
Combined Effects of Micro- and Nanoplastics at the Predicted Environmental Concentration on Functional State of Intestinal Barrier in Caenorhabditis elegans
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nanopolystyrene Characterizations
2.2. Animal Maintenance
2.3. Exposure and Assessment Endpoints
2.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.5. Statistical Analyses
3. Results
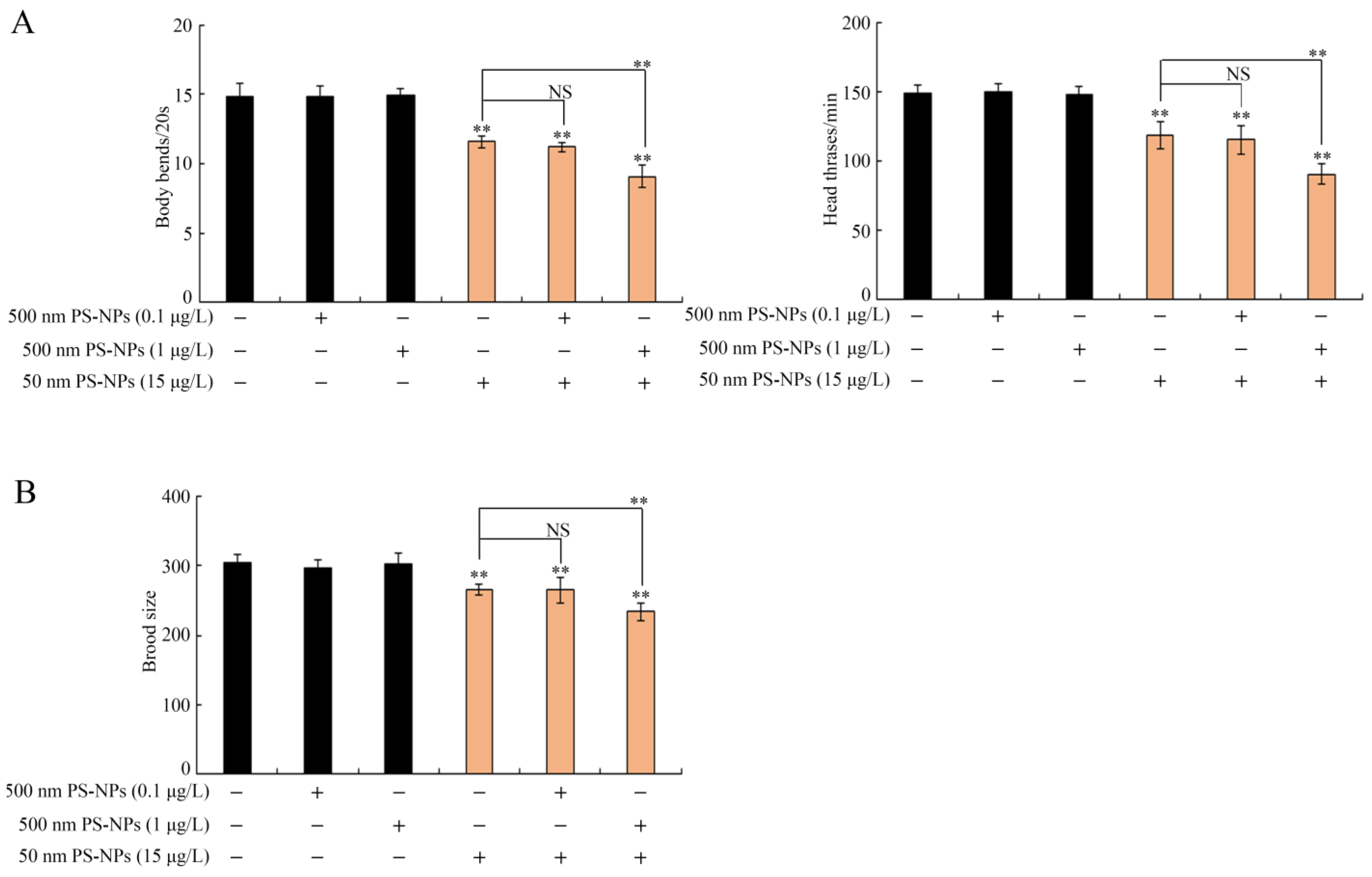
3.1. Combined Effects of PS-50 and PS-500 on Locomotion Behavior and Brood Size
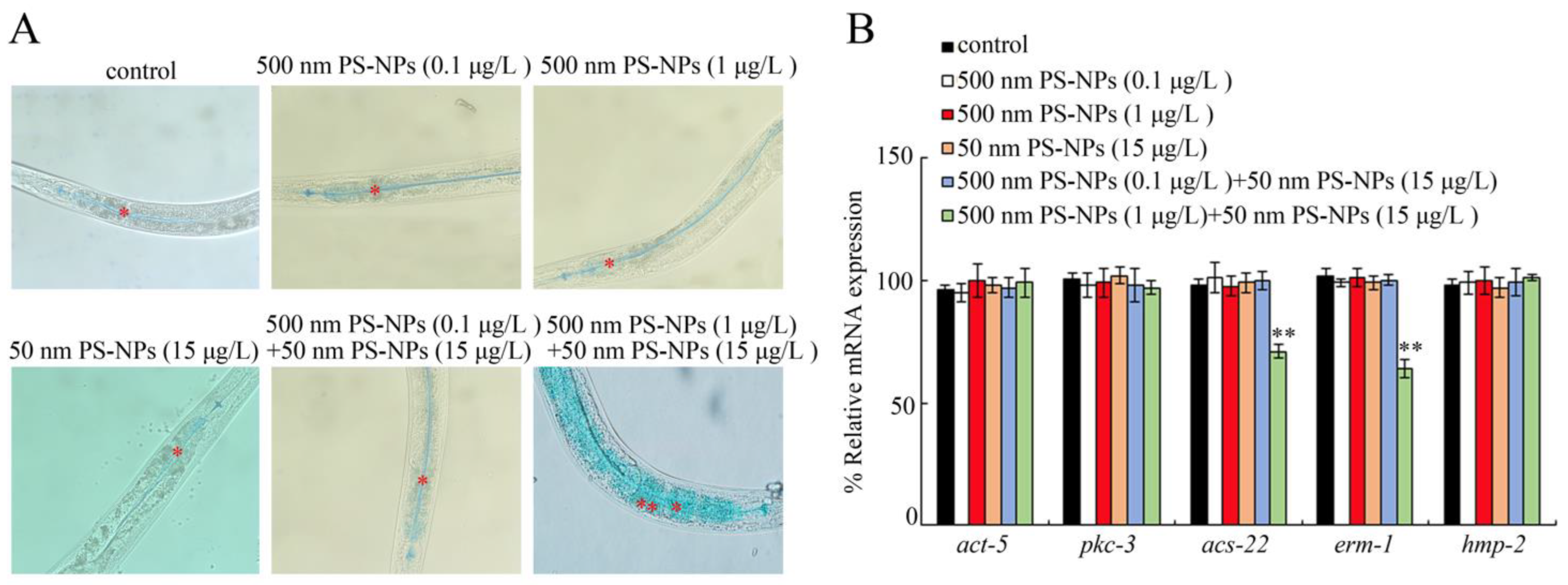
3.2. Combined Effects of PS-50 and PS-500 on Intestinal Morphology
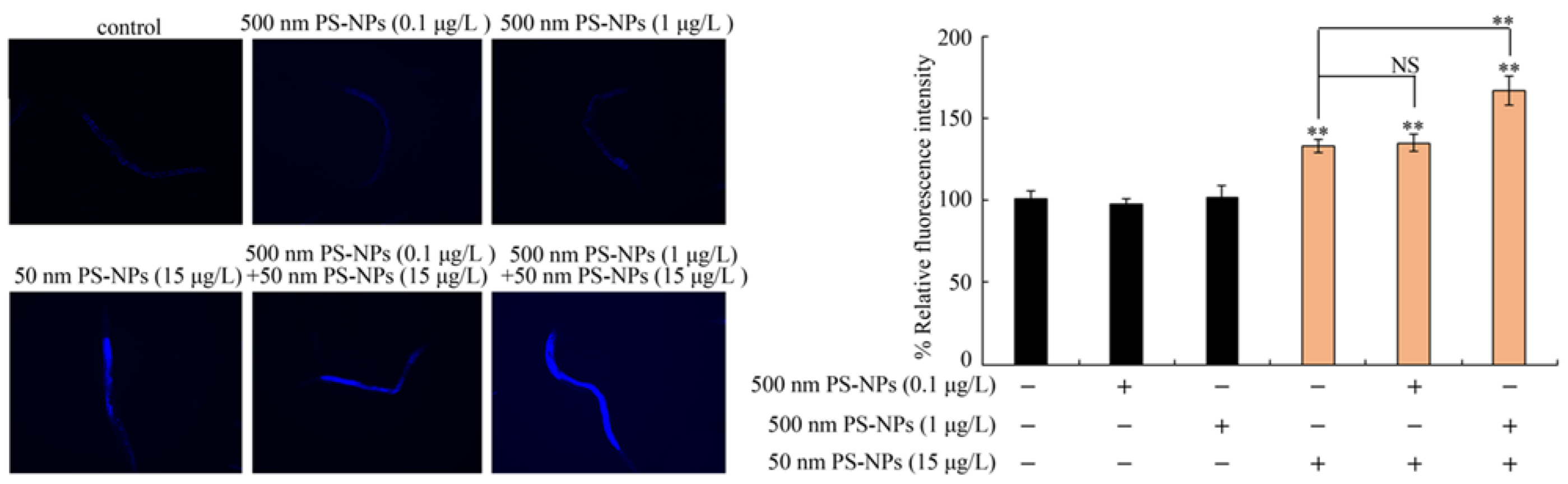
3.3. Combined Effects of PS-50 and PS-500 on Intestinal Permeability
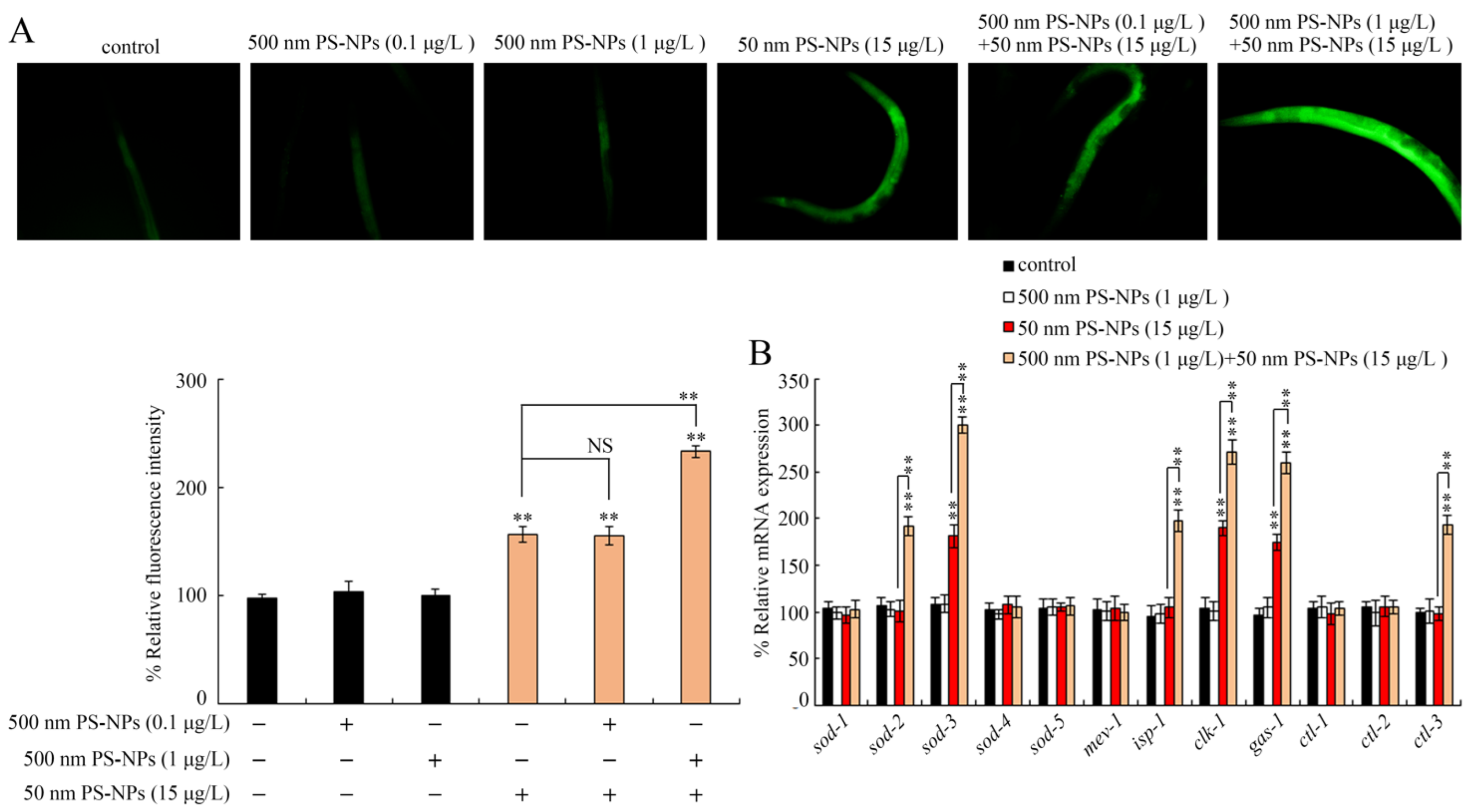
3.4. Combined Effects of PS-50 and PS-500 on Intestinal Autofluorescence
3.5. Combined Effects of PS-50 and PS-500 on Induction of Oxidative Stress
3.6. Combined Effects of PS-50 and PS-500 on Molecular Basis of Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, Y.; Ashokkumar, V.; Amobonye, A.; Bhattacharjee, G.; Sirohi, R.; Singh, V.; Flora, G.; Kumar, V.; Pillai, S.; Zhang, Z.; et al. Current research trends on cosmetic microplastic pollution and its impacts on the ecosystem: A review. Environ. Pollut. 2023, 320, 121106. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Coley, I.; Mendez-Cuadro, D.; Rodriguez-Cavallo, E.; de la Rosa, J.; Olivero-Verbel, J. Trace elements in microplastics in Cartagena: A hotspot for plastic pollution at the Caribbean. Mar. Pollut. Bull. 2019, 139, 402–411. [Google Scholar] [CrossRef]
- Jambeck, J.R.; Geyer, R.; Wilcox, C.; Siegler, T.R.; Perryman, M.; Andrady, A.; Narayan, R.; Law, K.L. Marine pollution. Plastic waste inputs from land into the ocean. Science 2015, 347, 768–771. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Y.; Zhang, L.; Xu, M.; Gao, L.; Zhao, B. The interaction of micro/nano plastics and the environment: Effects of ecological corona on the toxicity to aquatic organisms. Ecotoxicol. Environ. Saf. 2022, 243, 113997. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-W.; Doshi, N.; Mitragotri, S. Adaptive micro and nanoparticles: Temporal control over carrier properties to facilitate drug delivery. Adv. Drug Deliv. Rev. 2011, 63, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tian, L.; Wang, S.; Wang, D. Size-dependent transgenerational toxicity induced by nanoplastics in nematode Caenorhabditis elegans. Sci. Total Environ. 2021, 790, 148217. [Google Scholar] [CrossRef] [PubMed]
- Walczak, A.P.; Kramer, E.; Hendriksen, P.J.M.; Tromp, P.; Helsper, J.P.F.G.; van der Zande, M.; Rietjens, I.M.C.M.; Bouwmeester, H. Translocation of differently sized and charged polystyrene nanoparticles in in vitro intestinal cell models of increasing complexity. Nanotoxicology 2015, 9, 453–461. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H. Uptake and Accumulation of Polystyrene Microplastics in Zebrafish (Danio rerio) and Toxic Effects in Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Zitouni, N.; Bousserrhine, N.; Missawi, O.; Boughattas, I.; Chèvre, N.; Santos, R.; Belbekhouche, S.; Alphonse, V.; Tisserand, F.; Balmassiere, L.; et al. Uptake, tissue distribution and toxicological effects of environmental microplastics in early juvenile fish Dicentrarchus labrax. J. Hazard. Mater. 2021, 403, 124055. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Shen, L.; Ye, X.; Bai, G.; Liao, C.; Chen, Z.; Peng, T.; Li, X.; Kang, X.; An, G. Prenatal and postnatal exposure to polystyrene microplastics induces testis developmental disorder and affects male fertility in mice. J. Hazard. Mater. 2023, 445, 130544. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Wang, D. Toxicity comparison between pristine and sulfonate modified nanopolystyrene particles in affecting locomotion behavior, sensory perception, and neuronal development in Caenorhabditis elegans. Sci. Total Environ. 2020, 703, 134817. [Google Scholar] [CrossRef]
- Qu, M.; Qiu, Y.; Kong, Y.; Wang, D. Amino modification enhances reproductive toxicity of nanopolystyrene on gonad development and reproductive capacity in nematode Caenorhabditis elegans. Environ. Pollut. 2019, 254 Pt A, 112978. [Google Scholar] [CrossRef]
- Xiang, C.; Chen, H.; Liu, X.; Dang, Y.; Li, X.; Yu, Y.; Li, B.; Li, X.; Sun, Y.; Ding, P.; et al. UV-aged microplastics induces neurotoxicity by affecting the neurotransmission in larval zebrafish. Chemosphere 2023, 324, 138252. [Google Scholar] [CrossRef]
- Gao, G.; Zhao, X.; Jin, P.; Gao, K.; Beardall, J. Current understanding and challenges for aquatic primary producers in a world with rising micro- and nano-plastic levels. J. Hazard. Mater. 2021, 406, 124685. [Google Scholar] [CrossRef]
- Kwak, J.I.; An, Y.-J. Microplastic digestion generates fragmented nanoplastics in soils and damages earthworm spermatogenesis and coelomocyte viability. J. Hazard. Mater. 2021, 402, 124034. [Google Scholar] [CrossRef]
- Wei, W.; Yang, Q.; Xiang, D.; Chen, X.; Wen, Z.; Wang, X.; Xu, X.; Peng, C.; Yang, L.; Luo, M.; et al. Combined impacts of microplastics and cadmium on the liver function, immune response, and intestinal microbiota of crucian carp (Carassius carassius). Ecotoxicol. Environ. Saf. 2023, 261, 115104. [Google Scholar] [CrossRef]
- Zheng, Q.; Cui, L.; Liao, H.; Junaid, M.; Li, Z.; Liu, S.; Gao, D.; Zheng, Y.; Lu, S.; Qiu, J.; et al. Combined exposure to polystyrene nanoplastics and bisphenol A induces hepato- and intestinal-toxicity and disturbs gut microbiota in channel catfish (Ictalurus punctatus). Sci. Total Environ. 2023, 891, 164319. [Google Scholar] [CrossRef]
- Li, L.; Xi, W.-S.; Su, Q.; Li, Y.; Yan, G.-H.; Liu, Y.; Wang, H.; Cao, A. Unexpected Size Effect: The Interplay between Different-Sized Nanoparticles in Their Cellular Uptake. Small 2019, 15, e1901687. [Google Scholar] [CrossRef]
- Liang, B.; Zhong, Y.; Huang, Y.; Lin, X.; Liu, J.; Lin, L.; Hu, M.; Jiang, J.; Dai, M.; Wang, B.; et al. Underestimated health risks: Polystyrene micro- and nanoplastics jointly induce intestinal barrier dysfunction by ROS-mediated epithelial cell apoptosis. Part. Fibre Toxicol. 2021, 18, 20. [Google Scholar] [CrossRef]
- Lenz, R.; Enders, K.; Nielsen, T.G. Microplastic exposure studies should be environmentally realistic. Proc. Natl. Acad. Sci. USA 2016, 113, E4121–E4122. [Google Scholar] [CrossRef]
- Al-Sid-Cheikh, M.; Rowland, S.J.; Stevenson, K.; Rouleau, C.; Henry, T.B.; Thompson, R.C. Uptake, whole-body distribution, and depuration of nanoplastics by the scallop Pecten maximus at environmentally realistic concentrations. Environ. Sci. Technol. 2018, 52, 14480–14486. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Liu, H.; Guo, D.; Kong, Y.; Rui, Q.; Wang, D. Damage on functional state of intestinal barrier by microgravity stress in nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2019, 183, 109554. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, Y.; Hua, X.; Wang, D. Induction of transgenerational toxicity is associated with the activated germline insulin signals in nematodes exposed to nanoplastic at predicted environmental concentrations. Ecotoxicol. Environ. Saf. 2022, 243, 114022. [Google Scholar] [CrossRef]
- Yu, C.-W.; Luk, T.C.; Liao, V.H.-C. Long-term nanoplastics exposure results in multi and trans-generational reproduction decline associated with germline toxicity and epigenetic regulation in Caenorhabditis elegans. J. Hazard. Mater. 2021, 412, 125173. [Google Scholar] [CrossRef]
- Yu, Y.; Xie, D.; Yang, Y.; Tan, S.; Li, H.; Dang, Y.; Xiang, M.; Chen, H. Carboxyl-modified polystyrene microplastics induces neurotoxicity by affecting dopamine, glutamate, serotonin, and GABA neurotransmission in Caenorhabditis elegans. J. Hazard. Mater. 2023, 445, 130543. [Google Scholar] [CrossRef]
- Zhao, L.; Qu, M.; Wong, G.; Wang, D. Transgenerational toxicity of nanopolystyrene particles in the range of μg L−1 in the nematode Caenorhabditis elegans. Environ. Sci. Nano 2017, 4, 2356–2366. [Google Scholar] [CrossRef]
- Lei, L.; Wu, S.; Lu, S.; Liu, M.; Song, Y.; Fu, Z.; Shi, H.; Raley-Susman, K.M.; He, D. Microplastic particles cause intestinal damage and other adverse effects in zebrafish Danio rerio and nematode Caenorhabditis elegans. Sci. Total Environ. 2018, 619–620, 1–8. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W.; Wang, Y.; Liu, H.; Zhang, S.; Ji, X.; Qiao, K. Oxidative stress, intestinal damage, and cell apoptosis: Toxicity induced by fluopyram in Caenorhabditis elegans. Chemosphere 2022, 286 Pt 3, 131830. [Google Scholar]
- Yu, Y.; Chen, H.; Hua, X.; Dang, Y.; Han, Y.; Yu, Z.; Chen, X.; Ding, P.; Li, H. Polystyrene microplastics (PS-MPs) toxicity induced oxidative stress and intestinal injury in nematode Caenorhabditis elegans. Sci. Total Environ. 2020, 726, 138679. [Google Scholar] [CrossRef]
- Liu, H.; Tian, L.; Qu, M.; Wang, D. Acetylation regulation associated with the induction of protective response to polystyrene nanoparticles in Caenorhabditis elegans. J. Hazard. Mater. 2021, 411, 125035. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kwak, J.I.; Wang, D.; An, Y.-J. Multigenerational effects of polyethylene terephthalate microfibers in Caenorhabditis elegans. Environ. Res. 2021, 193, 110569. [Google Scholar] [CrossRef] [PubMed]
- Savage-Dunn, C.J.W. TGF-Beta Signaling. In WormBook; 2013; pp. 1–34. [Google Scholar]
- Zhao, Y.; Wu, Q.; Wang, D. An epigenetic signal encoded protection mechanism is activated by graphene oxide to inhibit its induced reproductive toxicity in Caenorhabditis elegans. Biomaterials 2016, 79, 15–24. [Google Scholar] [CrossRef]
- Hua, X.; Feng, X.; Liang, G.; Chao, J.; Wang, D. Long-term exposure to tire-derived 6-PPD quinone causes intestinal toxicity by affecting functional state of intestinal barrier in Caenorhabditis elegans. Sci. Total Environ. 2023, 861, 160591. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-K.; Park, S.-K. Phosphatidylserine modulates response to oxidative stress through hormesis and increases lifespan via DAF-16 in Caenorhabditis elegans. Biogerontology 2020, 21, 231–244. [Google Scholar] [CrossRef]
- Hekimi, S.; Guarente, L. Genetics and the specificity of the aging process. Science 2003, 299, 1351–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Han, X.; Wang, D.; Zhao, F.; Wang, D. Coal combustion related fine particulate matter (PM2.5) induces toxicity in Caenorhabditis elegans by dysregulating microRNA expression. Toxicol. Res. 2017, 6, 432–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanase, S.; Onodera, A.; Tedesco, P.; Johnson, T.E.; Ishii, N. SOD-1 deletions in Caenorhabditis elegans alter the localization of intracellular reactive oxygen species and show molecular compensation. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2009, 64, 530–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirt, N.; Body-Malapel, M. Immunotoxicity and intestinal effects of nano- and microplastics: A review of the literature. Part. Fibre Toxicol. 2020, 17, 57. [Google Scholar] [CrossRef]
- Liu, T.; Hou, B.; Zhang, Y.; Wang, Z. Determination of Biological and Molecular Attributes Related to Polystyrene Microplastic-Induced Reproductive Toxicity and Its Reversibility in Male Mice. Int. J. Environ. Res. Public Health 2022, 19, 14093. [Google Scholar] [CrossRef]
- Liu, X.; Liang, C.; Zhou, M.; Chang, Z.; Li, L. Exposure of Cyprinus carpio var. larvae to PVC microplastics reveals significant immunological alterations and irreversible histological organ damage. Ecotoxicol. Environ. Saf. 2023, 249, 114377. [Google Scholar] [CrossRef]
- Cheng, X.; Dong, S.; Chen, D.; Rui, Q.; Guo, J.; Dayong, W.; Jiang, J. Potential of esterase DmtH in transforming plastic additive dimethyl terephthalate to less toxic mono-methyl terephthalate. Ecotoxicol. Environ. Saf. 2020, 187, 109848. [Google Scholar] [CrossRef]
- Yang, Y.; Dong, W.; Wu, Q.; Wang, D. Response of G protein-coupled receptor CED-1 in germline to polystyrene nanoparticles in Caenorhabditis elegans. Nanoscale Adv. 2021, 3, 1997–2006. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Yin, L.; Pu, Y.; Wang, D. Using the nematode Caenorhabditis elegans as a model animal for assessing the toxicity induced by microcystin-LR. J. Environ. Sci. 2009, 21, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Jia, X.; Guo, T.; Cheng, L.; Han, X.; Wu, Q.; Wang, D. A circular RNA circ_0000115 in response to graphene oxide in nematodes. RSC Adv. 2019, 9, 13722–13735. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Qu, M.; Rui, Q.; Wang, D. Combinational effect of titanium dioxide nanoparticles and nanopolystyrene particles at environmentally relevant concentrations on nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2018, 161, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Nida, A.; Kong, Y.; Du, H.; Xiao, G.; Wang, D. Nanopolystyrene at predicted environmental concentration enhances microcystin-LR toxicity by inducing intestinal damage in Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2019, 183, 109568. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Hu, Y.; Cai, T.; Lin, X.; Wang, D. Regulation of longevity by genes required for the functions of AIY interneuron in nematode Caenorhabditis elegans. Mech. Ageing Dev. 2010, 131, 732–738. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, Z.; Jia, R.; Li, G.; Xi, Z.; Wang, D. Transgenerational effects of traffic-related fine particulate matter (PM2.5) on nematode Caenorhabditis elegans. J. Hazard. Mater. 2014, 274, 106–114. [Google Scholar] [CrossRef]
- Hua, X.; Feng, X.; Liang, G.; Chao, J.; Wang, D. Exposure to 6-PPD Quinone at Environmentally Relevant Concentrations Causes Abnormal Locomotion Behaviors and Neurodegeneration in Caenorhabditis elegans. Environ. Sci. Technol. 2023, 57, 4940–4950. [Google Scholar] [CrossRef]
- Wu, Q.; Yin, L.; Li, X.; Tang, M.; Zhang, T.; Wang, D. Contributions of altered permeability of intestinal barrier and defecation behavior to toxicity formation from graphene oxide in nematode Caenorhabditis elegans. Nanoscale 2013, 5, 9934–9943. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Tan, X.; Shi, X.; Han, P.; Liu, H. Combined Effects of Micro- and Nanoplastics at the Predicted Environmental Concentration on Functional State of Intestinal Barrier in Caenorhabditis elegans. Toxics 2023, 11, 653. https://doi.org/10.3390/toxics11080653
Wu Y, Tan X, Shi X, Han P, Liu H. Combined Effects of Micro- and Nanoplastics at the Predicted Environmental Concentration on Functional State of Intestinal Barrier in Caenorhabditis elegans. Toxics. 2023; 11(8):653. https://doi.org/10.3390/toxics11080653
Chicago/Turabian StyleWu, Yu, Xiaochao Tan, Xian Shi, Peiyu Han, and Huanliang Liu. 2023. "Combined Effects of Micro- and Nanoplastics at the Predicted Environmental Concentration on Functional State of Intestinal Barrier in Caenorhabditis elegans" Toxics 11, no. 8: 653. https://doi.org/10.3390/toxics11080653