

Immunotoxicity and Transcriptome Analyses of Zebrafish (Danio rerio) Embryos Exposed to 6:2 FTSA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Chemicals and Test Solutions
2.2. Adult Zebrafish Rearing and Embryo Collection
2.3. Experimental Design
2.4. Biochemical Index Detection
2.5. Immune-Related Protein Expression
2.6. Transcriptomic Analysis
2.7. Statistical Analysis
3. Results
3.1. Effect of 6:2 FTSA on Oxidative Stress
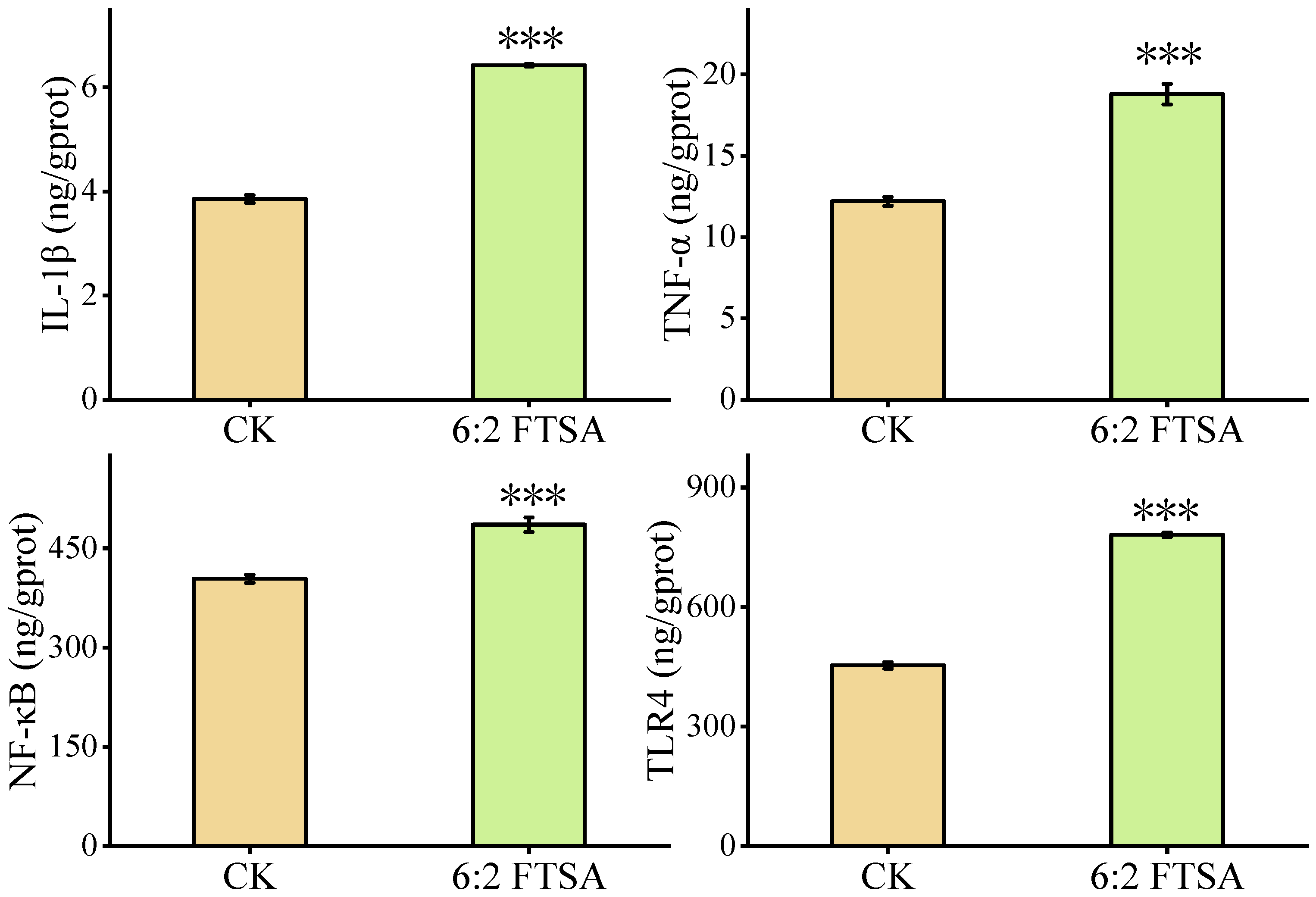
3.2. Effect of 6:2 FTSA on the Inflammatory Response
3.3. Effect of 6:2 FTSA on Immune-Related Proteins
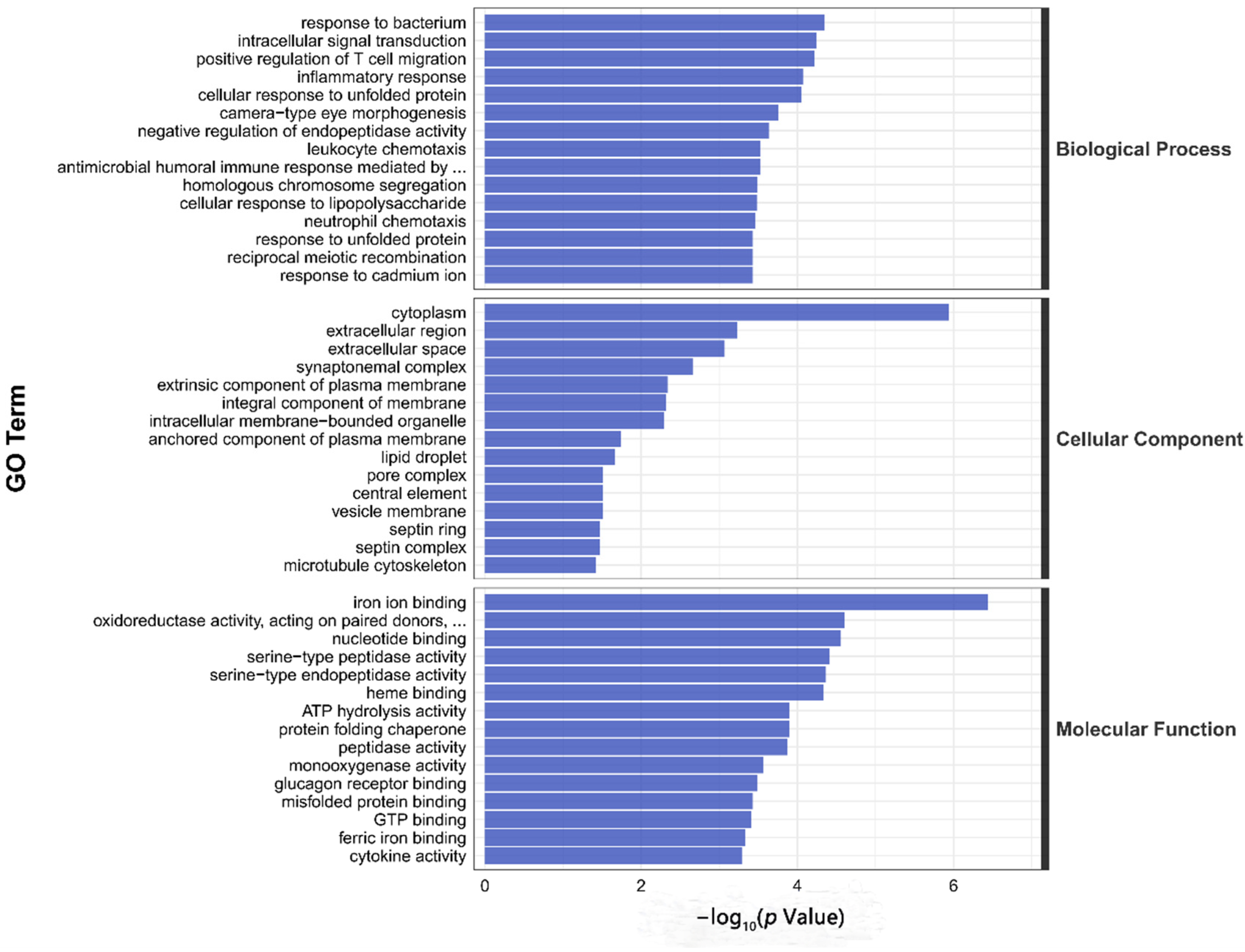
3.4. Transcriptome Sequencing Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chu, S.; Letcher, R.J. Linear and branched perfluorooctane sulfonate isomers in technical product and environmental samples by in-port derivatization-gas chromatography-mass spectrometry. Anal. Chem. 2009, 81, 4256–4262. [Google Scholar] [CrossRef] [PubMed]
- Falandysz, J.; Taniyasu, S.; Gulkowska, A.; Yamashita, N.; Schulte-Oehlmann, U. Is fish a major source of fluorinated surfactants and repellents in humans living on the Baltic Coast? Environ. Sci. Technol. 2006, 40, 748–751. [Google Scholar] [CrossRef] [PubMed]
- Uwayezu, J.-N.; Yeung, L.W.; Bäckström, M. Sorption of PFOS isomers on goethite as a function of pH, dissolved organic matter (humic and fulvic acid) and sulfate. Chemosphere 2019, 233, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Song, X.; Liu, Z.; Ding, X.; Chen, H.; Ding, D. Occurrence, source apportionment, plant bioaccumulation and human exposure of legacy and emerging per- and polyfluoroalkyl substances in soil and plant leaves near a landfill in China. Sci. Total. Environ. 2021, 776, 145731. [Google Scholar] [CrossRef] [PubMed]
- Rayne, S.; Forest, K. Perfluoroalkyl sulfonic and carboxylic acids: A critical review of physicochemical properties, levels and patterns in waters and wastewaters, and treatment methods. J. Environ. Sci. Health Part A 2009, 44, 1145–1199. [Google Scholar] [CrossRef]
- Jantzen, C.E.; Annunziato, K.M.; Cooper, K.R. Behavioral, morphometric, and gene expression effects in adult zebrafish (Danio rerio) embryonically exposed to PFOA, PFOS, and PFNA. Aquat. Toxicol. 2016, 180, 123–130. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Ge, X.; Wang, D.; Wang, T.; Zhang, L.; Tanguay, R.L.; Simonich, M.; Huang, C.; Dong, Q. Chronic perfluorooctanesulphonic acid (PFOS) exposure produces estrogenic effects in zebrafish. Environ. Pollut. 2016, 218, 702–708. [Google Scholar] [CrossRef]
- Schröter-Kermani, C.; Müller, J.; Jürling, H.; Conrad, A.; Schulte, C. Retrospective monitoring of perfluorocarboxylates and perfluorosulfonates in human plasma archived by the German Environmental Specimen Bank. Int. J. Hyg. Environ. Health 2013, 216, 633–640. [Google Scholar] [CrossRef]
- Louisse, J.; Rijkers, D.; Stoopen, G.; Janssen, A.; Staats, M.; Hoogenboom, R.; Kersten, S.; Peijnenburg, A. Perfluorooctanoic acid (PFOA), perfluorooctane sulfonic acid (PFOS), and perfluorononanoic acid (PFNA) increase triglyceride levels and decrease cholesterogenic gene expression in human HepaRG liver cells. Arch. Toxicol. 2020, 94, 3137–3155. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, K.; Li, W.; Chai, Y.; Zhu, J.; Chu, B.; Li, N.; Yan, J.; Zhang, S.; Yang, Y. Varied thyroid disrupting effects of perfluorooctanoic acid (PFOA) and its novel alternatives hexafluoropropylene-oxide-dimer-acid (GenX) and ammonium 4,8-dioxa-3H-perfluorononanoate (ADONA) in vitro. Environ. Int. 2021, 156, 106745. [Google Scholar] [CrossRef]
- Coperchini, F.; Croce, L.; Denegri, M.; Pignatti, P.; Agozzino, M.; Netti, G.S.; Imbriani, M.; Rotondi, M.; Chiovato, L. Adverse effects of in vitro GenX exposure on rat thyroid cell viability, DNA integrity and thyroid-related genes expression. Environ. Pollut. 2020, 264, 114778. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lai, H.; Wang, Q.; Martínez, R.; Zhang, M.; Liu, Y.; Huang, J.; Deng, M.; Tu, W. Immunotoxicity of F 53B, an alternative to PFOS, on zebrafish (Danio rerio) at different early life stages. Sci. Total. Environ. 2021, 790, 148165. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.J.; Pyo, M.C.; Park, Y.; Kim, B.Y.; Lee, K.-W. Hexafluoropropylene oxide dimer acid (GenX) exposure induces apoptosis in HepG2 cells. Heliyon 2021, 7, e08272. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huang, J.; Zhang, K.; Yu, G.; Deng, S.; Wang, B. Stability of 6:2 fluorotelomer sulfonate in advanced oxidation processes: Degradation kinetics and pathway. Environ. Sci. Pollut. Res. 2013, 21, 4634–4642. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-H.; Shi, Y.; Strynar, M.; Chu, K.-H. Desulfonation and defluorination of 6:2 fluorotelomer sulfonic acid (6:2 FTSA) by Rhodococcus jostii RHA1: Carbon and sulfur sources, enzymes, and pathways. J. Hazard. Mater. 2021, 423, 127052. [Google Scholar] [CrossRef] [PubMed]
- Sheng, N.; Zhou, X.; Zheng, F.; Pan, Y.; Guo, X.; Guo, Y.; Sun, Y.; Dai, J. Comparative hepatotoxicity of 6:2 fluorotelomer carboxylic acid and 6:2 fluorotelomer sulfonic acid, two fluorinated alternatives to long-chain perfluoroalkyl acids, on adult male mice. Arch. Toxicol. 2016, 91, 2909–2919. [Google Scholar] [CrossRef]
- Marchiandi, J.; Szabo, D.; Dagnino, S.; Green, M.P.; Clarke, B.O. Occurrence and fate of legacy and novel per- and polyfluoroalkyl substances (PFASs) in freshwater after an industrial fire of unknown chemical stockpiles. Environ. Pollut. 2021, 278, 116839. [Google Scholar] [CrossRef]
- Wang, S.; Ding, G.; Liu, Y.; Dou, Z.; Chen, H.; Ya, M.; Lin, X.; Li, Q.; Li, Y.; Wang, X. Legacy and emerging persistent organic pollutants in the marginal seas of China: Occurrence and phase partitioning. Sci. Total. Environ. 2022, 827, 154274. [Google Scholar] [CrossRef]
- Feng, X.; Ye, M.; Li, Y.; Zhou, J.; Sun, B.; Zhu, Y.; Zhu, L. Potential sources and sediment-pore water partitioning behaviors of emerging per/polyfluoroalkyl substances in the South Yellow Sea. J. Hazard. Mater. 2020, 389, 122124. [Google Scholar] [CrossRef]
- Åkerblom, S.; Negm, N.; Wu, P.; Bishop, K.; Ahrens, L. Variation and accumulation patterns of poly- and perfluoroalkyl substances (PFAS) in European perch (Perca fluviatilis) across a gradient of pristine Swedish lakes. Sci. Total Environ. 2017, 599-600, 1685–1692. [Google Scholar] [CrossRef]
- Munoz, G.; Desrosiers, M.; Vetter, L.; Duy, S.V.; Jarjour, J.; Liu, J.; Sauvé, S. Bioaccumulation of Zwitterionic Polyfluoroalkyl Substances in Earthworms Exposed to Aqueous Film-Forming Foam Impacted Soils. Environ. Sci. Technol. 2020, 54, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Kaboré, H.A.; Goeury, K.; Desrosiers, M.; Duy, S.V.; Liu, J.; Cabana, G.; Munoz, G.; Sauvé, S. Novel and legacy per- and polyfluoroalkyl substances (PFAS) in freshwater sporting fish from background and firefighting foam impacted ecosystems in Eastern Canada. Sci. Total. Environ. 2021, 816, 151563. [Google Scholar] [CrossRef] [PubMed]
- Hoke, R.A.; Ferrell, B.D.; Ryan, T.; Sloman, T.L.; Green, J.W.; Nabb, D.L.; Mingoia, R.; Buck, R.C.; Korzeniowski, S.H. Aquatic hazard, bioaccumulation and screening risk assessment for 6:2 fluorotelomer sulfonate. Chemosphere 2015, 128, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Menger, F.; Pohl, J.; Ahrens, L.; Carlsson, G.; Örn, S. Behavioural effects and bioconcentration of per- and polyfluoroalkyl substances (PFASs) in zebrafish (Danio rerio) embryos. Chemosphere 2019, 245, 125573. [Google Scholar] [CrossRef] [PubMed]
- Martínez, R.; Navarro-Martín, L.; Luccarelli, C.; Codina, A.E.; Raldúa, D.; Barata, C.; Tauler, R.; Piña, B. Unravelling the mechanisms of PFOS toxicity by combining morphological and transcriptomic analyses in zebrafish embryos. Sci. Total. Environ. 2019, 674, 462–471. [Google Scholar] [CrossRef]
- Dang, Y.; Wang, F.; Liu, C. Real-time PCR array to study the effects of chemicals on the growth hormone/insulin-like growth factors (GH/IGFs) axis of zebrafish embryos/larvae. Chemosphere 2018, 207, 365–376. [Google Scholar] [CrossRef]
- Chen, J.; Tanguay, R.L.; Tal, T.L.; Gai, Z.; Ma, X.; Bai, C.; Tilton, S.C.; Jin, D.; Yang, D.; Huang, C.; et al. Early life perfluorooctanesulphonic acid (PFOS) exposure impairs zebrafish organogenesis. Aquat. Toxicol. 2014, 150, 124–132. [Google Scholar] [CrossRef]
- Guo, J.; Wu, P.; Cao, J.; Luo, Y.; Chen, J.; Wang, G.; Guo, W.; Wang, T.; He, X. The PFOS disturbed immunomodulatory functions via nuclear Factor-kappaB signaling in liver of zebrafish (Danio rerio). Fish Shellfish Immunol. 2019, 91, 87–98. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Liang, L.; Pan, Y.; Bin, L.; Liu, Y.; Huang, W.; Li, R.; Lai, K.P. Immunotoxicity mechanisms of perfluorinated compounds PFOA and PFOS. Chemosphere 2021, 291, 132892. [Google Scholar] [CrossRef]
- Huang, J.; Wang, Q.; Liu, S.; Lai, H.; Tu, W. Comparative chronic toxicities of PFOS and its novel alternatives on the immune system associated with intestinal microbiota dysbiosis in adult zebrafish. J. Hazard. Mater. 2022, 425, 127950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liu, K.; Li, J.; Chen, L.; Lin, K. Uptake and depuration kinetics of lead (Pb) and biomarker responses in the earthworm Eisenia fetida after simultaneous exposure to decabromodiphenyl ether (BDE209). Ecotoxicol. Environ. Saf. 2015, 113, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, Y.; Zhang, J.-Y.; Lin, H.; Chen, J.; Hong, H. The toxicity of 2,6-dichlorobenzoquinone on the early life stage of zebrafish: A survey on the endpoints at developmental toxicity, oxidative stress, genotoxicity and cytotoxicity. Environ. Pollut. 2018, 245, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wu, Y.; Wang, Q.; Wan, J.; Deng, M.; Tu, W. Comparison of toxicokinetics and toxic effects of PFOS and its novel alternative OBS in zebrafish larvae. Chemosphere 2020, 265, 129116. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, J.; Deng, M.; Jin, Y.; Yang, H.; Liu, Y.; Cao, Q.; Mennigen, J.A.; Tu, W. Acute exposure to environmentally relevant concentrations of Chinese PFOS alternative F-53B induces oxidative stress in early developing zebrafish. Chemosphere 2019, 235, 945–951. [Google Scholar] [CrossRef]
- Zhang, C.; Du, Z.; Wang, J.; Wang, J.; Zhou, T.; Li, B.; Zhu, L.; Li, W.; Hou, K. Exposed zebrafish (Danio rerio) to imidazolium-based ionic liquids with different anions and alkyl-chain lengths. Chemosphere 2018, 203, 381–386. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiao, Y.; Li, Z.; Tao, Y.; Yang, Y. Hazards of phthalates (PAEs) exposure: A review of aquatic animal toxicology studies. Sci. Total. Environ. 2021, 771, 145418. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, J.; Du, Z.; Li, B.; Zhu, L.; Wang, J.; Zhang, S. Toxic effect of [Omim]BF 4 and [Omim]Br on antioxidant stress and oxidative damage in earthworms (Eisenia fetida). Environ. Toxicol. Pharmacol. 2018, 60, 37–44. [Google Scholar] [CrossRef]
- Zheng, J.-L.; Yuan, S.-S.; Wu, C.-W.; Li, W.-Y. Chronic waterborne zinc and cadmium exposures induced different responses towards oxidative stress in the liver of zebrafish. Aquat. Toxicol. 2016, 177, 261–268. [Google Scholar] [CrossRef]
- Wu, Y.; Deng, M.; Jin, Y.; Mu, X.; He, X.; Luu, N.-T.; Yang, C.; Tu, W. Uptake and elimination of emerging polyfluoroalkyl substance F-53B in zebrafish larvae: Response of oxidative stress biomarkers. Chemosphere 2018, 215, 182–188. [Google Scholar] [CrossRef]
- Wang, S.; Zhuang, C.; Du, J.; Wu, C.; You, H. The presence of MWCNTs reduces developmental toxicity of PFOS in early life stage of zebrafish. Environ. Pollut. 2017, 222, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Cai, J.; Wang, S.; You, H. Oxidative stress and apotosis to Zebrafish (Danio rerio) embryos exposed to perfluorooctane sulfonate (PFOS) and ZnO nanoparticles. Int. J. Occup. Med. Environ. Health 2017, 30, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, Q.; Liu, S.; Zhang, M.; Liu, Y.; Sun, L.; Wu, Y.; Tu, W. Crosstalk between histological alterations, oxidative stress and immune aberrations of the emerging PFOS alternative OBS in developing zebrafish. Sci. Total. Environ. 2021, 774, 145443. [Google Scholar] [CrossRef]
- Yang, H.; Lai, H.; Huang, J.; Sun, L.; Mennigen, J.A.; Wang, Q.; Liu, Y.; Jin, Y.; Tu, W. Polystyrene microplastics decrease F–53B bioaccumulation but induce inflammatory stress in larval zebrafish. Chemosphere 2020, 255, 127040. [Google Scholar] [CrossRef]
- Bian, K.; Harari, Y.; Zhong, M.; Lai, M.; Castro, G.; Weisbrodt, N.; Murad, F. Down-Regulation of Inducible Nitric-Oxide Synthase (NOS-2) During Parasite-Induced Gut Inflammation: A Path to Identify a Selective NOS-2 Inhibitor. Mol. Pharmacol. 2001, 59, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, M.; Jeong, Y. Purification and characterization of lysozyme from filipino venus, Ruditapes philippinarum. Food Sci. Biotechnol. 2012, 21, 1463–1468. [Google Scholar] [CrossRef]
- Chen, M.; Guo, T.; He, K.; Zhu, L.; Jin, H.; Wang, Q.; Liu, M.; Yang, L. Biotransformation and bioconcentration of 6:2 and 8:2 polyfluoroalkyl phosphate diesters in common carp (prinus carpio): Underestimated ecological risks. Sci. Total. Environ. 2018, 656, 201–208. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Y.; Ma, X.; Sun, W.; Liu, H. Perfluorooctanoic acid-induced lipid metabolism disorder in SD rat liver and its effect on the expression of fatty acid metabolism-related proteins. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2022, 47, 18–25. [Google Scholar]
- Bonato, M.; Corrà, F.; Bellio, M.; Guidolin, L.; Tallandini, L.; Irato, P.; Santovito, G. PFAS Environmental Pollution and Antioxidant Responses: An Overview of the Impact on Human Field. Int. J. Environ. Res. Public Health 2020, 17, 8020. [Google Scholar] [CrossRef]
- Park, S.; Moon, N.R.; Kang, S.; Kim, D. Ferulic acid and vinpocetine intake improves memory function by enhancing insulin sensitivity and reducing neuroinflammation and oxidative stress in type 2 diabetic animals with induced Alzheimer’s disease. J. Funct. Foods 2022, 95, 105180. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Li, H.; Huna, R.; Tan, X.; Li, N.; Zhang, Y.; Jiao, X.; Liu, M. C5aR antagonist inhibits LPS-induced inflammation in human gingival fibroblasts via NF-kappaB and MAPK signaling pathways. J. Appl. Oral. Sci. 2023, 31, e20220404. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, L.; Fang, W.; Zhang, X.; Zhong, Y. Perfluorooctanoic acid-induced immunotoxicity via NF-kappa B pathway in zebrafish (Danio rerio) kidney. Fish Shellfish. Immunol. 2021, 113, 9–19. [Google Scholar] [CrossRef]
- Zhang, H.; Fang, W.; Wang, D.; Gao, N.; Ding, Y.; Chen, C. The role of interleukin family in perfluorooctanoic acid (PFOA)-induced immunotoxicity. J. Hazard. Mater. 2014, 280, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Kutikhin, A.G.; E Yuzhalin, A. C-type lectin receptors and RIG-I-like receptors: New points on the oncogenomics map. Cancer Manag. Res. 2012, 4, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Cyktor, J.C.; Mellors, J.W. Toll-Like Receptor Agonists: Can They Exact a Toll on Human Immunodeficiency Virus Persistence? Clin. Infect. Dis. 2017, 64, 1696–1698. [Google Scholar] [CrossRef]
- Kent, A.; Blander, J.M. Nod-Like Receptors: Key Molecular Switches in the Conundrum of Cancer. Front. Immunol. 2014, 5, 185. [Google Scholar] [CrossRef]
- Jing, L.; Zheng, D.; Sun, X.; Shi, Z. DBDPE upregulates NOD-like receptor signaling to induce NLRP3 inflammasome-mediated HAECs pyroptosis. Environ. Pollut. 2023, 318, 120882. [Google Scholar] [CrossRef]
- Shaw, D.K.; McClure, E.E.; Wang, X.; Pedra, J.H.F. Deviant Behavior: Tick-Borne Pathogens and Inflammasome Signaling. Veter- Sci. 2016, 3, 27. [Google Scholar] [CrossRef]
- Ngo, V.N.; Young, R.M.; Schmitz, R.; Jhavar, S.; Xiao, W.; Lim, K.H.; Kohlhammer, H.; Xu, W.; Yang, Y.; Zhao, H.; et al. Oncogenically active MYD88 mutations in human lymphoma. Nature 2011, 470, 115–119. [Google Scholar] [CrossRef]
- Chang, M.X.; Wang, Y.P.; Nie, P. Zebrafish peptidoglycan recognition protein SC (zfPGRP-SC) mediates multiple intracellular signaling pathways. Fish Shellfish Immunol. 2009, 26, 264–274. [Google Scholar] [CrossRef]
- Farkhondeh, T.; Mehrpour, O.; Buhrmann, C.; Pourbagher-Shahri, A.M.; Shakibaei, M.; Samarghandian, S. Organophosphorus Compounds and MAPK Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 4258. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Qiu, W.; Zhang, S.; Wang, J.; Yang, X.; Xu, B.; Magnuson, J.T.; Xu, E.G.; Wu, M.; Zheng, C. Poly- and Perfluoroalkyl Substances Induce Immunotoxicity via the TLR Pathway in Zebrafish: Links to Carbon Chain Length. Environ. Sci. Technol. 2023, 15, 6139–6149. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Zhou, B. The Role of Nrf2 and MAPK Pathways in PFOS-Induced Oxidative Stress in Zebrafish Embryos. Toxicol. Sci. 2010, 115, 391–400. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Ren, Z.; Chen, M. Immunotoxicity and Transcriptome Analyses of Zebrafish (Danio rerio) Embryos Exposed to 6:2 FTSA. Toxics 2023, 11, 459. https://doi.org/10.3390/toxics11050459
Zhang J, Ren Z, Chen M. Immunotoxicity and Transcriptome Analyses of Zebrafish (Danio rerio) Embryos Exposed to 6:2 FTSA. Toxics. 2023; 11(5):459. https://doi.org/10.3390/toxics11050459
Chicago/Turabian StyleZhang, Jing, Zongming Ren, and Meng Chen. 2023. "Immunotoxicity and Transcriptome Analyses of Zebrafish (Danio rerio) Embryos Exposed to 6:2 FTSA" Toxics 11, no. 5: 459. https://doi.org/10.3390/toxics11050459