
Palliative Role of Zamzam Water against Cyclosporine-Induced Nephrotoxicity through Modulating Autophagy and Apoptosis Crosstalk

 ,
,  ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
2.3. Experimental Design
2.4. Specimen Collection
2.5. Biochemical Examination
2.5.1. Renal Function Parameters
2.5.2. Assessment of Oxidative Stress Biomarkers
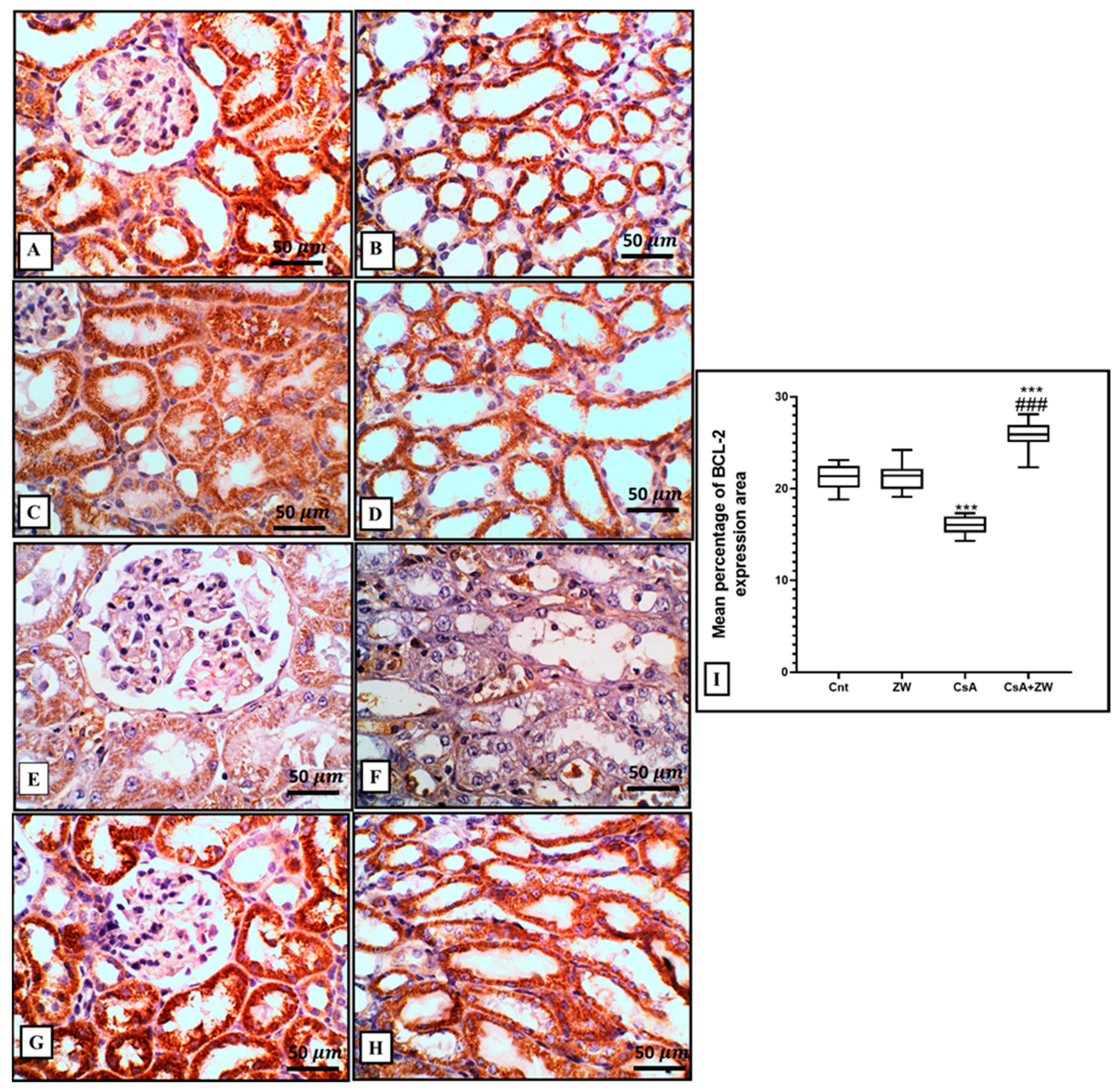
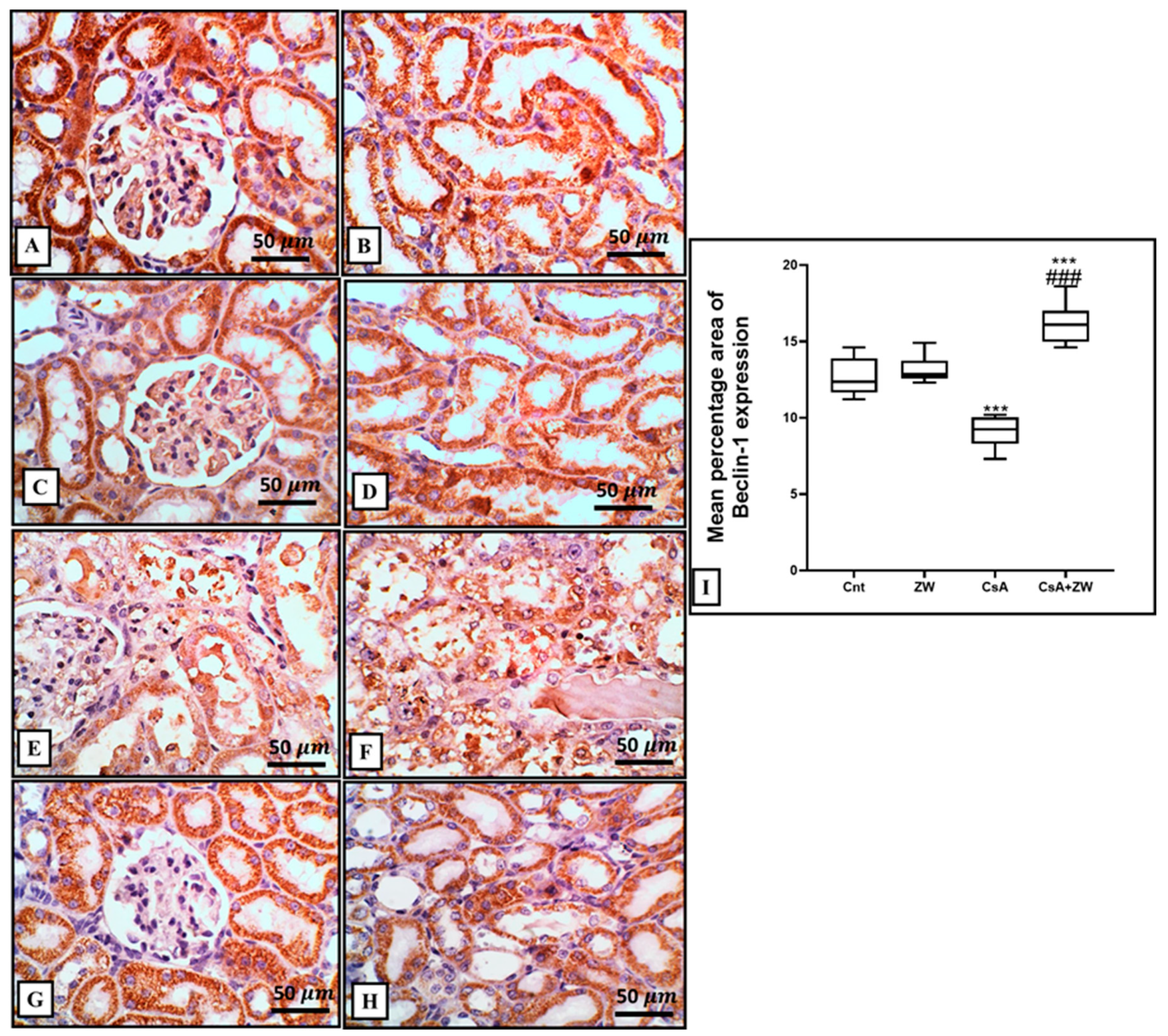
2.6. Histopathological and Immunohistochemical Examination
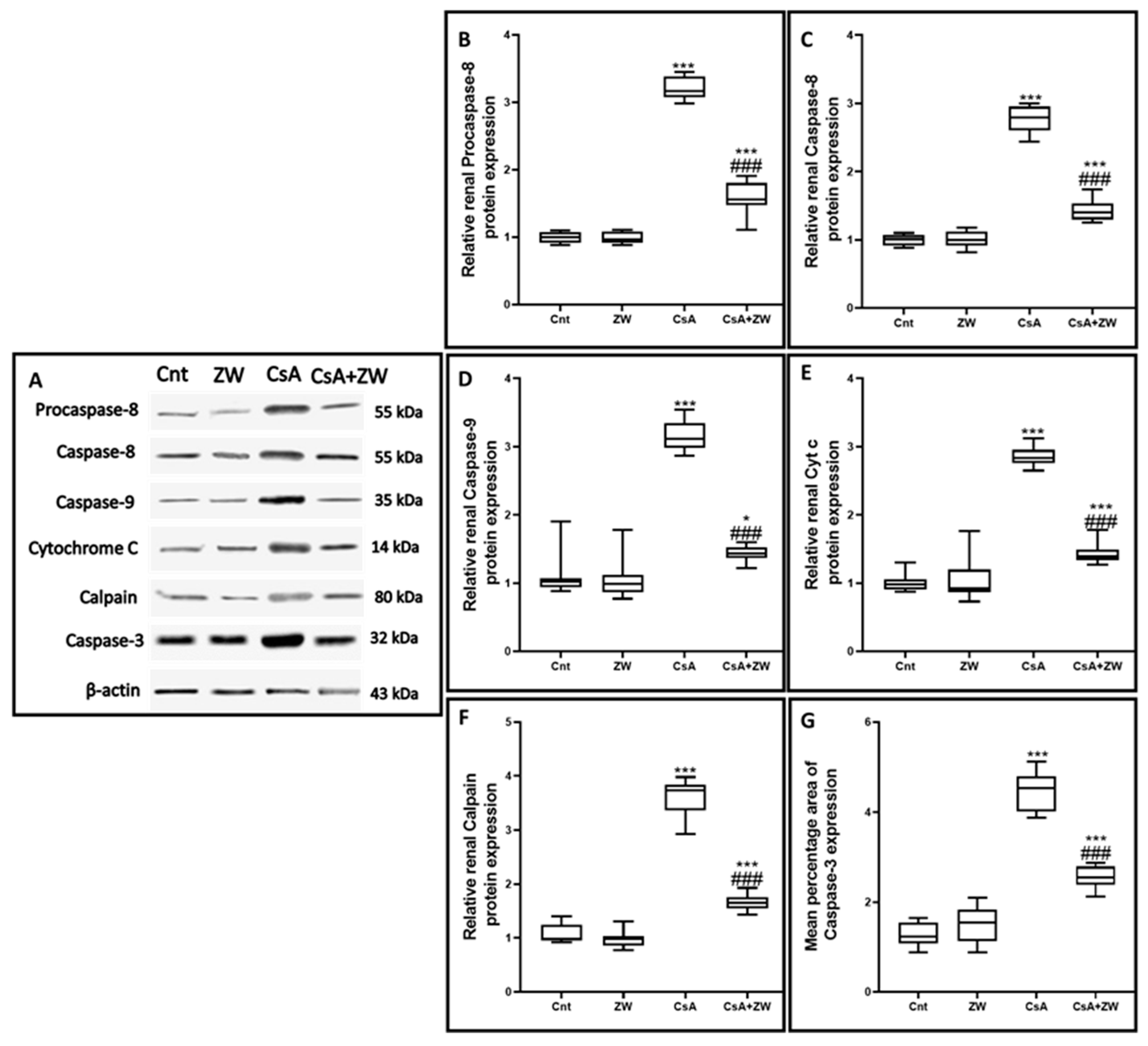
2.7. Western Blotting of Autophagy-Activating Kinase ULK1, Atg5, Procaspase-8, Caspase-8, Caspase-9, Calpain, and Cytochrome c
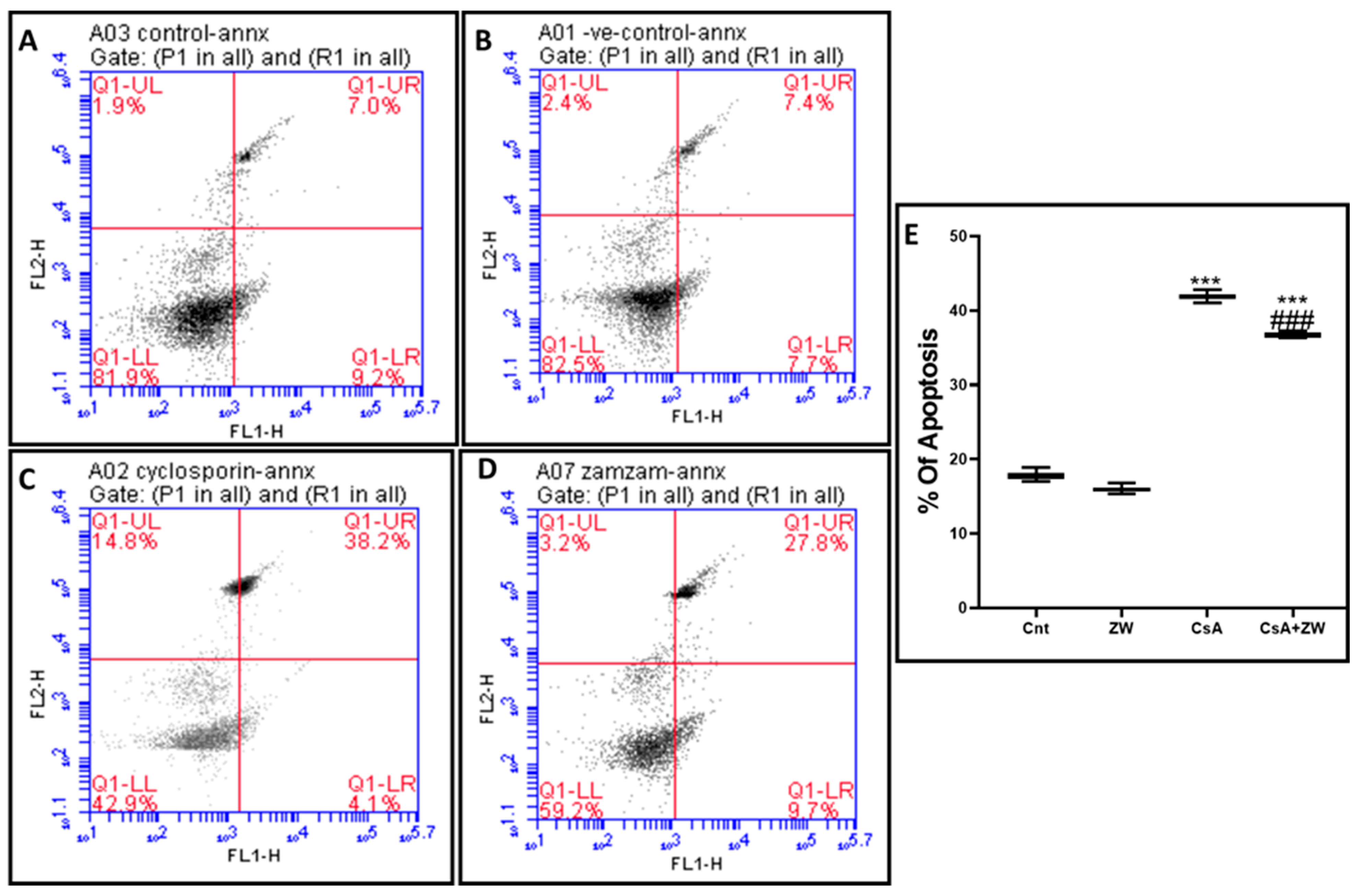
2.8. Analysis of Apoptosis with Flow Cytometry
2.9. RT-PCR Assessment
2.10. Analysis of Zamzam Water
2.11. Statistical Analysis
3. Results
3.1. Effect of Zamzam Water on CsA A-Induced Renal Impairment
3.2. Effect of Zamzam Water against CsA-Induced Oxidative Stress
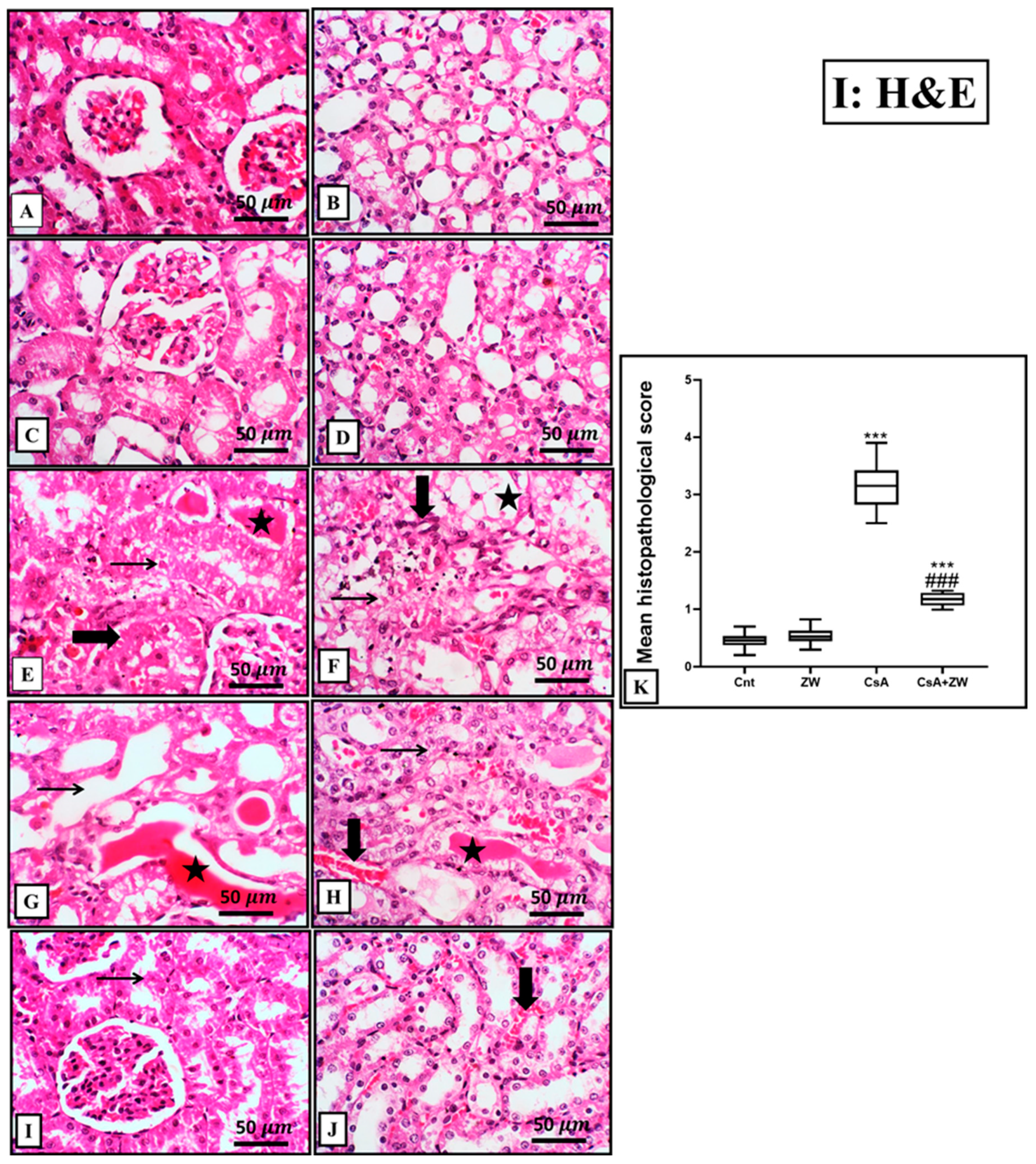
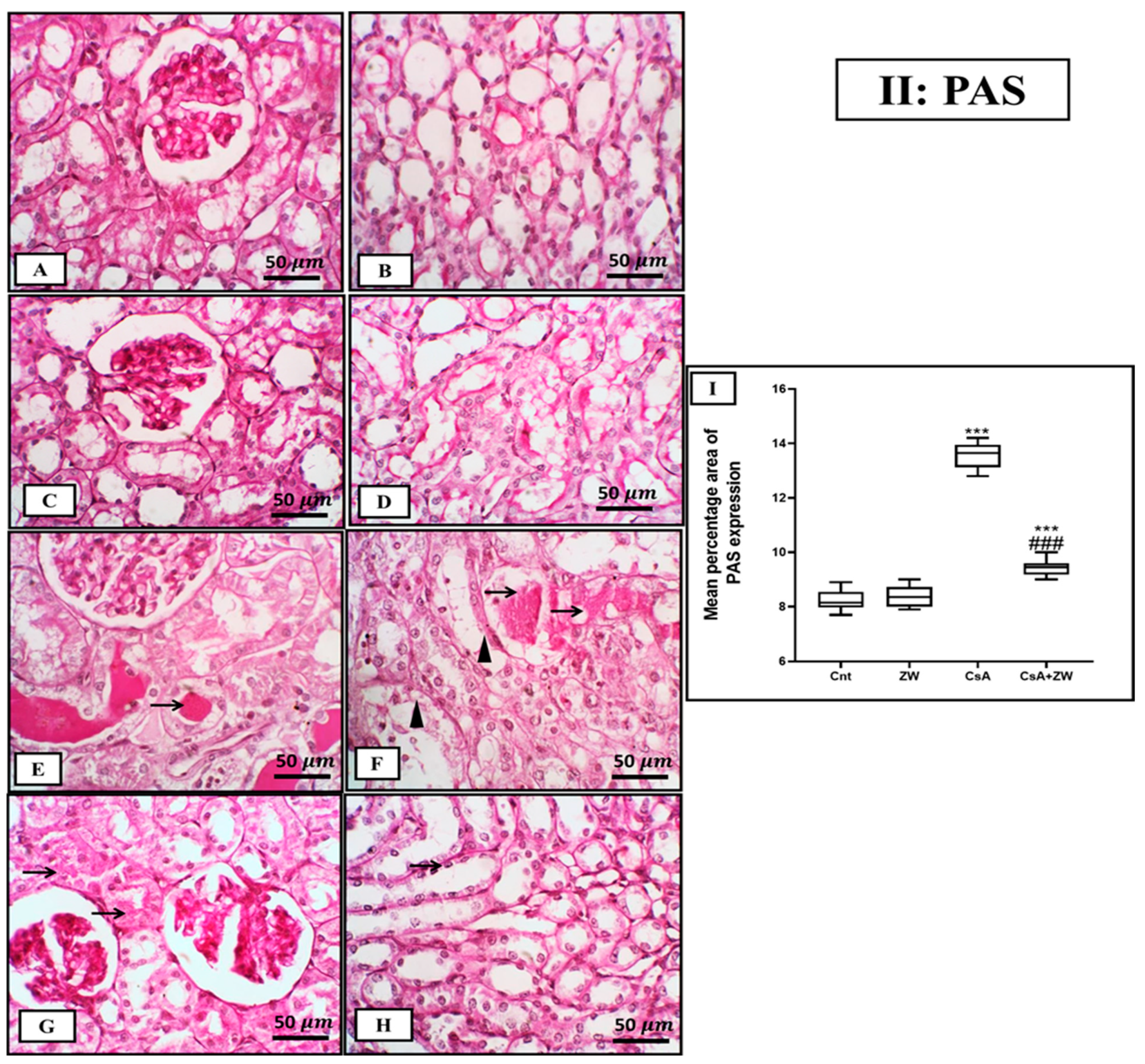
3.3. Ameliorative Impact of Zamzam Water on Renal Morphology
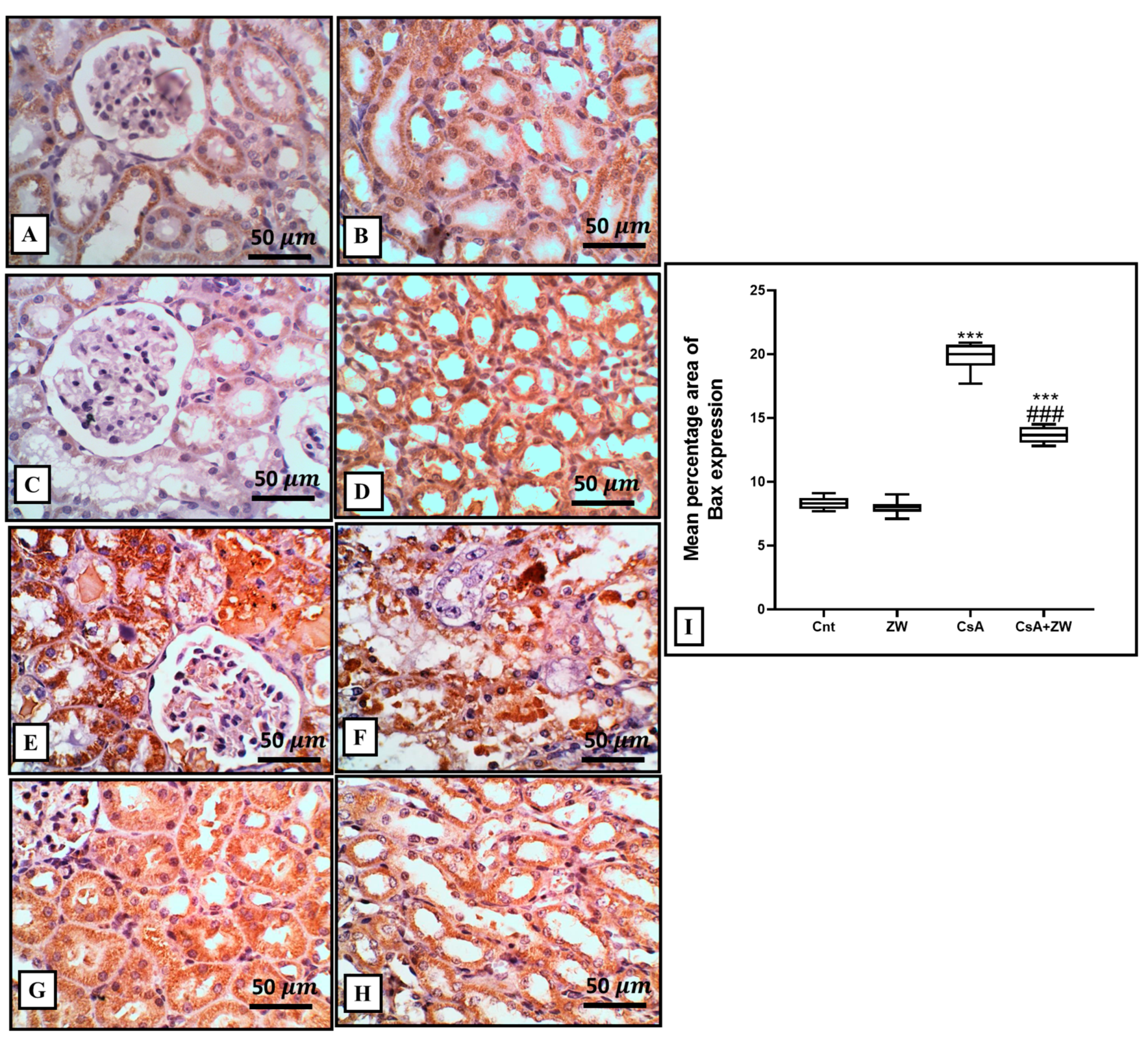
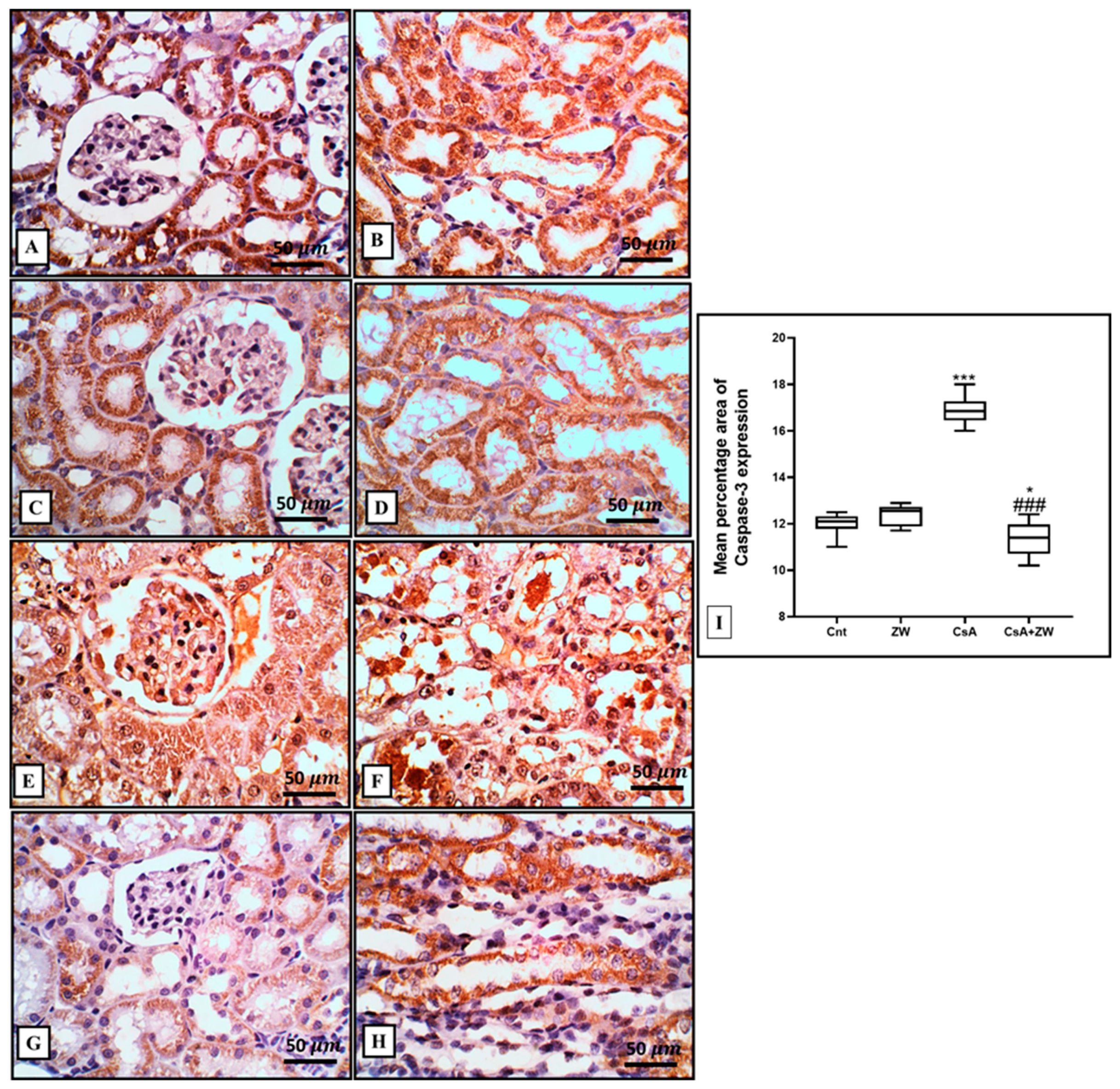
3.4. Effect of Zamzam Water against CsA-Induced Renal Tubular Apoptosis
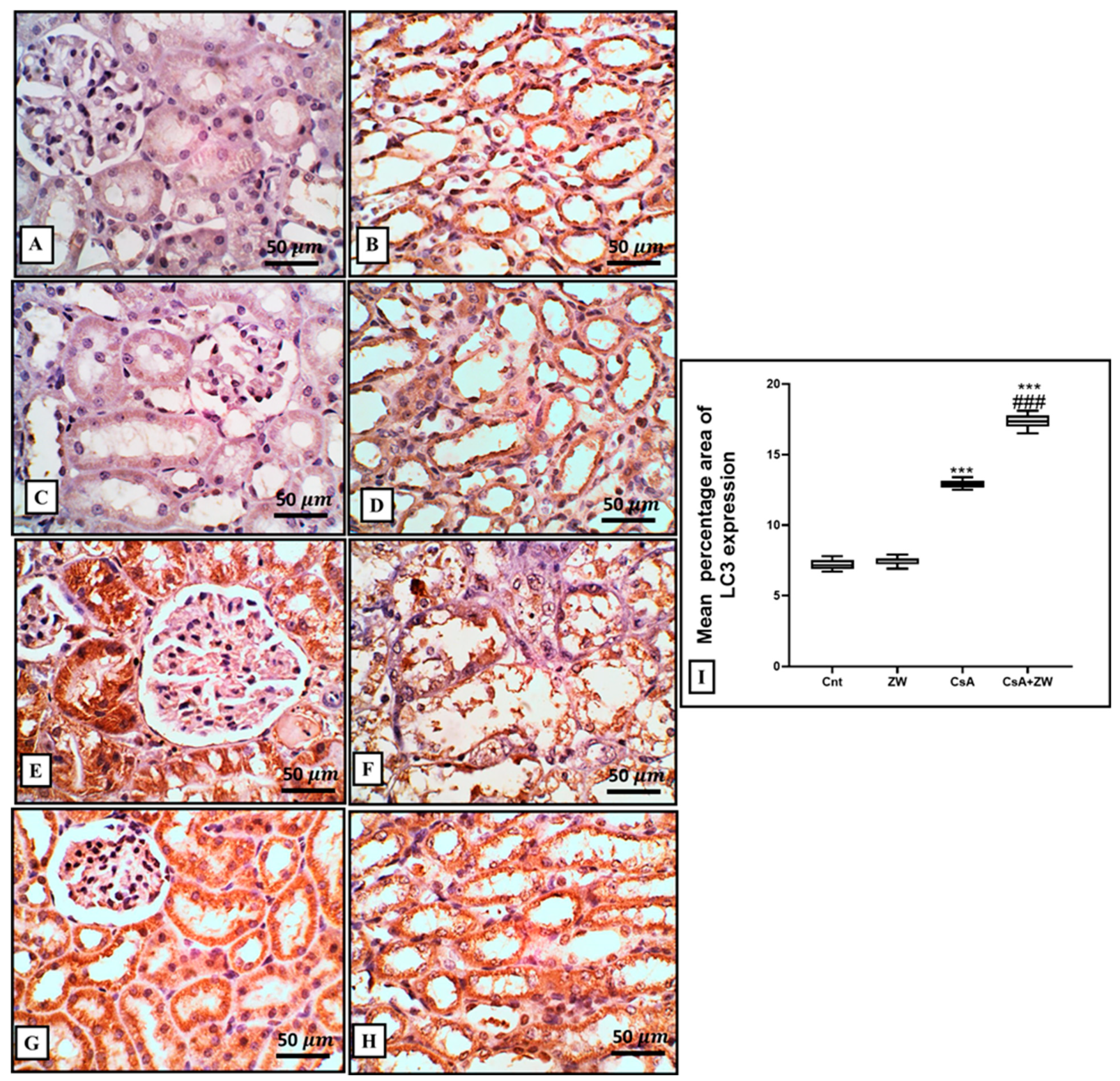
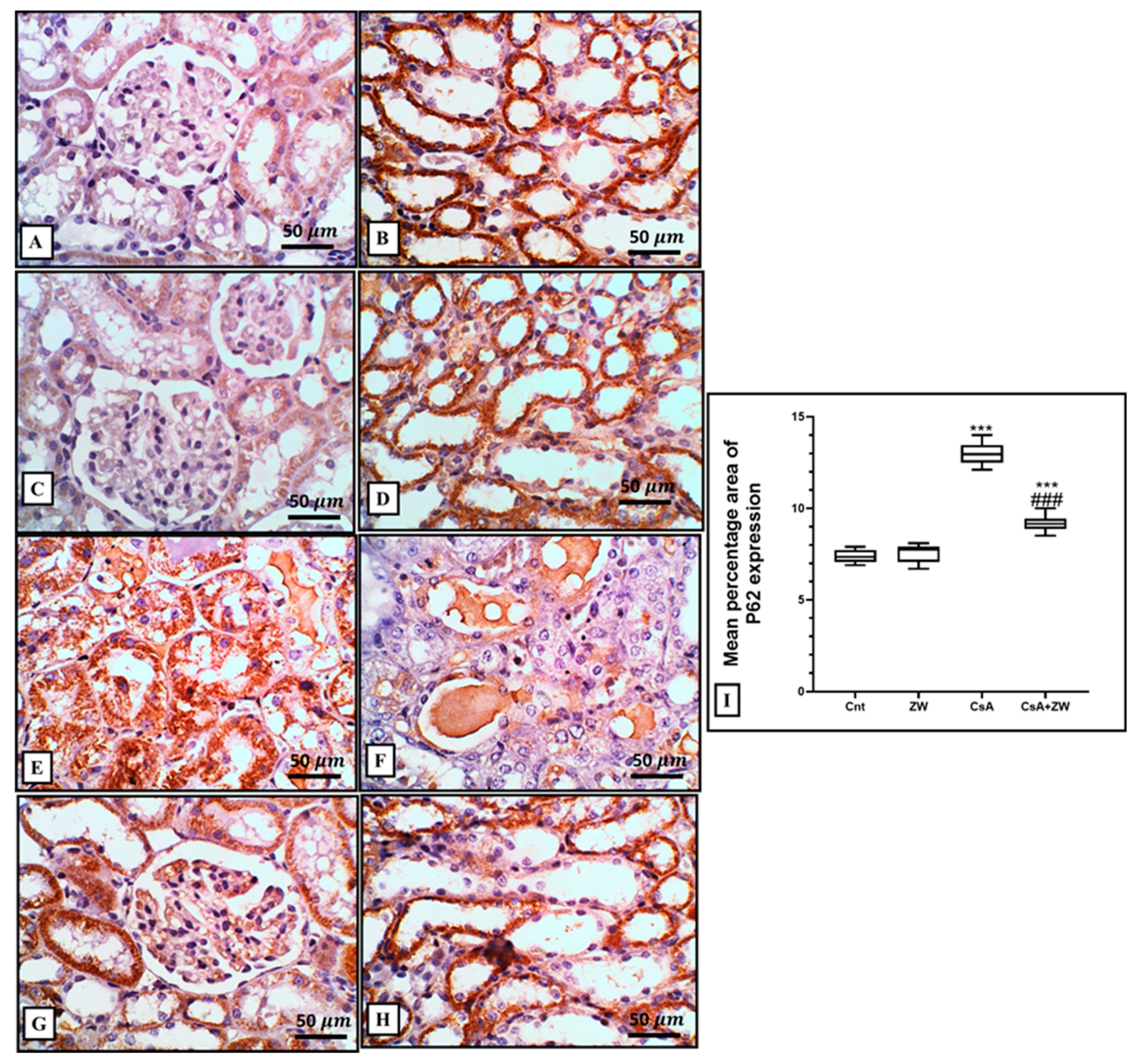
3.5. Effect of Zamzam Water on Autophagic Magnitude in the Kidney Epithelial Cells of Rats with CsA-Induced Nephrotoxicity
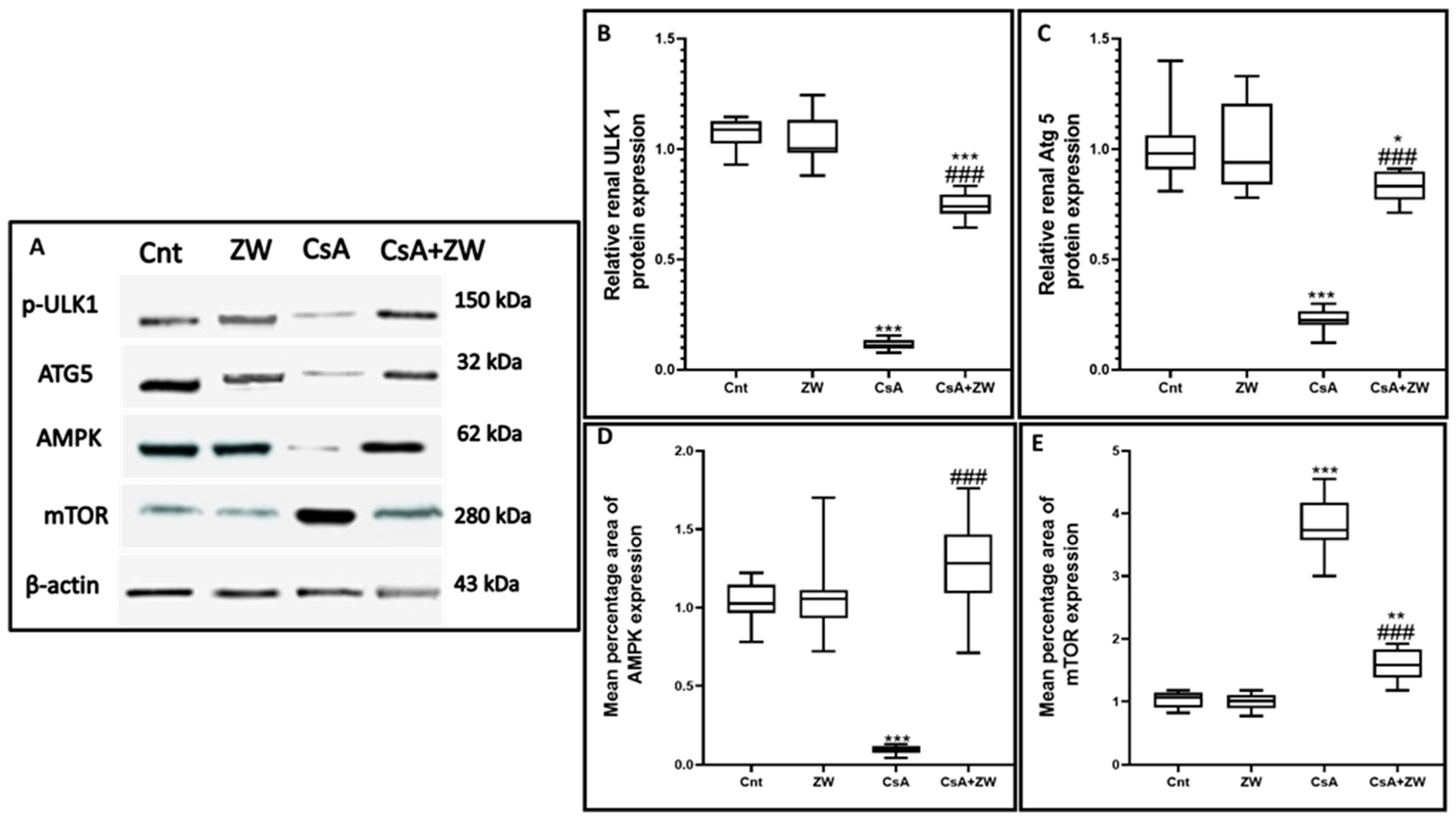
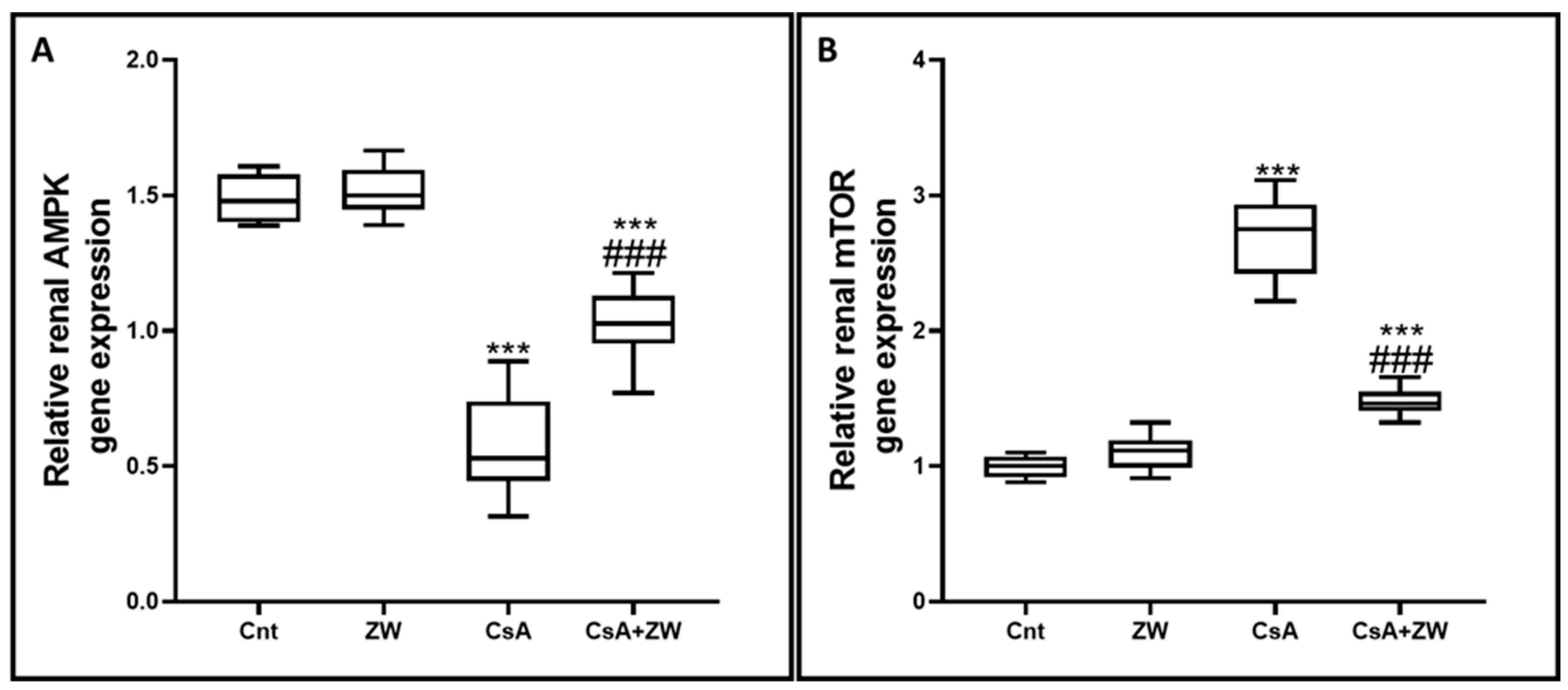
3.6. Effect of Zamzam Water on the AMPK/mTOR Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chighizola, C.B.; Ong, V.H.; Meroni, P.L. The Use of Cyclosporine A in Rheumatology: A 2016 Comprehensive Review. Clin. Rev. Allergy Immunol. 2017, 52, 401–423. [Google Scholar] [CrossRef] [PubMed]
- Periman, L.M.; Mah, F.S.; Karpecki, P.M. A Review of the Mechanism of Action of Cyclosporine A: The Role of Cyclosporine A in Dry Eye Disease and Recent Formulation Developments. Clin. Ophthalmol. 2020, 14, 4187–4200. [Google Scholar] [CrossRef]
- Sommerer, C.; Giese, T. Nuclear Factor of Activated T Cells-Regulated Gene Expression as Predictive Biomarker of Personal Response to Calcineurin Inhibitors. Ther. Drug Monit. 2016, 38 (Suppl. 1), S50–S56. [Google Scholar] [CrossRef]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Zhang, X.; Gu, X.; Li, X.; Shang, L. Progress in understanding the role of lncRNA in programmed cell death. Cell Death Discov. 2021, 7, 30. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Z.; Huang, Y.; Bai, C.; Zhang, X.; Fang, M.; Ju, Z.; Liu, B. Membrane dynamics of ATG4B and LC3 in autophagosome formation. J. Mol. Cell Biol. 2022, 13, 853–863. [Google Scholar] [CrossRef]
- Gupta, R.; Ambasta, R.K.; Kumar, P. Autophagy and apoptosis cascade: Which is more prominent in neuronal death? Cell. Mol. Life Sci. 2021, 78, 8001–8047. [Google Scholar] [CrossRef] [PubMed]
- Kasprowska-Liśkiewicz, D. The cell on the edge of life and death: Crosstalk between autophagy and apoptosis. Postepy Hig. Med. Dosw. (Online) 2017, 71, 825–841. [Google Scholar] [CrossRef]
- Fu, Y.; Xiang, Y.; Wu, W.; Cai, J.; Tang, C.; Dong, Z. Persistent Activation of Autophagy after Cisplatin Nephrotoxicity Promotes Renal Fibrosis and Chronic Kidney Disease. Front. Pharmacol. 2022, 13, 918732. [Google Scholar] [CrossRef] [PubMed]
- Pallet, N.; Bouvier, N.; Bendjallabah, A.; Rabant, M.; Flinois, J.P.; Hertig, A.; Legendre, C.; Beaune, P.; Thervet, E.; Anglicheau, D. Cyclosporine-induced endoplasmic reticulum stress triggers tubular phenotypic changes and death. Am. J. Transplant. 2008, 8, 2283–2296. [Google Scholar] [CrossRef]
- Pallet, N.; Bouvier, N.; Legendre, C.; Gilleron, J.; Codogno, P.; Beaune, P.; Thervet, E.; Anglicheau, D. Autophagy protects renal tubular cells against cyclosporine toxicity. Autophagy 2008, 4, 783–791. [Google Scholar] [CrossRef]
- Ciechomska, I.A.; Gabrusiewicz, K.; Szczepankiewicz, A.A.; Kaminska, B. Endoplasmic reticulum stress triggers autophagy in malignant glioma cells undergoing cyclosporine a-induced cell death. Oncogene 2013, 32, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.; Kim, J. Cyclosporin A aggravates hydrogen peroxide-induced cell death in kidney proximal tubule epithelial cells. Anat. Cell Biol. 2019, 52, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Han, S.W.; Li, C.; Ahn, K.O.; Lim, S.W.; Song, H.G.; Jang, Y.S.; Cho, Y.M.; Jang, Y.M.; Ghee, J.Y.; Kim, J.Y.; et al. Prolonged endoplasmic reticulum stress induces apoptotic cell death in an experimental model of chronic cyclosporine nephropathy. Am. J. Nephrol. 2008, 28, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Al-Barakah, F.N.; Al-jassas, A.M.; Aly, A.A. Water quality assessment and hydrochemical characterization of Zamzam groundwater, Saudi Arabia. Appl. Water Sci. 2017, 7, 3985–3996. [Google Scholar] [CrossRef]
- Alfadul, S.M.; Khan, M.A. Water quality of bottled water in the kingdom of Saudi Arabia: A comparative study with Riyadh municipal and Zamzam water. J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2011, 46, 151928. [Google Scholar] [CrossRef] [PubMed]
- Shomar, B. Zamzam water: Concentration of trace elements and other characteristics. Chemosphere 2012, 86, 6005. [Google Scholar] [CrossRef]
- Al-Doghaither, H.A.; Al-Ghafari, A.B.; Rahimulddin, S.A.; Al-Zahrani, S.M.; Omar, A.S.; Omar, U.M. Evaluation of the potential anticancer activity of Zamzam water in human colon cancer cell line. Cancer Oncol. Res. 2016, 4, 33–41. [Google Scholar] [CrossRef]
- Omar, U.M.; Al Doghaither, H.A.; Rahimulddin, S.A.; Al Zahrani, S.M.; Al-Ghafari, A.B. In Vitro Cytotoxic and Anticancer Effects of Zamzam Water in Human Lung Cancer (A594) Cell Line. Malays. J. Med. Sci. 2017, 24, 15–25. [Google Scholar] [CrossRef]
- Ateyya, H. Amelioration of cyclosporine induced nephrotoxicity by dipeptidyl peptidase inhibitor vildagliptin. Int. Immunopharmacol. 2015, 28, 571–577. [Google Scholar] [CrossRef]
- El-Sheikh, A.A.K.; Morsy, M.A.; Abdel-Latif, R.G. Modulation of eNOS/iNOS by nebivolol protects against cyclosporine A-mediated nephrotoxicity through targeting inflammatory and apoptotic pathways. Environ. Toxicol. Pharmacol. 2019, 69, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Abdel-latif, R.G.; Morsy, M.A.; El-Moselhy, M.A.; Khalifa, M.A. Sildenafil protects against nitric oxide deficiency-related nephrotoxicity in cyclosporine A treated rats. Eur. J. Pharmacol. 2013, 705, 126–134. [Google Scholar] [CrossRef]
- Mohamadin, A.M.; El-Beshbishy, H.A.; El-Mahdy, M.A. Green tea extract attenuates cyclosporine A-induced oxidative stress in rats. Pharmacol. Res. 2005, 51, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mraisel, A.; Abu Ali, A. Protective effect of Zamzam water against kidneys damage induced in male rats: Immunohistochemistry evidence. J. Biosci. Appl. Res. 2017, 3, 42–47. [Google Scholar] [CrossRef]
- Kumar, H.; Kumar, A.; Kumari, P.; Tulsani, N.B. A test strip for the estimation of urea in serum. Indian J. Clin. Biochem. 2000, 15, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Piéroni, L.; Delanaye, P.; Boutten, A.; Bargnoux, A.S.; Rozet, E.; Delatour, V.; Carlier, M.C.; Hanser, A.M.; Cavalier, E.; Froissart, M.; et al. Société Française de Biologie Clinique. A multicentric evaluation of IDMS-traceable creatinine enzymatic assays. Clin. Chim. Acta 2011, 412, 2070–2075. [Google Scholar] [CrossRef]
- Mesbah, L.; Soraya, B.; Narimane, S.; Jean, P.F. Protective effect of flavonides against the toxicity of vinblastine cyclophosphamide and paracetamol by inhibition of lipid-peroxydation and increase of liver glutathione. Hematology 2004, 7, 59–67. [Google Scholar]
- Xu, C.; Liu, S.; Liu, Z.; Song, F.; Liu, S. Superoxide generated by pyrogallol reduces highly water-soluble tetrazolium salt to produce a soluble formazan: A simple assay for measuring superoxide anion radical scavenging activities of biological and abiological samples. Anal. Chim. Acta 2013, 793, 53–60. [Google Scholar] [CrossRef]
- Zhang, Z.; Miyatake, S.; Saiki, M.; Asahi, M.; Yukawa, H.; Toda, H.; Kikuchi, H.; Yoshimura, S.I.; Hashimoto, N. Selenium and glutathione peroxidase mRNA in rat glioma. Biol. Trace Elem. Res. 2000, 73, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Baureder, M.; Barane, E.; Hederstedt, L. In vitro assembly of catalase. J. Biol. Chem. 2014, 289, 28411–28420. [Google Scholar] [CrossRef]
- Zahran, R.; Ghozy, A.; Elkholy, S.S.; El-Taweel, F.; El-Magd, M.A. Combination therapy with melatonin, stem cells and extracellular vesicles is effective in limiting renal ischemia-reperfusion injury in a rat model. Int. J. Urol. 2020, 27, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Naim, A.B.; Neamatallah, T.; Eid, B.G.; Esmat, A.; Alamoudi, A.J.; Abd El-Aziz, G.S.; Ashour, O.M. 2-Methoxyestradiol attenuates testosterone-induced benign prostate hyperplasia in rats through inhibition of HIF-1_/TGF- _/Smad2 Axis. Oxid. Med. Cell. Longev. 2018, 2018, 4389484. [Google Scholar] [CrossRef]
- Liu, S.; Lutz, J.; Chang, J.; Liu, D.; Heemann, U.; Baumann, M. TRAF6 knockdown promotes survival and inhibits inflammatory response to lipopolysaccharides in rat primary renal proximal tubule cells. Acta Physiol. 2010, 199, 339–346. [Google Scholar] [CrossRef]
- Kaya, E.; Ozer Kaya, S.; Yilmaz, S.; Ceribasi, A.O.; Turk, G. Evaluation of ameliorating effect of lycopene against testicular toxicity due to diethylnitrosamine using biochemical, spermatological and histopathological data. Andrologia 2019, 51, e13274. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, D.; Haragsim, L. Cyclosporine: A review. J. Transplant. 2012, 2012, 230386. [Google Scholar] [CrossRef]
- Ghazipour, A.M.; Shirpoor, A.; Ghiasi, R.; Pourheydar, B.; Khalaji, N.; Naderi, R. Cyclosporine A induces testicular injury via mitochondrial apoptotic pathway by regulation of mir-34a and sirt-1 in male rats: The rescue effect of curcumin. Chem. Biol. Interact. 2020, 327, 109180. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Cosemi, E.; Kamel, S.; Mohammed, S.; Elhefnawy, M.; Farid, L.; Shaker, S. Oncolytic action of Zamzam water on azoxyonethone (aom) induced colon tumors in rats. In Proceedings of the Thirteenth International Water Technology Conference, IWTC 13, Hurghada, Egypt, 12–15 March 2009. [Google Scholar]
- Saif, A.; Sarhan, O.; Elmogy, M.; Mutwally, D.H. Hepatoprotective effects of Zamzam water against carbon tetrachloride induced liver damage in rats: Biochemical, histopathological, and molecular evidences. Life Sci. J. 2014, 11, 300–308. [Google Scholar]
- Almalki, W.; Arafa, E.S.; Abdallah, A.; Mahfoz, A.M.; Osman, A.; Abd El-Latif, H.; Shahid, I. Zinc chloride protects against streptozotocin-induced diabetic nephropathy in rats. Pharmacol. Pharm. 2016, 07, 331–342. [Google Scholar] [CrossRef]
- Momeni, M.; Gharedaghi, Z.; Amin, M.M.; Poursafa, P.; Mansourian, M. Does water hardness have preventive effect on cardiovascular disease? Int. J. Prev. Med. 2014, 5, 159–163. [Google Scholar]
- Haleagrahara, N.; Yee, T.; Chakravarthi, S.; Lee, N. Protective effect of n-acetylcysteine on cyclosporine a-induced changes in lipid hydroperoxide levels and renal dysfunction in rats. Arch. Med. Sci. 2009, 5, 16–22. [Google Scholar]
- Helmy, M.M.; Helmy, M.W.; El-Mas, M.M. Upregulation of cystathionine-γ-lyase/hydrogen sulfide pathway underlies the celecoxib counteraction of cyclosporine-induced hypertension and renal insult in rats. Prostaglandins Other Lipid Mediat. 2019, 141, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ciarcia, R.; Damiano, S.; Florio, A.; Spagnuolo, M.; Zacchia, E.; Squillacioti, C.; Mirabella, N.; Florio, S.; Pagnini, U.; Garofano, T.; et al. The Protective Effect of Apocynin on Cyclosporine A-Induced Hypertension and Nephrotoxicity in Rats. J. Cell. Biochem. 2015, 116, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Yang, J.W.; Han, B.G.; Kwon, H.J.; Kim, J.H.; Choi, S.O. Protective Role of Apelin against Cyclosporine-Induced Renal Tubular Injury in Rats. Transplant. Proc. 2017, 49, 1499–1509. [Google Scholar] [CrossRef]
- Yoon, S.P. Insights into the possible mechanism of cyclosporine-induced chronic nephrotoxicity; arteriolopathy. Nephrourol. Mon. 2012, 4, 489–490. [Google Scholar] [CrossRef]
- Lee, J. Use of antioxidants to prevent cyclosporine a toxicity. Toxicol. Res. 2010, 26, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Capasso, G.; Di Gennaro, C.I.; Della Ragione, F.; Manna, C.; Ciarcia, R.; Florio, S.; Perna, A.; Pollastro, R.M.; Damiano, S.; Mazzoni, O.; et al. In vivo effect of the natural antioxidant hydroxytyrosol on cyclosporine nephrotoxicity in rats. Nephrol. Dial. Transplant. 2008, 23, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Abdelhadya, D.H.; El-Magd, M.A.; Elbialy, Z.I.; Saleh, A.A. Bromuconazole-induced hepatotoxicity is accompanied by upregulation of PXR/CYP3A1 and downregulation of CAR/CYP2B1 gene expression. Toxicol. Mech. Methods 2017, 27, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Abu Gazia, M.; El-Magd, M.A. Effect of pristine and functionalized multiwalled carbon nanotubes on rat renal cortex. Acta Histochem. 2019, 121, 207–217. [Google Scholar] [CrossRef]
- El-Magd, M.A.; Kahilo, K.A.; Nasr, N.E.; Kamal, T.; Shukry, M.; Saleh, A.A. A potential mechanism associated with lead-induced testicular toxicity in rats. Andrologia 2017, 49, e12750. [Google Scholar] [CrossRef]
- Singh, A.; Ramnath, R.D.; Foster, R.R.; Wylie, E.C.; Fridén, V.; Dasgupta, I.; Haraldsson, B.; Welsh, G.I.; Mathieson, P.W.; Satchell, S.C. Reactive oxygen species modulate the barrier function of the human glomerular endothelial glycocalyx. PLoS ONE 2013, 8, e55852. [Google Scholar] [CrossRef]
- Cao, Y.; Roursgaard, M.; Danielsen, P.H.; Møller, P.; Loft, S. Carbon black nanoparticles promote endothelial activation and lipid accumulation in macrophages independently of intracellular ROS production. PLoS ONE 2014, 9, e106711. [Google Scholar] [CrossRef]
- El Maleky, W.; Mahfoz, A.M.; Osman, A.O.; Abd El-Latif, H.A. Investigation of the impacts of Zamzam water on streptozotocin-induced diabetic nephropathy in rats. In-vivo and in-vitro study. Biomed. Pharmacother. 2021, 138, 111474. [Google Scholar] [CrossRef]
- Abdullah, A.M.; Abdelsalam, E.; Abdullah, B.; Khaled, A. Antioxidant effects of Zamzam water in normal rats and those under induced-oxidative stress. J. Med. Plants Res. 2012, 6, 5507–5512. [Google Scholar]
- Papachristoforou, E.; Lambadiari, V.; Maratou, E.; Makrilakis, K. Association of Glycemic Indices (Hyperglycemia, Glucose Variability, and Hypoglycemia) with Oxidative Stress and Diabetic Complications. J. Diabetes Res. 2020, 2020, 7489795. [Google Scholar] [CrossRef] [PubMed]
- Mahfoz, A.M.; El-Latif, H.A.; Ahmed, L.A.; Hassanein, N.M.; Shoka, A.A. Anti-diabetic and renoprotective effects of aliskiren in streptozotocin-induced diabetic nephropathy in female rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2016, 389, 1315–1324. [Google Scholar] [CrossRef]
- Satta, H.; Iwamoto, T.; Kawai, Y.; Koguchi, N.; Shibata, K.; Kobayashi, N.; Yoshida, M.; Nakayama, M. Amelioration of hemodialysis-induced oxidative stress and fatigue with a hemodialysis system employing electrolyzed water containing molecular hydrogen. Ren. Replace Ther. 2021, 7, 37. [Google Scholar] [CrossRef]
- Bamosa, A.; Elnour, A.; Kaatabi, H.; Al Meheithif, A.; Aleissa, K.; Al-Almaie, S. Zamzam water ameliorates oxidative stress and reduces hemoglobinA1c in Type 2 diabetic patients. J. Diabetes Metab. 2013, 4, 249. [Google Scholar] [CrossRef]
- Rao, S.R.; Sundararajan, S.; Subbarayan, R.; Murugan Girija, D. Cyclosporine-A induces endoplasmic reticulum stress and influences pro-apoptotic factors in human gingival fibroblasts. Mol. Cell. Biochem. 2017, 429, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.B.; Santamaría, B.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A. Mechanisms of renal apoptosis in health and disease. J. Am. Soc. Nephrol. 2008, 19, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ren, B.; Wang, Y.; Zou, C.; Qiao, Q.; Diao, Z.; Mi, Y.; Zhu, D.; Liu, X. Sesamol Induces Human Hepatocellular Carcinoma Cells Apoptosis by Impairing Mitochondrial Function and Suppressing Autophagy. Sci. Rep. 2017, 7, 45728. [Google Scholar] [CrossRef] [PubMed]
- Koppelstaetter, C.; Kern, G.; Leierer, G.; Mair, S.M.; Mayer, G.; Leierer, J. Effect of cyclosporine, tacrolimus and sirolimus on cellular senescence in renal epithelial cells. Toxicol. In Vitro 2018, 48, 86–92. [Google Scholar] [CrossRef]
- Tu, H.P.; Chen, Y.T.; Chiu, H.C.; Chin, Y.T.; Huang, S.M.; Cheng, L.C.; Fu, E.; Chiang, C.Y. Cyclosporine A enhances apoptosis in gingival keratinocytes of rats and in OECM1 cells via the mitochondrial pathway. J. Periodontal. Res. 2009, 44, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Øzbay, L.A.; Smidt, K.; Mortensen, D.M.; Carstens, J.; Jørgensen, K.A.; Rungby, J. Cyclosporin and tacrolimus impair insulin secretion and transcriptional regulation in INS-1E beta-cells. Br. J. Pharmacol. 2011, 162, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Rosa, M.D.; Distefano, G.; Gagliano, C.; Rusciano, D.; Malaguarnera, L. Autophagy in Diabetic Retinopathy. Curr. Neuropharmacol. 2016, 14, 810–825. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Choi, S.I.; Jeung, E.B.; Yoo, Y.M. Cyclosporine A induces apoptotic and autophagic cell death in rat pituitary GH3 cells. PLoS ONE 2014, 9, e108981. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Y.; Zou, H.; Wang, B.; Sun, Q.; Fu, A.; Wang, Y.; Wang, Y.; Xu, X.; Li, W. Probiotic Bacillus amyloliquefaciens SC06 Induces Autophagy to Protect against Pathogens in Macrophages. Front. Microbiol. 2017, 8, 469. [Google Scholar] [CrossRef]
- Inoue, K.; Kuwana, H.; Shimamura, Y.; Ogata, K.; Taniguchi, Y.; Kagawa, T.; Horino, T.; Takao, T.; Morita, T.; Sasaki, S.; et al. Cisplatin-induced macroautophagy occurs prior to apoptosis in proximal tubules in vivo. Clin. Exp. Nephrol. 2010, 14, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wei, Q.; Dong, G.; Komatsu, M.; Su, Y.; Dong, Z. Autophagy in proximal tubules protects against acute kidney injury. Kidney Int. 2012, 82, 1271–1283. [Google Scholar] [CrossRef]
- Li, J.; Xu, Z.; Jiang, L.; Mao, J.; Zeng, Z.; Fang, L.; He, W.; Yuan, W.; Yang, J.; Dai, C. Rictor/mTORC2 protects against cisplatin-induced tubular cell death and acute kidney injury. Kidney Int. 2014, 86, 86–102. [Google Scholar] [CrossRef]
- Ozkok, A.; Edelstein, C.L. Pathophysiology of cisplatin-induced acute kidney injury. Biomed. Res. Int. 2014, 2014, 967826. [Google Scholar] [CrossRef]
- Park, H.G.; Yi, H.; Kim, S.H.; Yu, H.S.; Ahn, Y.M.; Lee, Y.H.; Roh, M.S.; Kim, Y.S. The effect of cyclosporine A on the phosphorylation of the AMPK pathway in the rat hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 1933–1937. [Google Scholar] [CrossRef]
- Zhang, D.; Pan, J.; Xiang, X.; Liu, Y.; Dong, G.; Livingston, M.J.; Chen, J.K.; Yin, X.M.; Dong, Z. Protein Kinase Cδ Suppresses Autophagy to Induce Kidney Cell Apoptosis in Cisplatin Nephrotoxicity. J. Am. Soc. Nephrol. 2017, 28, 11311144. [Google Scholar] [CrossRef]
- Li, H.; Tang, Y.; Wen, L.; Kong, X.; Chen, X.; Liu, P.; Zhou, Z.; Chen, W.; Xiao, C.; Xiao, P.; et al. Neferine reduces cisplatin-induced nephrotoxicity by enhancing autophagy via the AMPK/mTOR signaling pathway. Biochem. Biophys. Res. Commun. 2017, 484, 694–701. [Google Scholar] [CrossRef]
- Pyrzynska, B.; Serrano, M.; Martínez, A.C.; Kaminska, B. Tumor suppressor p53 mediates apoptotic cell death triggered by cyclosporin A. J. Biol. Chem. 2002, 277, 14102–14108. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Gallego, R.; Mund, M.; Cosentino, K.; Schneider, J.; Unsay, J.; Schraermeyer, U.; Engelhardt, J.; Ries, J.; García-Sáez, A.J. Bax assembly into rings and arcs in apoptotic mitochondria is linked to membrane pores. EMBO J. 2016, 35, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Komatsu, M. Impaired G1-arrest, autophagy, and apoptosis in Atg7-knockout mice. Circ. Res. 2012, 111, 962–964. [Google Scholar] [CrossRef] [PubMed]
- Qiang, W.; Shen, T.; Noman, M.; Guo, J.; Jin, Z.; Lin, D.; Pan, J.; Lu, H.; Li, X.; Gong, F. Fibroblast Growth Factor 21 Augments Autophagy and Reduces Apoptosis in Damaged Liver to Improve Tissue Regeneration in Zebrafish. Front. Cell Dev. Biol. 2021, 9, 756743. [Google Scholar] [CrossRef]
- Fung, C.; Lock, R.; Gao, S.; Salas, E.; Debnath, J. Induction of autophagy during extracellular matrix detachment promotes cell survival. Mol. Biol. Cell 2008, 19, 797–806. [Google Scholar] [CrossRef]
- Vessoni, A.T.; Filippi-Chiela, E.C.; Menck, C.F.; Lenz, G. Autophagy and genomic integrity. Cell Death Differ. 2013, 20, 1444–1454. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) | Gene Accession Number | |
|---|---|---|---|
| AMPK | AGCTCGCAGTGGCTTATCAT | GGGGCTGTCTGCTATGAGAG | NM_023991.1 |
| mTOR | ACGAAGGAGACAGACCGAAG | CGACGAAGTCACTAGATTCA | AM_943028.1 |
| GAPDH | CACCCTGTTGCTGTAGCCATATTC | GACATCAAGAAGGTGGTGAAGCAG | XM_017592435.1 |
| Groups | Serum Creatinine (mg/dL) | Blood Urea Nitrogen (mg/dL) | Creatinine Clearance Level (mL/min) |
|---|---|---|---|
| Control | 0.68 ± 0.07 | 21.60 ± 4.60 | 0.248 ± 0.016 |
| ZW | 0.66 ± 0.08 | 22.80 ± 5.20 | 0.2520.018 |
| CsA | 1.99 ± 0.19 *** | 90.40 ±7.15 *** | 0.015 ± 0.003 *** |
| CsA + Zamzam water | 0.94 ± 0.12 **,### | 34.40 ± 4.42 ***,### | 0.098 ± 0.021 ***,### |
| Classes | MDA (nmol/g Tissue) | SOD (U/g/Tissue) | CAT (U/g/Tissue) | GPx (U/g/Tissue) |
|---|---|---|---|---|
| control | 11.33 ± 1.04 | 3.99 ± 0.19 | 2.20 ± 0.16 | 125 ±7.91 |
| ZW | 11.70 ± 0.93 | 4.15 ± 0.22 | 2.25 ± 0.15 | 128 ± 7.07 |
| CsA | 29.81 ± 3.41 *** | 1.00 ± 0.13 *** | 0.84 ± 0.09 *** | 36.28 ± 5.67 *** |
| CsA+Zam | 17.65 ± 1.83 ***,### | 2.13 ± 0.38 ***,### | 1.26 ± 0.22 ***,### | 89.97 ± 7.14 ***,### |
| Sample | ZW | Tap Water |
|---|---|---|
| Total hardness | 670 mg/L | 88 mg/L |
| Calcium hardness | 422 mg/L | 134 mg/L |
| Total alkalinity | 302 mg/L | 44 mg/L |
| Magnesium hardness | 211 mg/L | 77 mg/L |
| Calcium | 189 mg/L | 52 mg/L |
| Magnesium | 520 mg/L | 74 mg/L |
| Potassium | 121 mg/L | 12 mg/L |
| PH | 7.8 | 7 |
| Nitrite | 0.01 mg/L | 0.001 mg/L |
| Nitrates | 173 mg/L | 13 mg/L |
| Chlorine | 343 mg/L | 99 mg/L |
| Phosphates | 0.27 mg/L | 0.002 mg/L |
| Bicarbonate | 364 mg/L | 110 mg/L |
| Sulfites | 370 mg/L | 210 mg/L |
| Zinc | 944 mg/L | 186 mg/L |
| Selenium | 320 mg/L | 69 mg/L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taha, M.; Elazab, S.T.; Baokbah, T.A.S.; Al-Kushi, A.G.; Mahmoud, M.E.; Abdelbagi, O.; Qusty, N.F.; El-Shenbaby, I.; Babateen, O.; Badawy, A.M.; et al. Palliative Role of Zamzam Water against Cyclosporine-Induced Nephrotoxicity through Modulating Autophagy and Apoptosis Crosstalk. Toxics 2023, 11, 377. https://doi.org/10.3390/toxics11040377
Taha M, Elazab ST, Baokbah TAS, Al-Kushi AG, Mahmoud ME, Abdelbagi O, Qusty NF, El-Shenbaby I, Babateen O, Badawy AM, et al. Palliative Role of Zamzam Water against Cyclosporine-Induced Nephrotoxicity through Modulating Autophagy and Apoptosis Crosstalk. Toxics. 2023; 11(4):377. https://doi.org/10.3390/toxics11040377
Chicago/Turabian StyleTaha, Medhat, Sara T. Elazab, Tourki A. S. Baokbah, Abdullah G. Al-Kushi, Mohamed Ezzat Mahmoud, Omer Abdelbagi, Naeem F. Qusty, Ibrahim El-Shenbaby, Omar Babateen, Alaa. M. Badawy, and et al. 2023. "Palliative Role of Zamzam Water against Cyclosporine-Induced Nephrotoxicity through Modulating Autophagy and Apoptosis Crosstalk" Toxics 11, no. 4: 377. https://doi.org/10.3390/toxics11040377