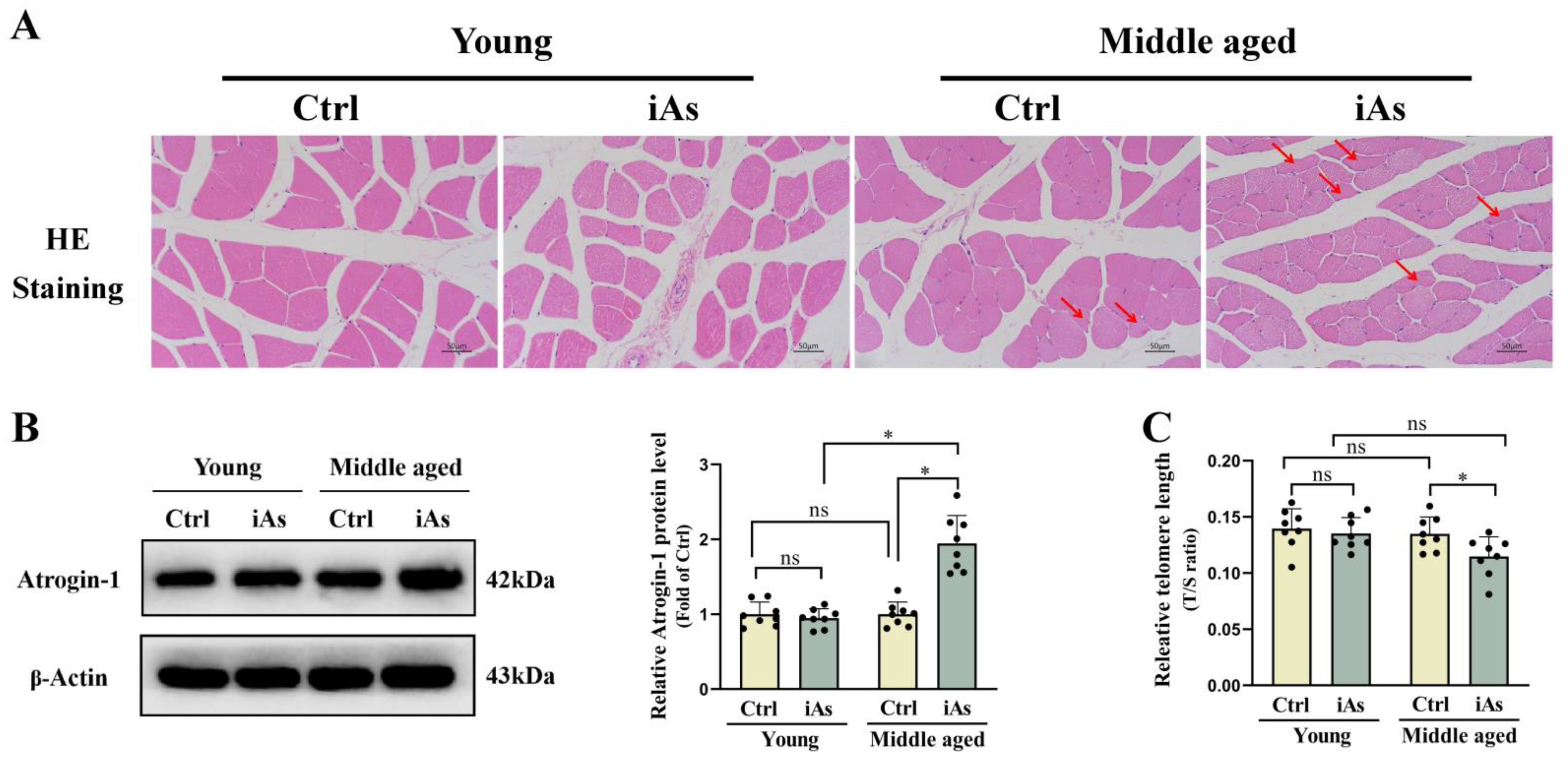
3.2. Arsenite Exposure Accelerated Skeletal Muscle Atrophy and Telomere Shortening in Middle-Aged Rats
3.3. Arsenite Exposure Induced Type II to Type I Myofiber Switching in Middle-Aged Rats
3.4. Influence of Arsenite Exposure on Serum Levels of MT and EPO in Rats
3.5. Influence of Arsenite Exposure on EPOR Expression of Skeletal Muscle in Rats
3.6. Effect of Exogenous MT on Arsenite-Induced Skeletal Muscle Injury in Middle-Aged Rats
4. Discussion
4.1. Arsenic Exposure and Skeletal Muscle Injury
4.2. MT in Arsenite-Induced Skeletal Muscle Injury
4.3. EPO Sensitivity in Arsenite-Induced Skeletal Muscle Injury
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, Q.Y.; Costa, M. Arsenic: A Global Environmental Challenge. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Rastegari Mehr, M.; Keshavarzi, B.; Moore, F.; Hooda, P.S.; Busquets, R.; Ghorbani, Z. Arsenic in the rock-soil-plant system and related health risk in a magmatic-metamorphic belt, West of Iran. Environ. Geochem. Health 2020, 42, 3659–3673. [Google Scholar] [CrossRef] [PubMed]
- Abdul, K.S.; Jayasinghe, S.S.; Chandana, E.P.; Jayasumana, C.; De Silva, P.M. Arsenic and human health effects: A review. Environ. Toxicol. Pharmacol. 2015, 40, 828–846. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, M.S.; Rahman, M.M.; Mise, N.; Sikder, M.T.; Ichihara, G.; Uddin, M.K.; Kurasaki, M.; Ichihara, S. Environmental arsenic exposure and its contribution to human diseases, toxicity mechanism and management. Environ. Pollut. 2021, 289, 117940. [Google Scholar] [CrossRef]
- Yang, L.; Chai, Y.; Yu, J.; Wei, B.; Xia, Y.; Wu, K.; Gao, J.; Guo, Z.; Cui, N. Associations of arsenic metabolites, methylation capacity, and skin lesions caused by chronic exposure to high arsenic in tube well water. Environ. Toxicol. 2017, 32, 28–36. [Google Scholar] [CrossRef]
- Mondal, V.; Hosen, Z.; Hossen, F.; Siddique, A.E.; Tony, S.R.; Islam, Z.; Islam, M.S.; Hossain, S.; Islam, K.; Sarker, M.K.; et al. Arsenic exposure-related hyperglycemia is linked to insulin resistance with concomitant reduction of skeletal muscle mass. Environ. Int. 2020, 143, 105890. [Google Scholar] [CrossRef]
- Sarker, M.K.; Tony, S.R.; Siddique, A.E.; Karim, M.R.; Haque, N.; Islam, Z.; Islam, M.S.; Khatun, M.; Islam, J.; Hossain, S.; et al. Arsenic Secondary Methylation Capacity Is Inversely Associated with Arsenic Exposure-Related Muscle Mass Reduction. Int. J. Environ. Res. Public Health 2021, 18, 9730. [Google Scholar] [CrossRef]
- Chen, C.M.; Chung, M.N.; Chiu, C.Y.; Liu, S.H.; Lan, K.C. Inorganic Arsenic Exposure Decreases Muscle Mass and Enhances Denervation-Induced Muscle Atrophy in Mice. Molecules 2020, 25, 3057. [Google Scholar] [CrossRef]
- Yen, Y.P.; Tsai, K.S.; Chen, Y.W.; Huang, C.F.; Yang, R.S.; Liu, S.H. Arsenic inhibits myogenic differentiation and muscle regeneration. Environ. Health Perspect. 2010, 118, 949–956. [Google Scholar] [CrossRef]
- Chiu, C.Y.; Chung, M.N.; Lan, K.C.; Yang, R.S.; Liu, S.H. Exposure of low-concentration arsenic induces myotube atrophy by inhibiting an Akt signaling pathway. Toxicol. Vitro 2020, 65, 104829. [Google Scholar] [CrossRef]
- Merz, K.E.; Thurmond, D.C. Role of Skeletal Muscle in Insulin Resistance and Glucose Uptake. Compr. Physiol. 2020, 10, 785–809. [Google Scholar] [PubMed]
- Westerblad, H.; Bruton, J.D.; Katz, A. Skeletal muscle: Energy metabolism, fiber types, fatigue and adaptability. Exp. Cell Res. 2010, 316, 3093–3099. [Google Scholar] [CrossRef] [PubMed]
- Angulo, J.; El Assar, M.; Rodríguez-Mañas, L. Frailty and sarcopenia as the basis for the phenotypic manifestation of chronic diseases in older adults. Mol. Asp. Med. 2016, 50, 1–32. [Google Scholar] [CrossRef]
- Nilwik, R.; Snijders, T.; Leenders, M.; Groen, B.B.; van Kranenburg, J.; Verdijk, L.B.; van Loon, L.J. The decline in skeletal muscle mass with aging is mainly attributed to a reduction in type II muscle fiber size. Exp. Gerontol. 2013, 48, 492–498. [Google Scholar] [CrossRef] [PubMed]
- Araki, H.; Hino, S.; Anan, K.; Kuribayashi, K.; Etoh, K.; Seko, D.; Takase, R.; Kohrogi, K.; Hino, Y.; Ono, Y.; et al. LSD1 defines the fiber type-selective responsiveness to environmental stress in skeletal muscle. Elife. 2023, 12, e84618. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Connor, K.M.; Aderman, C.M.; Willett, K.L.; Aspegren, O.P.; Smith, L.E. Suppression of retinal neovascularization by erythropoietin siRNA in a mouse model of proliferative retinopathy. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1329–1335. [Google Scholar] [CrossRef]
- Suresh, S.; Rajvanshi, P.K.; Noguchi, C.T. The Many Facets of Erythropoietin Physiologic and Metabolic Response. Front. Physiol. 2020, 10, 1534. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.H.; Lu, I.C.; Tai, M.H.; Chai, C.Y.; Kwan, A.L.; Huang, S.H. Erythropoietin Alleviates Burn-induced Muscle Wasting. Int. J. Med. Sci. 2020, 17, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Suzuki, N.; Yamamoto, M.; Gassmann, M.; Noguchi, C.T. Endogenous erythropoietin signaling facilitates skeletal muscle repair and recovery following pharmacologically induced damage. FASEB J. 2012, 26, 2847–2858. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Bello, V.E.; Sanchis-Gomar, F.; Romagnoli, M.; Derbre, F.; Gomez-Cabrera, M.C.; Viña, J. Three weeks of erythropoietin treatment hampers skeletal muscle mitochondrial biogenesis in rats. J. Physiol. Biochem. 2012, 68, 593–601. [Google Scholar] [CrossRef]
- Lombardero, M.; Kovacs, K.; Scheithauer, B.W. Erythropoietin: A hormone with multiple functions. Pathobiology 2011, 78, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Nijholt, K.T.; Meems, L.M.G.; Ruifrok, W.P.T.; Maass, A.H.; Yurista, S.R.; Pavez-Giani, M.G.; Mahmoud, B.; Wolters, A.H.G.; van Veldhuisen, D.J.; van Gilst, W.H.; et al. The erythropoietin receptor expressed in skeletal muscle is essential for mitochondrial biogenesis and physiological exercise. Pflug. Arch. 2021, 473, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Kiriyama, N.; Ogawa, K.; Inoue, R.; Haque, M.A.; Nakagawa, H. Effect of pentavalent inorganic arsenic salt on erythropoietin production and autophagy induction. Arch. Biochem. Biophys. 2023, 734, 109487. [Google Scholar] [PubMed]
- Medina, S.; Zhang, H.; Santos-Medina, L.V.; Yee, Z.A.; Martin, K.J.; Wan, G.; Bolt, A.M.; Zhou, X.; Stýblo, M.; Liu, K.J. Arsenite Methyltransferase Is an Important Mediator of Hematotoxicity Induced by Arsenic in Drinking Water. Water 2023, 15, 448. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin and healthy aging. Vitam. Horm. 2021, 115, 67–88. [Google Scholar]
- Jin, H.; Xie, W.; Hu, P.; Tang, K.; Wang, X.; Wu, Y.; He, M.; Yu, D.; Li, Y. The role of melatonin in sarcopenia: Advances and application prospects. Exp. Gerontol. 2021, 149, 111319. [Google Scholar] [CrossRef]
- Coto-Montes, A.; Boga, J.A.; Tan, D.X.; Reiter, R.J. Melatonin as a Potential Agent in the Treatment of Sarcopenia. Int. J. Mol. Sci. 2016, 17, 1771. [Google Scholar]
- Hameed, E.N.; Hadi Al Tukmagi, H.F.; Allami, H.C.A. Melatonin Improves Erythropoietin Hyporesponsiveness via Suppression of Inflammation. Rev. Recent Clin. Trials 2019, 14, 203–208. [Google Scholar] [CrossRef]
- Herrera, J.; Nava, M.; Romero, F.; Rodríguez-Iturbe, B. Melatonin Prevents Oxidative Stress Resulting from Iron and Erythropoietin Administration. Am. J. Kidney Dis. 2001, 37, 750–757. [Google Scholar] [CrossRef]
- Nordstrom, D.K. Public health. Worldwide occurrences of arsenic in ground water. Science 2002, 296, 2143–2145. [Google Scholar] [CrossRef]
- Guo, X.; Fu, X.; Liu, X.; Wang, J.; Li, Z.; Gao, L.; Li, Y.; Zhang, W. Role of Pigment Epithelium-Derived Factor in Arsenic-Induced Vascular Endothelial Dysfunction in a Rat Model. Biol. Trace Elem. Res. 2019, 190, 405–413. [Google Scholar]
- Waghe, P.; Sarkar, S.N.; Sarath, T.S.; Kandasamy, K.; Choudhury, S.; Gupta, P.; Harikumar, S.; Mishra, S.K. Subchronic exposure to arsenic through drinking water alters expression of cancer-related genes in rat liver. Biol. Trace Elem. Res. 2017, 176, 350–354. [Google Scholar] [CrossRef]
- Chi, L.; Tu, P.; Liu, C.W.; Lai, Y.; Xue, J.; Ru, H.; Lu, K. Chronic Arsenic Exposure Induces Oxidative Stress and Perturbs Serum Lysolipids and Fecal Unsaturated Fatty Acid Metabolism. Chem. Res. Toxicol. 2019, 32, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Baeeri, M.; Didari, T.; Khalid, M.; Mohammadi-Nejad, S.; Daghighi, S.M.; Farhadi, R.; Rahimifard, M.; Bayrami, Z.; Haghi-Aminjan, H.; Foroumadi, R.; et al. Molecular Evidence of the Inhibitory Potential of Melatonin against NaAsO2-Induced Aging in Male Rats. Molecules 2021, 26, 6603. [Google Scholar] [CrossRef] [PubMed]
- Abdollahzade, N.; Mihanfar, A.; Majidinia, M. Molecular mechanisms underlying ameliorative impact of melatonin against age-dependent chronic arsenic toxicity in rats’ brains. J. Exp. Zool. A Ecol. Integr. Physiol. 2022, 337, 1010–1024. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef]
- Wang, V.C.; Neese, S.L.; Korol, D.L.; Schantz, S.L. Estradiol impairs response inhibition in young and middle-aged, but not old rats. Neurotoxicol. Teratol. 2011, 33, 405–414. [Google Scholar]
- Babur, E.; Tufan, E.; Barutçu, Ö.; Aslan-Gülpınar, E.; Tan, B.; Süer, C.; Dursun, N. Neurodegeneration-related genes are differentially expressed in middle-aged rats compared to young-adult rats having equal performance on long-term memory and synaptic plasticity. Brain Res. Bull. 2022, 182, 90–101. [Google Scholar] [CrossRef]
- Schiaffino, S.; Dyar, K.A.; Ciciliot, S.; Blaauw, B.; Sandri, M. Mechanisms regulating skeletal muscle growth and atrophy. FEBS J. 2013, 280, 4294–4314. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, J.H.; Lee, D.C. Urine melatonin levels are inversely associated with sarcopenia in postmenopausal women. Menopause 2014, 21, 39–44. [Google Scholar] [CrossRef]
- Abdollahzade, N.; Majidinia, M.; Babri, S. Melatonin: A pleiotropic hormone as a novel potent therapeutic candidate in arsenic toxicity. Mol. Biol. Rep. 2021, 48, 6603–6618. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Chen, W.; Wang, D.; Ma, L.; Tao, J.; Zhang, A. Subchronic Arsenite Exposure Induced Atrophy and Erythropoietin Sensitivity Reduction in Skeletal Muscle Were Relevant to Declined Serum Melatonin Levels in Middle-Aged Rats. Toxics 2023, 11, 689. https://doi.org/10.3390/toxics11080689
Chen X, Chen W, Wang D, Ma L, Tao J, Zhang A. Subchronic Arsenite Exposure Induced Atrophy and Erythropoietin Sensitivity Reduction in Skeletal Muscle Were Relevant to Declined Serum Melatonin Levels in Middle-Aged Rats. Toxics. 2023; 11(8):689. https://doi.org/10.3390/toxics11080689
Chicago/Turabian StyleChen, Xiong, Wanying Chen, Dapeng Wang, Lu Ma, Junyan Tao, and Aihua Zhang. 2023. "Subchronic Arsenite Exposure Induced Atrophy and Erythropoietin Sensitivity Reduction in Skeletal Muscle Were Relevant to Declined Serum Melatonin Levels in Middle-Aged Rats" Toxics 11, no. 8: 689. https://doi.org/10.3390/toxics11080689