
In Vitro Genotoxicity Evaluation of PAHs in Mixtures Using Experimental Design
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Mixture Preparation
2.2. Cellular Culture
2.3. Cytokinesis-Block Micronucleus Assay
2.4. Cytokinesis-Block Micronucleus Assay
2.5. Experimental Factors and Domain of Interest
2.6. Determination of Genotoxic Equivalent Factors
3. Results and Discussion
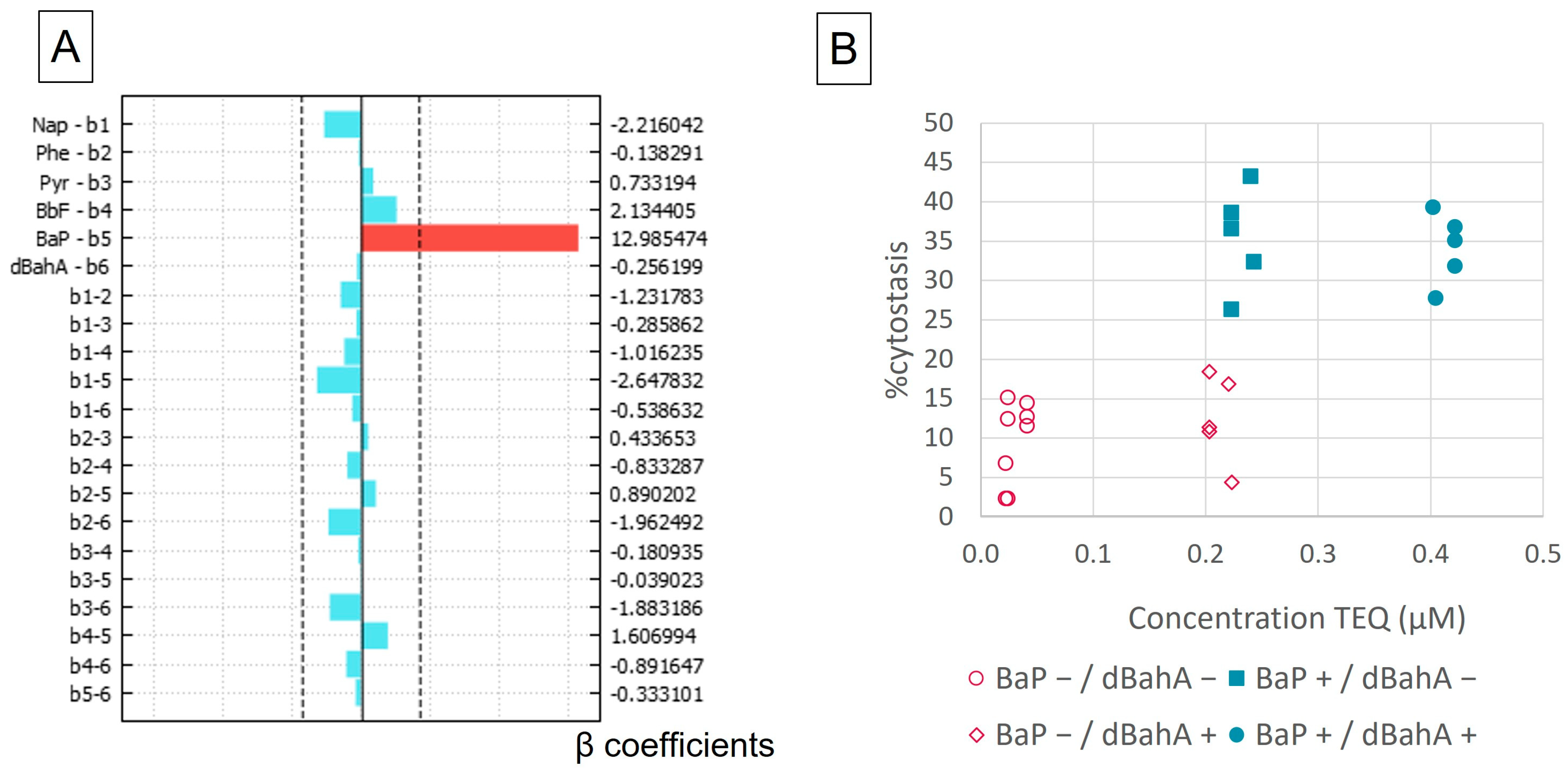
3.1. Cytostasis
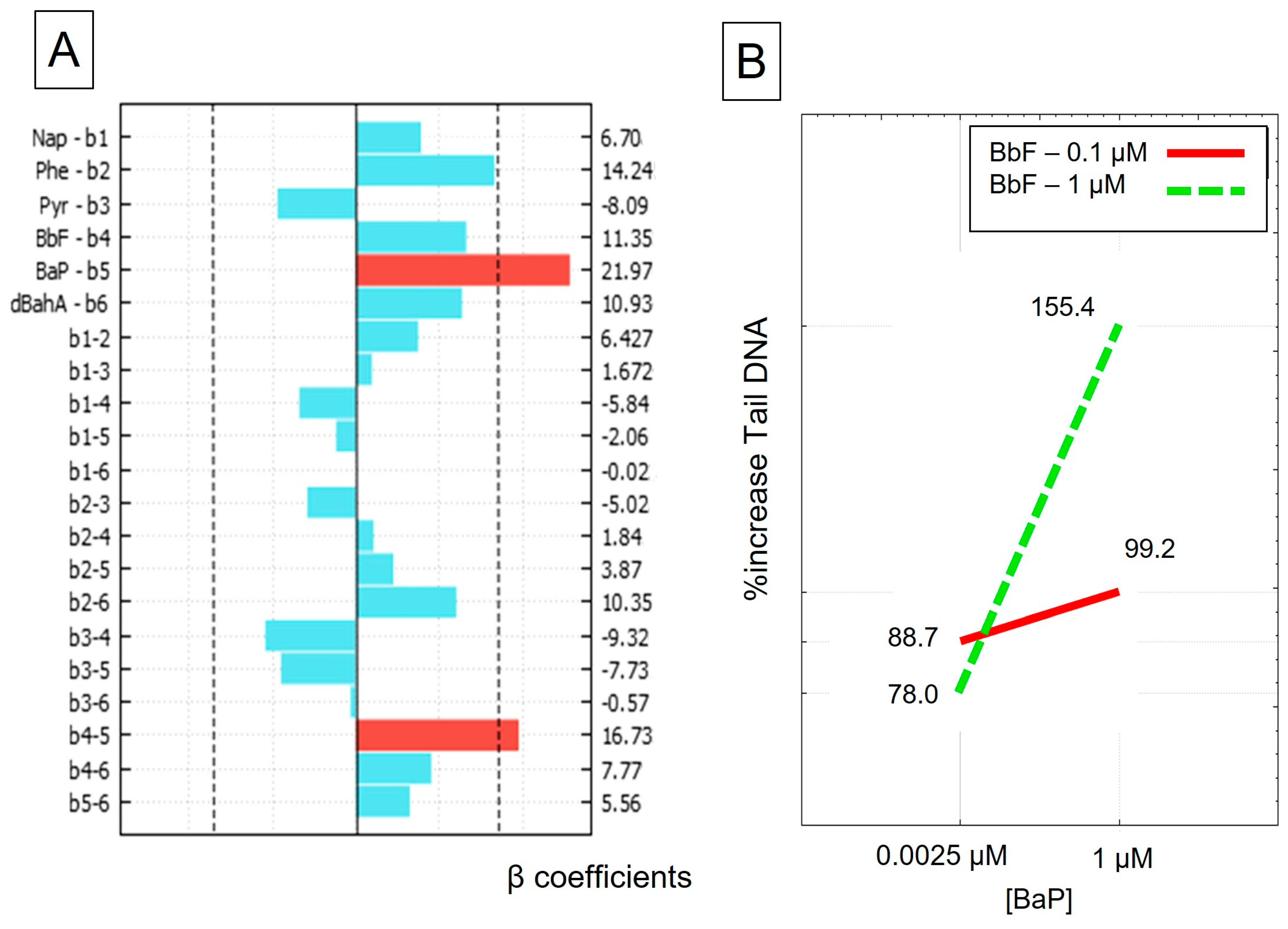
3.2. DNA Damage–Comet Assay
3.3. Chromosomal Damage–CBMN Assay
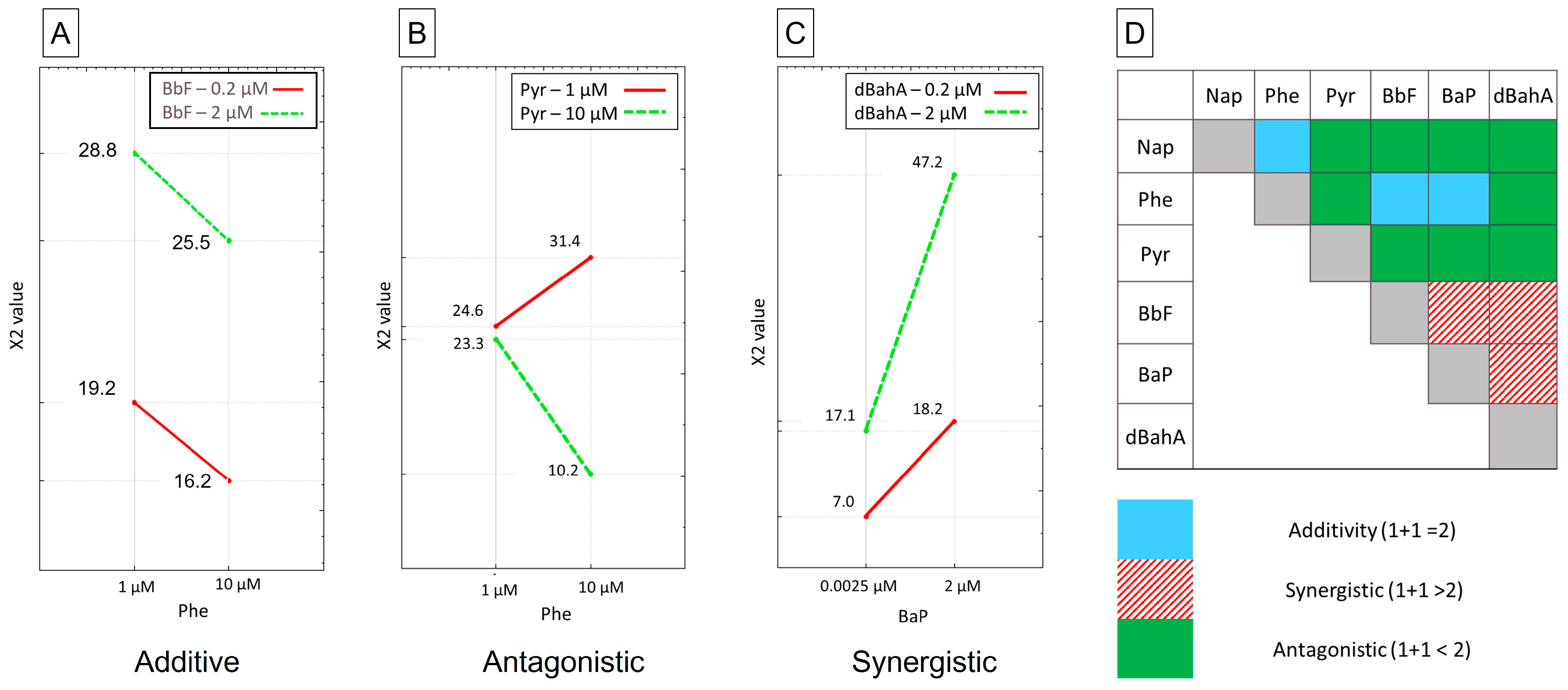
3.4. Determination of Genotoxic Equivalent Factors
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- U.S. EPA. Exposure Factors Handbook Chapter 5 (Update): Soil and Dust Ingestion; U.S. EPA Office of Research and Development: Washington, DC, USA, 2017.
- World Health Organization (Ed.) WHO Guidelines for Indoor Air Quality: Selected Pollutants; WHO: Copenhagen, Denmark, 2010; ISBN 978-92-890-0213-4. [Google Scholar]
- Moya, J.; Bearer, C.F.; Etzel, R.A. Children’s Behavior and Physiology and How It Affects Exposure to Environmental Contaminants. Pediatrics 2004, 113, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Weiss, J.M.; Gustafsson, Å.; Gerde, P.; Bergman, Å.; Lindh, C.H.; Krais, A.M. Daily Intake of Phthalates, MEHP, and DINCH by Ingestion and Inhalation. Chemosphere 2018, 208, 40–49. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (Ed.) Chemical Agents and Related Occupations; International Agency for Research on Cancer: Lyon, France, 2012; ISBN 978-92-832-0138-0.
- Annas, A.; Brittebo, E.; Hellman, B. Evaluation of Benzo(a)Pyrene-Induced DNA Damage in Human Endothelial Cells Using Alkaline Single Cell Gel Electrophoresis. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2000, 471, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Garry, S.; Nesslany, F.; Aliouat, E.; Haguenoer, J.-M.; Marzin, D. Assessment of Genotoxic Effect of Benzo[a]Pyrene in Endotracheally Treated Rat Using the Comet Assay. Mutat. Res. 2003, 534, 33–43. [Google Scholar] [CrossRef]
- Park, S.-Y.; Lee, S.-M.; Ye, S.-K.; Yoon, S.-H.; Chung, M.-H.; Choi, J. Benzo[a]Pyrene-Induced DNA Damage and P53 Modulation in Human Hepatoma HepG2 Cells for the Identification of Potential Biomarkers for PAH Monitoring and Risk Assessment. Toxicol. Lett. 2006, 167, 27–33. [Google Scholar] [CrossRef]
- Labash, C.; Avlasevich, S.L.; Carlson, K.; Berg, A.; Torous, D.K.; Bryce, S.M.; Bemis, J.C.; MacGregor, J.T.; Dertinger, S.D. Mouse Pig-a and Micronucleus Assays Respond to N-Ethyl-N-Nitrosourea, Benzo[a]Pyrene, and Ethyl Carbamate, but Not Pyrene or Methyl Carbamate: Cross-Species Pig-a and Micronucleated Reticulocyte Assays. Environ. Mol. Mutagen. 2016, 57, 28–40. [Google Scholar] [CrossRef]
- Shamsuddin, A.K.M.; Sinopoli, N.T.; Hemminki, K.; Boesch, R.R.; Harris, C.C. Detection of Benzo(a)Pyrene: DNA Adducts in Human White Blood Cells. Cancer Res. 1985, 45, 66–68. [Google Scholar]
- Audebert, M.; Riu, A.; Jacques, C.; Hillenweck, A.; Jamin, E.L.; Zalko, D.; Cravedi, J.-P. Use of the ΓH2AX Assay for Assessing the Genotoxicity of Polycyclic Aromatic Hydrocarbons in Human Cell Lines. Toxicol. Lett. 2010, 199, 182–192. [Google Scholar] [CrossRef]
- Lim, H.; Sadiktsis, I.; de Oliveira Galvão, M.F.; Westerholm, R.; Dreij, K. Polycyclic Aromatic Compounds in Particulate Matter and Indoor Dust at Preschools in Stockholm, Sweden: Occurrence, Sources and Genotoxic Potential in Vitro. Sci. Total Environ. 2021, 755, 142709. [Google Scholar] [CrossRef]
- Sevastyanova, O.; Binkova, B.; Topinka, J.; Sram, R.J.; Kalina, I.; Popov, T.; Novakova, Z.; Farmer, P.B. In Vitro Genotoxicity of PAH Mixtures and Organic Extract from Urban Air Particles Part II: Human Cell Lines. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2007, 620, 123–134. [Google Scholar] [CrossRef]
- Tarantini, A.; Maître, A.; Lefèbvre, E.; Marques, M.; Rajhi, A.; Douki, T. Polycyclic Aromatic Hydrocarbons in Binary Mixtures Modulate the Efficiency of Benzo[a]Pyrene to Form DNA Adducts in Human Cells. Toxicology 2011, 279, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Muthusamy, S.; Xia, Q.; Lal, V.; Denison, M.S.; Ng, J.C. Micronucleus Formation by Single and Mixed Heavy Metals/Loids and PAH Compounds in HepG2 Cells. Mutage 2015, 30, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Mahadevan, B.; Marston, C.P.; Dashwood, W.-M.; Li, Y.; Pereira, C.; Baird, W.M. Effect of a Standardized Complex Mixture Derived from Coal Tar on the Metabolic Activation of Carcinogenic Polycyclic Aromatic Hydrocarbons in Human Cells in Culture. Chem. Res. Toxicol. 2005, 18, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Lacour, B.; Belon, J.-P. Physiologie; Elsevier Masson: Issy-les-Moulineaux, France, 2016; ISBN 978-2-294-75376-3. [Google Scholar]
- Denison, M.S.; Pandini, A.; Nagy, S.R.; Baldwin, E.P.; Bonati, L. Ligand Binding and Activation of the Ah Receptor. Chem. Interactions 2002, 141, 3–24. [Google Scholar] [CrossRef]
- Shimada, T.; Fujii-Kuriyama, Y. Metabolic Activation of Polycyclic Aromatic Hydrocarbons to Carcinogens by Cytochromes P450 1A1 And1B1. Cancer Sci. 2004, 95, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Denison, M.S.; Nagy, S.R. Activation of the Aryl Hydrocarbon Receptor by Structurally Diverse Exogenous and Endogenous Chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, D.O.; Arcaro, K.; Spink, D.C. Understanding the Human Health Effects of Chemical Mixtures. Environ. Health Perspect. 2002, 110, 25–42. [Google Scholar] [CrossRef]
- Nisbet, I.C.T.; LaGoy, P.K. Toxic Equivalency Factors (TEFs) for Polycyclic Aromatic Hydrocarbons (PAHs). Regul. Toxicol. Pharmacol. 1992, 16, 290–300. [Google Scholar] [CrossRef]
- Maertens, R.M.; Gagné, R.W.; Douglas, G.R.; Zhu, J.; White, P.A. Mutagenic and Carcinogenic Hazards of Settled House Dust II: Salmonella Mutagenicity. Environ. Sci. Technol. 2008, 42, 1754–1760. [Google Scholar] [CrossRef]
- Ossai, C.J.; Iwegbue, C.M.A.; Tesi, G.O.; Olisah, C.; Egobueze, F.E.; Nwajei, G.E.; Martincigh, B.S. Distribution, Sources and Exposure Risk of Polycyclic Aromatic Hydrocarbons in Soils, and Indoor and Outdoor Dust from Port Harcourt City, Nigeria. Environ. Sci. Process. Impacts 2021, 23, 1328–1350. [Google Scholar] [CrossRef]
- Živančev, J.; Antić, I.; Buljovčić, M.; Đurišić-Mladenović, N. A Case Study on the Occurrence of Polycyclic Aromatic Hydrocarbons in Indoor Dust of Serbian Households: Distribution, Source Apportionment and Health Risk Assessment. Chemosphere 2022, 295, 133856. [Google Scholar] [CrossRef] [PubMed]
- Staal, Y.C.M.; Hebels, D.G.A.J.; van Herwijnen, M.H.M.; Gottschalk, R.W.H.; van Schooten, F.J.; van Delft, J.H.M. Binary PAH Mixtures Cause Additive or Antagonistic Effects on Gene Expression but Synergistic Effects on DNA Adduct Formation. Carcinogenesis 2007, 28, 2632–2640. [Google Scholar] [CrossRef]
- Muthusamy, S.; Peng, C.; Ng, J.C. Genotoxicity Evaluation of Multi-Component Mixtures of Polyaromatic Hydrocarbons (PAHs), Arsenic, Cadmium, and Lead Using Flow Cytometry Based Micronucleus Test in HepG2 Cells. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2018, 827, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, S.J.; Bruce, E.D. Binary Mixtures of Polycyclic Aromatic Hydrocarbons Display Nonadditive Mixture Interactions in an In Vitro Liver Cell Model. Risk 2016, 36, 968–991. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Polycyclic Aromatic Hydrocarbons in Food—Scientific Opinion of the Panel on Contaminants in the Food Chain. EFSA J. 2008, 724, 724. [Google Scholar] [CrossRef]
- Audebert, M.; Zeman, F.; Beaudoin, R.; Péry, A.; Cravedi, J.-P. Comparative Potency Approach Based on H2AX Assay for Estimating the Genotoxicity of Polycyclic Aromatic Hydrocarbons. Toxicol. Appl. Pharmacol. 2012, 260, 58–64. [Google Scholar] [CrossRef]
- Tomasetig, F.; Tebby, C.; Graillot, V.; Zeman, F.; Pery, A.; Cravedi, J.P.; Audebert, M. Comparative Genotoxic Potential of 27 Polycyclic Aromatic Hydrocarbons in Three Human Cell Lines. Toxicol. Lett. 2020, 326, 99–105. [Google Scholar] [CrossRef]
- Muthusamy, S.; Peng, C.; Ng, J.C. Effects of Binary Mixtures of Benzo[a]Pyrene, Arsenic, Cadmium, and Lead on Oxidative Stress and Toxicity in HepG2 Cells. Chemosphere 2016, 165, 41–51. [Google Scholar] [CrossRef]
- Park, G.-J. Analytic Methods for Design Practice; Springer: London, UK, 2007; ISBN 978-1-84628-473-1. [Google Scholar]
- Deng, H.; Zhang, M.; He, J.; Wu, W.; Jin, L.; Zheng, W.; Lou, J.; Wang, B. Investigating Genetic Damage in Workers Occupationally Exposed to Methotrexate Using Three Genetic End-Points. Mutagenesis 2005, 20, 351–357. [Google Scholar] [CrossRef]
- Sahu, S.C.; Roy, S.; Zheng, J.; Ihrie, J. Contribution of Ionic Silver to Genotoxic Potential of Nanosilver in Human Liver HepG2 and Colon Caco2 Cells Evaluated by the Cytokinesis-Block Micronucleus Assay: Contribution of Ionic Silver to Genotoxic Potential of Nanosilver. J. Appl. Toxicol. 2016, 36, 532–542. [Google Scholar] [CrossRef]
- Pfuhler, S.; Pirow, R.; Downs, T.R.; Haase, A.; Hewitt, N.; Luch, A.; Merkel, M.; Petrick, C.; Said, A.; Schäfer-Korting, M.; et al. Validation of the 3D Reconstructed Human Skin Comet Assay, an Animal-Free Alternative for Following-up Positive Results from Standard in Vitro Genotoxicity Assays. Mutagenesis 2021, 36, 19–35. [Google Scholar] [CrossRef] [PubMed]
- OECD. Test No. 487: In Vitro Mammalian Cell Micronucleus Test. In OECD Guidelines for the Testing of Chemicals; Section 4; OECD: Paris, France, 2016; ISBN 978-92-64-26486-1. [Google Scholar]
- Tice, R.R.; Agurell, E.; Anderson, D.; Burlinson, B.; Hartmann, A.; Kobayashi, H.; Miyamae, Y.; Rojas, E.; Ryu, J.-C.; Sasaki, Y.F. Single Cell Gel/Comet Assay: Guidelines for in Vitro and in Vivo Genetic Toxicology Testing. Environ. Mol. Mutagen. 2000, 35, 206–221. [Google Scholar] [CrossRef]
- Reardon, J.T.; Sancar, A. Nucleotide Excision Repair. Prog. Nucleic Acid Res. Mol. Biol. 2005, 79, 183–235. [Google Scholar] [PubMed]
- Plumejeaud, S.; Reis, A.P.; Tassistro, V.; Patinha, C.; Noack, Y.; Orsière, T. Potentially Harmful Elements in House Dust from Estarreja, Portugal: Characterization and Genotoxicity of the Bioaccessible Fraction. Environ. Geochem. Health 2016, 40, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Botelho, M.C.; Costa, C.; Silva, S.; Costa, S.; Dhawan, A.; Oliveira, P.A.; Teixeira, J.P. Effects of Titanium Dioxide Nanoparticles in Human Gastric Epithelial Cells in Vitro. Biomed. Pharmacother. 2014, 68, 59–64. [Google Scholar] [CrossRef]
- Fenech, M. The in Vitro Micronucleus Technique. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2000, 455, 81–95. [Google Scholar] [CrossRef]
- OECD. Test No. 489: In Vivo Mammalian Alkaline Comet Assay. In OECD Guidelines for the Testing of Chemicals; Section 4; OECD: Paris, France, 2016; ISBN 978-92-64-26488-5. [Google Scholar]
- Rechtschaffner, R.L. Saturated Fractions of 2n and 3n Factorial Designs. Technometrics 1967, 9, 569–575. [Google Scholar] [CrossRef]
- Kutz, F.W.; Barnes, D.G.; Bottimore, D.P.; Greim, H.; Bretthauer, E.W. The International Toxicity Equivalency Factor (I-TEF) Method of Risk Assessment for Complex Mixtures of Dioxins and Related Compounds. Chemosphere 1990, 20, 751–757. [Google Scholar] [CrossRef]
- Van den Berg, M.; Birnbaum, L.; Bosveld, A.T.; Brunström, B.; Cook, P.; Feeley, M.; Giesy, J.P.; Hanberg, A.; Hasegawa, R.; Kennedy, S.W.; et al. Toxic Equivalency Factors (TEFs) for PCBs, PCDDs, PCDFs for Humans and Wildlife. Environ. Health Perspect. 1998, 106, 775–792. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Factors | Principal Effects | Comet Assay | Micronucleus Assay | ||
|---|---|---|---|---|---|---|
| Xi | bi | Level − | Level + | Level − | Level + | |
| X1 | Naphthalene | b1 | 1 | 10 | 1 | 10 |
| X2 | Phenanthrene | b2 | 1 | 10 | 1 | 10 |
| X3 | Pyrene | b3 | 1 | 10 | 1 | 10 |
| X4 | Benzo(b)fluoranthene | b4 | 0.1 | 1 | 0.2 | 2 |
| X5 | Benzo(a)pyrene | b5 | 0.0025 | 1 | 0.0025 | 2 |
| X6 | Dibenz(a,h)anthracene | b6 | 0.1 | 1 | 0.2 | 2 |
| Experiment | Nap | Phe | Pyr | BbF | BaP | dBahA | TEQ | %Increase in Tail DNA |
|---|---|---|---|---|---|---|---|---|
| 1_1 | 1 | 1 | 1 | 0.1 | 0.0025 | 0.1 | 0.12 | 75.41 |
| 1_2 | 1 | 1 | 1 | 0.1 | 0.0025 | 0.1 | 0.12 | 55.16 |
| 1_3 | 1 | 1 | 1 | 0.1 | 0.0025 | 0.1 | 0.12 | 84.94 |
| 2 | 1 | 10 | 10 | 1 | 1 | 1 | 2.12 | 172.36 |
| 3 | 10 | 1 | 10 | 1 | 1 | 1 | 2.12 | 122.65 |
| 4 | 10 | 10 | 1 | 1 | 1 | 1 | 2.12 | 244.26 |
| 5 | 10 | 10 | 10 | 0.1 | 1 | 1 | 2.04 | 141.01 |
| 6 | 10 | 10 | 10 | 1 | 0.0025 | 1 | 1.13 | 109.41 |
| 7 | 10 | 10 | 10 | 1 | 1 | 0.1 | 1.23 | 118.05 |
| 8 | 10 | 10 | 1 | 0.1 | 0.0025 | 0.1 | 0.13 | 104.17 |
| 9 | 10 | 1 | 10 | 0.1 | 0.0025 | 0.1 | 0.13 | 117.40 |
| 10 | 10 | 1 | 1 | 1 | 0.0025 | 0.1 | 0.21 | 61.90 |
| 11 | 10 | 1 | 1 | 0.1 | 1 | 0.1 | 1.12 | 87.87 |
| 12 | 10 | 1 | 1 | 0.1 | 0.0025 | 1 | 1.02 | 60.52 |
| 13 | 1 | 10 | 10 | 0.1 | 0.0025 | 0.1 | 0.13 | 71.06 |
| 14 | 1 | 10 | 1 | 1 | 0.0025 | 0.1 | 0.21 | 73.10 |
| 15 | 1 | 10 | 1 | 0.1 | 1 | 0.1 | 1.12 | 92.09 |
| 16 | 1 | 10 | 1 | 0.1 | 0.0025 | 1 | 1.02 | 82.46 |
| 17 | 1 | 1 | 10 | 1 | 0.0025 | 0.1 | 0.21 | 60.65 |
| 18 | 1 | 1 | 10 | 0.1 | 1 | 0.1 | 1.12 | 77.90 |
| 19 | 1 | 1 | 10 | 0.1 | 0.0025 | 1 | 1.02 | 71.00 |
| 20 | 1 | 1 | 1 | 1 | 1 | 0.1 | 1.20 | 150.34 |
| 21 | 1 | 1 | 1 | 1 | 0.0025 | 1 | 1.11 | 78.98 |
| 22 | 1 | 1 | 1 | 0.1 | 1 | 1 | 2.01 | 80.99 |
| Experiment | Nap | Phe | Pyr | BbF | BaP | dBahA | TEQ | %Cytostasis | χ2 |
|---|---|---|---|---|---|---|---|---|---|
| 1_1 | 1 | 1 | 1 | 0.2 | 0.0025 | 0.2 | 0.23 | 6.67 | 4.27 |
| 1_2 | 1 | 1 | 1 | 0.2 | 0.0025 | 0.2 | 0.23 | −0.63 | 6.08 |
| 1_3 | 1 | 1 | 1 | 0.2 | 0.0025 | 0.2 | 0.23 | 2.20 | 4.76 |
| 2 | 1 | 10 | 10 | 2 | 2 | 2 | 4.22 | 45.11 | 39.69 |
| 3 | 10 | 1 | 10 | 2 | 2 | 2 | 4.22 | 34.92 | 50.78 |
| 4 | 10 | 10 | 1 | 2 | 2 | 2 | 4.22 | 31.68 | 56.01 |
| 5 | 10 | 10 | 10 | 0.2 | 2 | 2 | 4.05 | 27.60 | 5.01 |
| 6 | 10 | 10 | 10 | 2 | 0.0025 | 2 | 2.23 | 4.31 | 0.57 |
| 7 | 10 | 10 | 10 | 2 | 2 | 0.2 | 2.43 | 40.96 | 14.44 |
| 8 | 10 | 10 | 1 | 0.2 | 0.0025 | 0.2 | 0.24 | 9.96 | 8.58 |
| 9 | 10 | 1 | 10 | 0.2 | 0.0025 | 0.2 | 0.24 | 13.99 | 6.27 |
| 10 | 10 | 1 | 1 | 2 | 0.0025 | 0.2 | 0.41 | 12.59 | 1.26 |
| 11 | 10 | 1 | 1 | 0.2 | 2 | 0.2 | 2.23 | 26.18 | 0.62 |
| 12 | 10 | 1 | 1 | 0.2 | 0.0025 | 2 | 2.03 | 18.31 | 2.35 |
| 13 | 1 | 10 | 10 | 0.2 | 0.0025 | 0.2 | 0.24 | 14.99 | 8.49 |
| 14 | 1 | 10 | 1 | 2 | 0.0025 | 0.2 | 0.41 | 11.45 | 18.33 |
| 15 | 1 | 10 | 1 | 0.2 | 2 | 0.2 | 2.23 | 38.46 | 35.05 |
| 16 | 1 | 10 | 1 | 0.2 | 0.0025 | 2 | 2.03 | 10.74 | 29.60 |
| 17 | 1 | 1 | 10 | 2 | 0.0025 | 0.2 | 0.41 | 14.30 | 1.31 |
| 18 | 1 | 1 | 10 | 0.2 | 2 | 0.2 | 2.23 | 34.99 | 15.53 |
| 19 | 1 | 1 | 10 | 0.2 | 0.0025 | 2 | 2.03 | 11.30 | 30.72 |
| 20 | 1 | 1 | 1 | 2 | 2 | 0.2 | 2.4 | 43.09 | 26.86 |
| 21 | 1 | 1 | 1 | 2 | 0.0025 | 2 | 2.21 | 16.79 | 36.53 |
| 22 | 1 | 1 | 1 | 0.2 | 2 | 2 | 4.02 | 39.14 | 64.79 |
| Single PAH | PAH in Mixtures | ||||
|---|---|---|---|---|---|
| PAH | TEF | GEF (γ H2AX) | GEF (CBMN Assay) | GEF (Comet-CBMN Assays) | Mean GEF for PAH Mixtures |
| from [22] | from [31] | from Present Study | |||
| Nap | 0.001 | 0 | 0.10 | 0.06–0 | 0.03 |
| Phe | 0.001 | 0 | 0.01 | 0.08–0.03 | 0.05 |
| Pyr | 0.001 | 0 | 0.01 | 0.02–0 | 0.01 |
| BbF | 0.1 | [0.69–1.46] | 0.22 | 0.74–0.48 | 0.61 |
| dBahA | 1 | [0.5–2.77] | 1.13 | 0.61–1.24 | 0.93 |
| BaP | 1 | 1.00 | 1.00 | 1.00 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castel, R.; Tassistro, V.; Claeys-Bruno, M.; Malleret, L.; Orsière, T. In Vitro Genotoxicity Evaluation of PAHs in Mixtures Using Experimental Design. Toxics 2023, 11, 470. https://doi.org/10.3390/toxics11050470
Castel R, Tassistro V, Claeys-Bruno M, Malleret L, Orsière T. In Vitro Genotoxicity Evaluation of PAHs in Mixtures Using Experimental Design. Toxics. 2023; 11(5):470. https://doi.org/10.3390/toxics11050470
Chicago/Turabian StyleCastel, Rebecca, Virginie Tassistro, Magalie Claeys-Bruno, Laure Malleret, and Thierry Orsière. 2023. "In Vitro Genotoxicity Evaluation of PAHs in Mixtures Using Experimental Design" Toxics 11, no. 5: 470. https://doi.org/10.3390/toxics11050470