1. Introduction
Mercury pollution in water bodies due to human activities is mainly caused by wastewater from industries that produce items such as chlor-alkali, plastics, batteries, electronics, and used medical devices [
1]. It is estimated that approximately 3.2 × 10
4 hm of agricultural land in China is contaminated with mercury. Mercury is highly toxic and can cause severe damage to human cells and limit normal cellular function when it enters the human body [
2]. Mercury is distributed in the atmosphere, soil, and water bodies; both soluble and insoluble mercury compounds are partially volatilized into the atmosphere, and the transport transformation of mercury occurs between land, water, and air. A general rule of thumb is that, compared to inorganic mercury, organic mercury is more volatile [
1], and methylmercury is the most volatile. Through microorganisms, inorganic mercury can be converted into more toxic methylmercury, dimethylmercury [
3], etc.
Numerous studies have examined the possibility of removing mercury with microbial organisms. For example, Yang Wen screened a mercury-resistant strain in a mercury-rich soil upstream of Miyun Reservoir in Beijing, and this strain was identified as
Pseudomonas aeruginosa [
4]. When mercury concentrations were 10 mg/L for 24 h under optimum conditions, the rates of mercury removal were over 90%. Zheng Yan et al. reported a highly resistant strain of mercury,
Pseudomonas aeruginosa, which can rapidly volatilize more than 68% of Hg
0 within 8 h, and Fan Taotao et al. isolated three strains of mercury-resistant and mercury-reducing bacteria from mercury-contaminated water in a chemical plant [
5], and these strains exhibit a mercury removal rate of more than 90% under optimal conditions. The mercury-tolerant strains in this study were selected from agricultural fields that were contaminated with heavy metals near molybdenum mines in Luanchuan County, and the rate of mercury removal was approximately 96% at mercury concentrations of 2–12 mg/L, a higher percentage than that previously reported in the literature.
Heavy metal-contaminated soil located around mining areas is a valuable source for isolating strains tolerant to heavy metals [
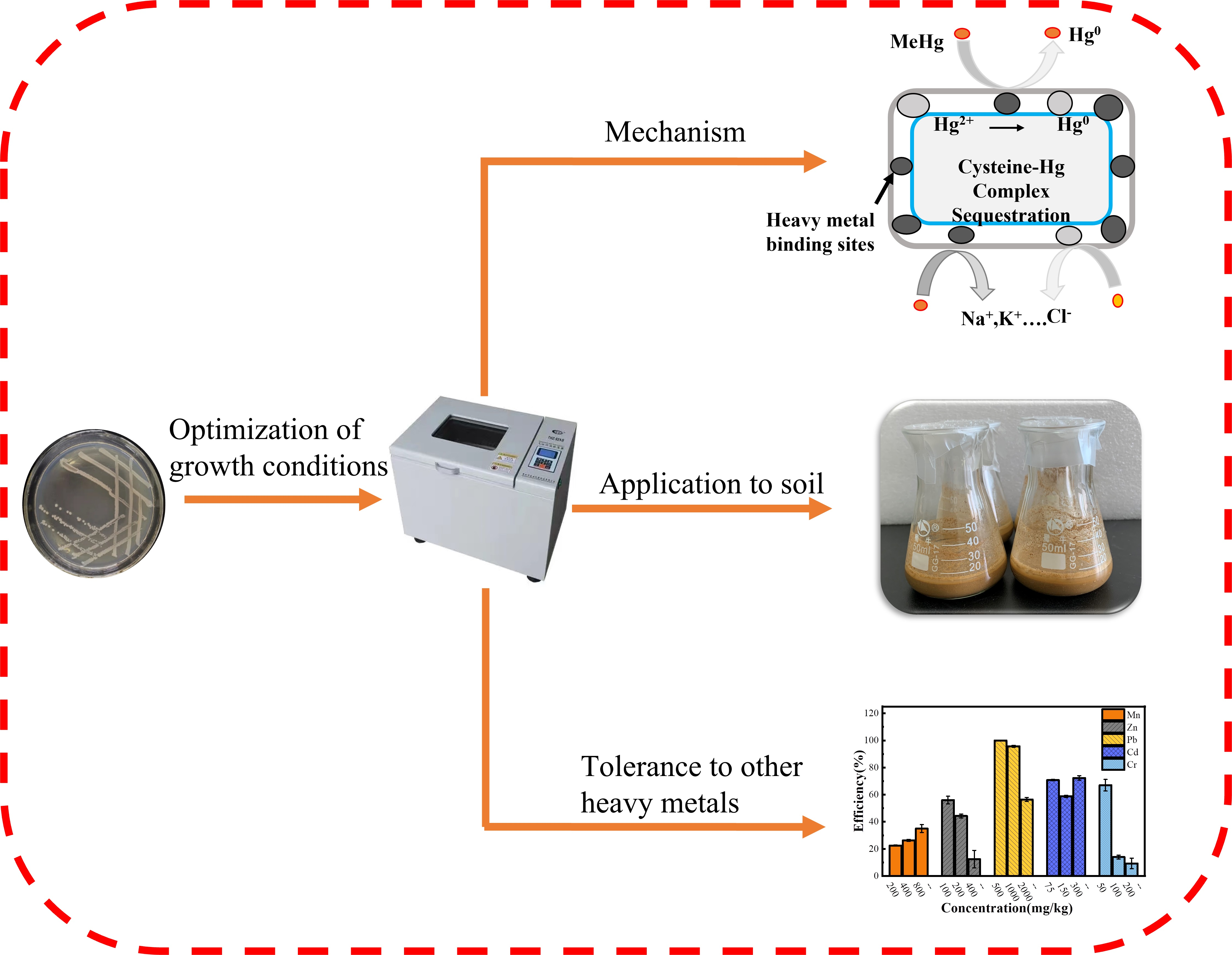
6]. A strain with mercury tolerance was isolated from agricultural fields in the molybdenum mining area of Luanchuan County, Luoyang. Its biological properties and volatile mercury energy were initially investigated. The possibility of using this strain to remediate mercury-contaminated environments was evaluated to provide a reference for future microbial remediation of mercury-contaminated soils in mining areas.
2. Materials and Methods
2.1. Soil Sample Source
In this experiment, mercury-contaminated soils were collected from agricultural fields in Luanchuan County, Luoyang City, Henan Province (33° N, 111° E) using the five-point sampling method, and the samples were primarily from the protoplasmic soil layer within the top 20 cm of the soil. The mercury content of the soil was determined by atomic fluorescence with a detection limit of 0.002 mg/kg.
2.2. Screening Mercury-Resistant Strains
Soil samples were collected from the heavy metal-contaminated soil in the mining area; 10 g of soil was placed in a 250 mL beaker, 90 mL of sterile water was added to create a bacterial suspension and the suspension was magnetically stirred for 30 min and left to stand. Then, 1 mL of supernatant was inoculated with a pipette gun in an LB medium containing 10 mg/L Hg
2+ in a 30 °C; the samples were shaken at 150 r/min in an incubator and the samples were incubated for 2 h for 10× series (10
−2, 10
−3, 10
−4 and 10
−5) gradient dilutions [
1]. A dilution of the sample was spread on a solid LB medium containing 20 mg/L Hg
2+ and incubated in a biochemical incubator at 37 °C for 48 h. After the colonies grew on the medium, the concentration of mercuric chloride in the LB solid medium was increased continuously. A high concentration of mercury-tolerant strains was screened by picking and culturing individual colonies on an LB solid medium containing a high mercury concentration.
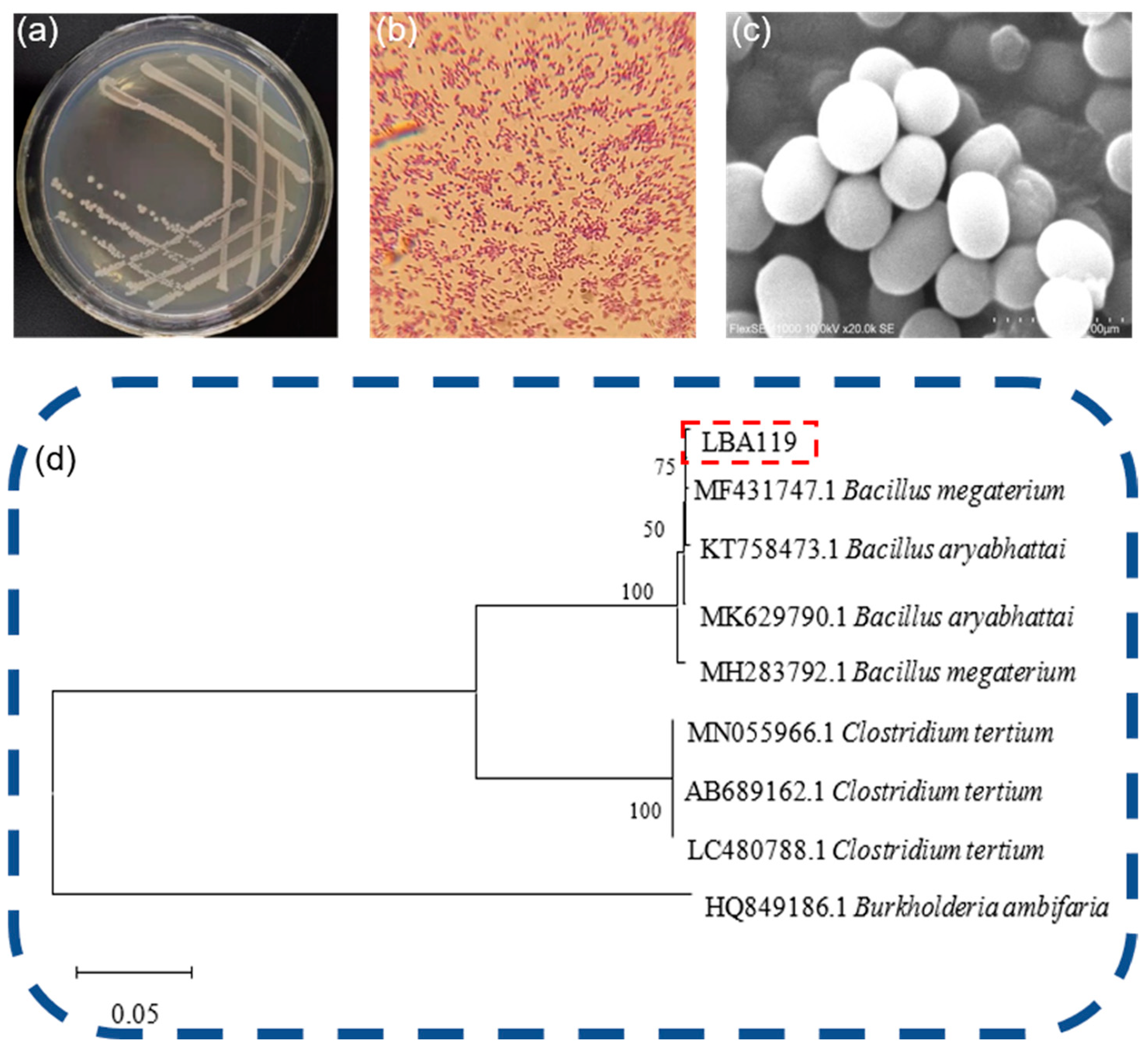
The selected mercury-resistant strains were cultured in line on LB solid medium without mercury, and the strains were repeatedly purified and screened. The morphological characteristics of the colonies on the medium were observed and recorded. Inoculations of the mercury-resistant strain were placed into LB liquid medium and were incubated at 30 °C and 150 rpm in a constant temperature shaker until the solution became cloudy. The solution was preserved using glycerol. With the aid of a pipette on an ultraclean table, 0.2 mL of glycerol was pipetted into a 1.5 mL centrifuge tube, and 0.2 mL of the bacterial seed solution was mixed with gentle shaking. The 1.5 mL centrifuge tube containing the bacterial solution was placed in a sealed bag and placed in a refrigerator at −40 °C for use in subsequent experiments.
2.3. Strain Identification
The bacteria were identified by colony morphology and biochemical tests. Biochemical tests included Gram stain [
7], the starch hydrolysis test, gelatin hydrolysis test, oil hydrolysis test, indole test, sugar fermentation test, methyl red test, volt-pop (VP) test, H
2S test, and the strain motility test.
2.4. Molecular Biology Identification
Phylogenetic trees are used to represent the affinities between various organisms and to infer the evolutionary history of species through the study of biological sequences, mainly through DNA sequences, protein sequences, protein structures, etc. Phylogenetic trees are constructed. The phylogenetic tree in this study was constructed based on the 16S rDNA of LBA119 bacteria and was compared with the validated 16S rDNA in the NCBI library.
Genomic DNA of the strain was extracted by the boiling template method, and the 16S rDNA gene [
8] was amplified by polymerase chain reaction (PCR) using bacterial universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-ACGGCTACCTTGTTACGA-3′). The temperature program was to preheat at 95 °C for 5 min, denaturation at 95 °C for 1 min, 55 °C annealing for 1 min, and 72 °C extension for 2 min, repeated for 30 cycles, and finally 72 °C extension for 10 min. The composition of the PCR reaction system is shown in
Table 1. The PCR [
9] products were sent to Shanghai Bioengineering Co., Ltd (Sangon Biotech (Shanghai) Co., Ltd., Shanghai, China). for sequencing, and the sequencing results were compared with the GenBank database of the National Center for Biotechnology Information (NCBI) to identify the genus of the strain. The phylogenetic tree was constructed by the neighbor-joining method (distance-based approach) of the MEGA X software (Version 11.0.13).
2.5. Exploration of Optimal Growth Conditions for Bacteria Strains
To investigate the effect of different media on the growth of experimental strains [
10], 50 mL of each of eight liquid media, including an LB medium, beef paste peptone medium, high type 1 medium, PDA medium, Choi medium, sand medium, glycerol medium, and PSA medium, was prepared in a 250 mL conical flask, and a standard liquid medium containing 10 mg/L Hg
2+ and no Hg
2+ was prepared. Inoculation was performed after the medium was adjusted to pH 7.0, sterilized at 121 °C for 30 min, and cooled (fresh bacterial seed solution to medium = 2:100). The culture was incubated for 36 h at 37 °C and 150 r/min in a constant temperature shaking incubator. The cultured bacterial solution was centrifuged at 8000 rpm for 10 min, the supernatant was discarded, and the centrifuge tube was placed in a vacuum drying oven and dried at 70 °C to a constant weight. Each group was divided into three replicates. The dry weight of the strain was calculated as follows:
where M
1 is the initial mass of the experimental group, M
2 is the initial mass of the control group, m
1 is the mass of the experimental group after drying, and m
2 is the mass of the control group after drying.
Under the optimum conditions, LB liquid medium containing 10 mg/L Hg2+ and LB liquid medium without Hg2+ were used. The following range of inoculums was used for the test strains: 1%, 2%, 3%, 4%, 5%, 6%, 7%, and 8%, and the OD600nm values were determined after 36 h of incubation at 150 rpm and 30 °C.
The pH of the liquid LB medium was adjusted to 4, 5, 6, 7, 8, 9, and 10 with hydrochloric acid (HCl) or sodium hydroxide (NaOH), and the OD600nm values were measured after inoculating fresh bacterial solutions into LB liquid medium and incubating the solutions at 37 °C for 36 h at 150 rpm to obtain the proper pH range for strain growth.
Based on the same medium preparation method, pH, and inoculation method, the OD600 nm values were determined after 36 h of incubation [
11] at 20 °C, 25, 30, 35 °C, and 40 °C in a constant temperature shaking incubator. Under the above optimal conditions, the bacteria were inoculated in liquid LB [
12] media containing 0, 0.5%, 1%, 2%, 4%, 6%, and 8% sodium chloride, and the OD
600 nm values were measured after incubation to obtain the optimal salt concentration. Each group of experiments was repeated three times.
2.6. Determination of the Tolerance of Strains to Mercury
Under optimal growth conditions, LB liquid medium was prepared with Hg2+ concentrations of 0, 1, 2, 4, 8, 16, 32, 64, and 128 mg/L, sterilized at 121 °C for 30 min, inoculated with 2% fresh bacterial broth into LB liquid medium, and incubated with shaking for 120 h. To determine the minimum inhibitory concentration of heavy metal mercury on the experimental strains, the optical density (OD600nm) values of each medium were measured regularly. The minimum inhibitory concentration (MIC) of Hg2+ in the experimental strains was determined at regular intervals by measuring the OD600 nm values of each medium for 120 h.
According to the minimum inhibitory concentration (MIC) of Hg2+ on the strains, 2% fresh bacterial solution was inoculated into the liquid LB medium with 5, 10, and 20 mg/L Hg2+; an LB medium without mercury was used as a blank control, and the optical density was measured regularly in a UV spectrophotometer to study the changes in the growth pattern of the strains under mercury stress.
2.7. Determination of Mercury Volatilization and Adsorption Capacity of the Strain
As part of the experiment, three groups were established. The positive control group contained no mercury, the negative control group was not inoculated with the strain. The experimental group was inoculated with 2% fresh bacterial broth in a liquid LB medium containing 10 mg/L Hg2+ and under optimal growth conditions, and three parallel treatments were conducted for each group. The samples were removed at regular intervals and centrifuged at 8000 rpm for 10 min. The process of digesting the supernatant and bacterium was performed as follows. A total of 1 mL supernatant from the centrifuged culture solution was mixed with 10 mL of HCl in a 50 mL Teflon crucible, the sample was heated continuously on an electric furnace, and the sample was digested until the volume was reduced to 3 mL. A total of 5 mL nitric acid was added to the solution, and the solution was heated for 1 h. The steps were repeated until the digestion was complete and the white fumes were exhausted. Then, the digestion solution was transferred to a 50 mL volumetric flask after it had cooled to room temperature and was shaken well with a fixed volume.
A cold atomic absorption spectrophotometer was used to measure the mercury concentration in the supernatant and the mercury content in the bacteria. The efficiency of total mercury removal was the initial mercury content of the culture medium minus the mercury content of the supernatant, divided by the initial mercury content of the culture medium; the efficiency of mercury adsorption by the bacteria was determined by the mercury content of the bacteria divided by the initial mercury content of the culture medium.
The mercury concentration of the supernatant and the mercury content of the bacteria were measured using a cold atomic absorption spectrophotometer. The efficiency of total mercury removal is the initial mercury content of the culture medium minus the mercury content of the supernatant, divided by the initial mercury content of the culture medium, and the efficiency of mercury adsorption by the bacteria is the mercury content of the bacteria divided by the initial mercury content of the culture medium. The total removal rate of mercury minus the adsorption efficiency of the bacteria on mercury is the volatilization efficiency of mercury [
13]. The heavy metal concentrations in the digestion solution and the blank control were measured three times. The calculation was performed as follows:
where a
1 is the initial mercury content of the culture solution (mg/L); b
1 is the mercury content of the supernatant (mg/L); c
1 is the mercury content of the bacteria (mg/L); q
1 is the total removal rate (%); q
2 is the adsorption efficiency (%); and q
3 is the volatilization efficiency (%).
2.8. Transformation Infrared Spectroscopy
To investigate the response mechanism of the LBA119 strain to mercury stress, Fourier transform infrared (FTIR) spectroscopy [
11] was used to analyze the changes in the adsorption functional groups of bacteria before and after treatment with the heavy metal Hg
2+. The bacteria were incubated in an LB medium with mercury (10 mg/L) and without mercury for 36 h. The cultures were centrifuged at 8000 rpm for 10 min, the supernatant was discarded, and the organisms were washed three times with phosphate buffer solution. The strains were freeze-dried under a vacuum, and the dried samples were thoroughly mixed with potassium bromide (KBr) powder in an agate mortar (bacterial mass: KBr mass = 1:100), pressed into thin slices using a solid press, and measured using a Fourier infrared spectrometer to record the infrared spectra in the region of 4000–400 cm
−1. The obtained spectral data were processed using Omnic 9.0 software (OMNIC 8.2.0.387).
2.9. Testing the Strains for Resistance to Different Types of Heavy Metal Ions
LB liquid medium containing single heavy metal ions (Mn
2+, Zn
2+, Pb
2+, Cd
2+, and Cr
6+) was prepared and inoculated with 2% fresh bacterial broth and incubated on a shaker at 30 °C for approximately 36 h. An LB medium with different concentration gradients (1, 100, 500, 1000, 1500, 2000, 3000, 4000, and 5000 mg/L) of heavy metals was first set to determine the crude tolerance of the strain to heavy metal concentrations [
14]. The concentration interval was then narrowed to determine the maximum concentration of heavy metals that could be tolerated. Bacterial concentrations were determined using the 600 nm optical density method (OD
600nm) to determine the minimum inhibitory concentration (MIC). Five heavy metal resistance assays were performed independently [
15], with three parallel groups for each group.
2.10. Study on the Bioremediation Effect of Strains of Bacteria on Mercury-Contaminated Soil
In this experiment, mercury-contaminated soil was collected in Luanchuan County, Henan Province, from farmland in the molybdenum mining area. The soil was air-dried and sieved, and then a specific quantity of HgCl2 solution was added to it. The soil samples were prepared with mercury concentrations of 5, 50, and 100 mg/L, stirred well, and left to stand for several days before being stirred again; then, the mercury soil was aged for 60 days.
2.11. Strain for Remediation of Mercury-Contaminated Soil
A logarithmic growth stage was achieved by incubating the bacteria. An adjustment of pH 4.6 was made to the seed solution used for inoculating LBA119 bacteria. A blank LB medium was added to the control group at the same concentration. After inoculation, the soil and inoculum were gently mixed in a shaker at 30 rpm for 20 min to conduct an exploratory experiment on the biological removal of mercury from the soil [
16].
The study involved several experimental groups, which were prepared as follows. (1) Restoration group: 100 g of mercury-aged soil samples, 95 mL of distilled water, and 5 mL of bacterial seed solution were added and shaken well to ensure that all the soil was suspended in the liquid. Sterile distilled water was replenished quantitatively every day to maintain the water content of the soil, and the new bacterial solution was added every 7 days, with an additional amount of 5 mL each time. (2) Control group: 100 g of aged soil samples containing mercury, 95 mL of distilled water, and 5 mL of the LB medium were added and shaken well, and 5 mL of fresh medium was added every 7 days.
In each experimental group, three replicates were conducted. Incubation for 7, 15, and 30 days was conducted in an incubator (temperature 30 °C, 8 h of light time and 16 h of darkness (simulated mine light time)). Each group was provided 0.5 g of air-dried soil that was ground and digested. Using cold atomic fluorescence spectrometry [
17], the mercury content was determined, and the mercury removal efficiency was calculated.
Mercury remediation efficiency was calculated by subtracting the initial mercury content of the soil from the mercury content at the time of sampling. The results are the mean ± standard deviation of three independent replicates. The efficiency of the strains for remediating mercury in the soil was calculated as follows:
where q is the removal efficiency (%) of mercury in soil by the strain; a is the initial mercury content of soil; and b is the mercury content of soil at different sampling times.
4. Discussion
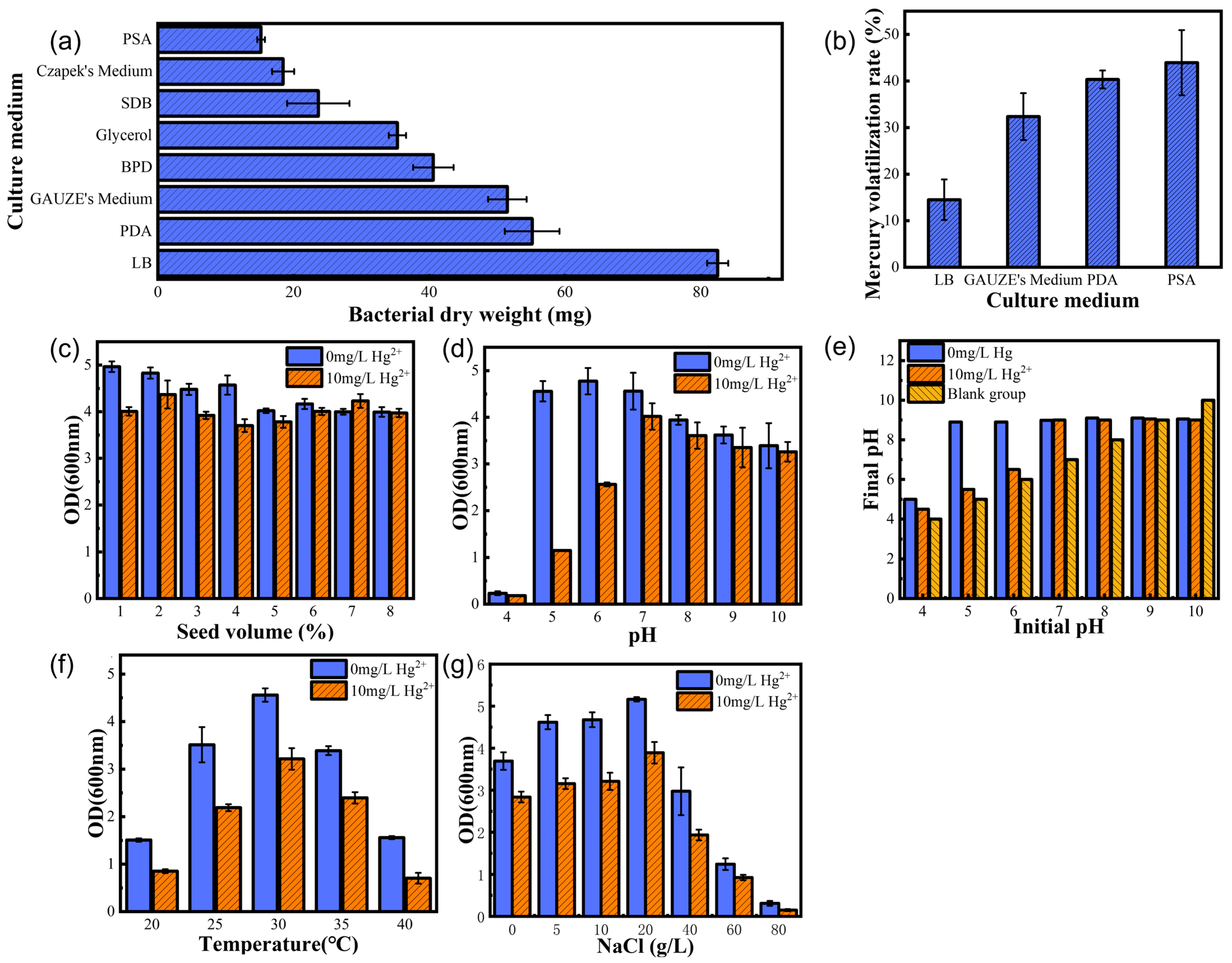
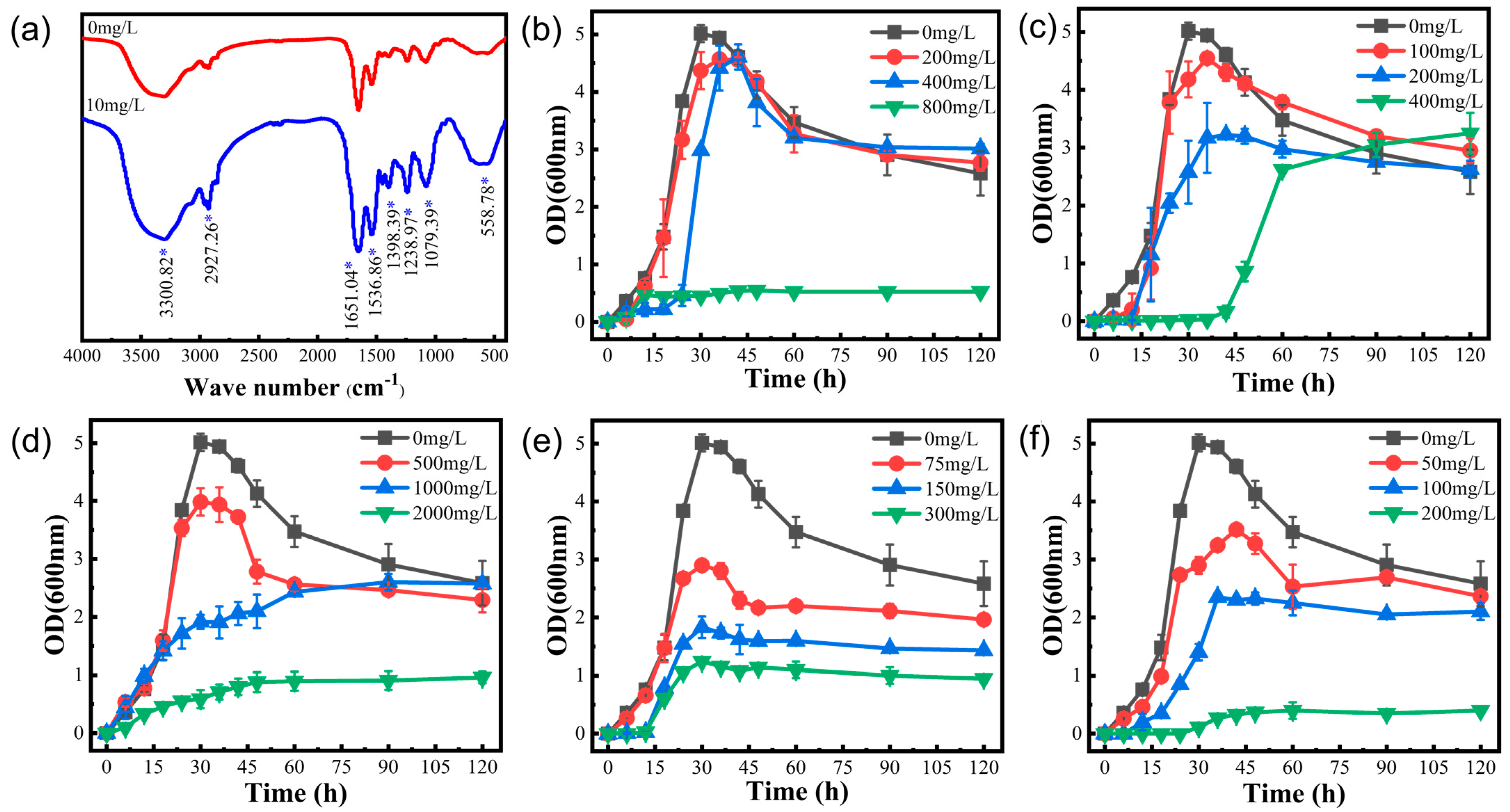
Four factors affect the growth and reproduction of the strain: optimum medium, optimum temperature, optimum pH, and optimum inoculum. Firstly, the mercury experiments in different media showed (
Figure 2b) that the volatility of mercury was higher in PSA, PDA, and GAUZE’s medium, probably because the media contained reducing sugars that could reduce Hg
2+ and volatilize after reducing Hg
2+ to Hg
0. Therefore, these media are not suitable for studies related to the study of strain tolerance to Hg. Taner Sar et al. [
23] studied the effect of inoculum level on ethanol production. We found that a low inoculum level resulted in a prolonged delay period. In contrast, a high inoculum level resulted in violent competition for survival among microorganisms and the death of a significant number of microorganisms. As shown in
Figure 2d, too high or too low pH environments resulted in significant differences in strain growth, which is because extreme pH not only affects the activity and stability of enzymes in bacteria but also reduces microbial resistance to high temperatures and nutrient uptake, and also affects the ability of bacteria to produce fatty acids, which is consistent with the extremophile green alga Coccomyxa melkonianii at different pH values, which is consistent with the lipid production and morphological behavior of the extremophile green alga [
24]. Since microorganisms do not possess thermoregulatory mechanisms, external temperature directly affects the activity of microbial intracellular enzymes, which are reduced at low temperatures. The metabolic rate and the ability of bacteria to synthesize substances decrease at low temperatures, leading to a decrease in reproductive capacity (
Figure 2f), in agreement with the view of Rosa Margesin et al. [
25], who studied the effect of different temperatures on a series of growth parameters of three bacteria. As shown in
Figure 2c, in the mercury-containing environment, the strains showed easy protein secretion and expression under optimal growth conditions compared to the control, which would increase tolerance to inorganic mercury. As a result, we determined that the optimal growth conditions for this strain in a mercury-containing environment were an LB medium with 2% inoculum, 30 °C, and an initial pH of 7.0.
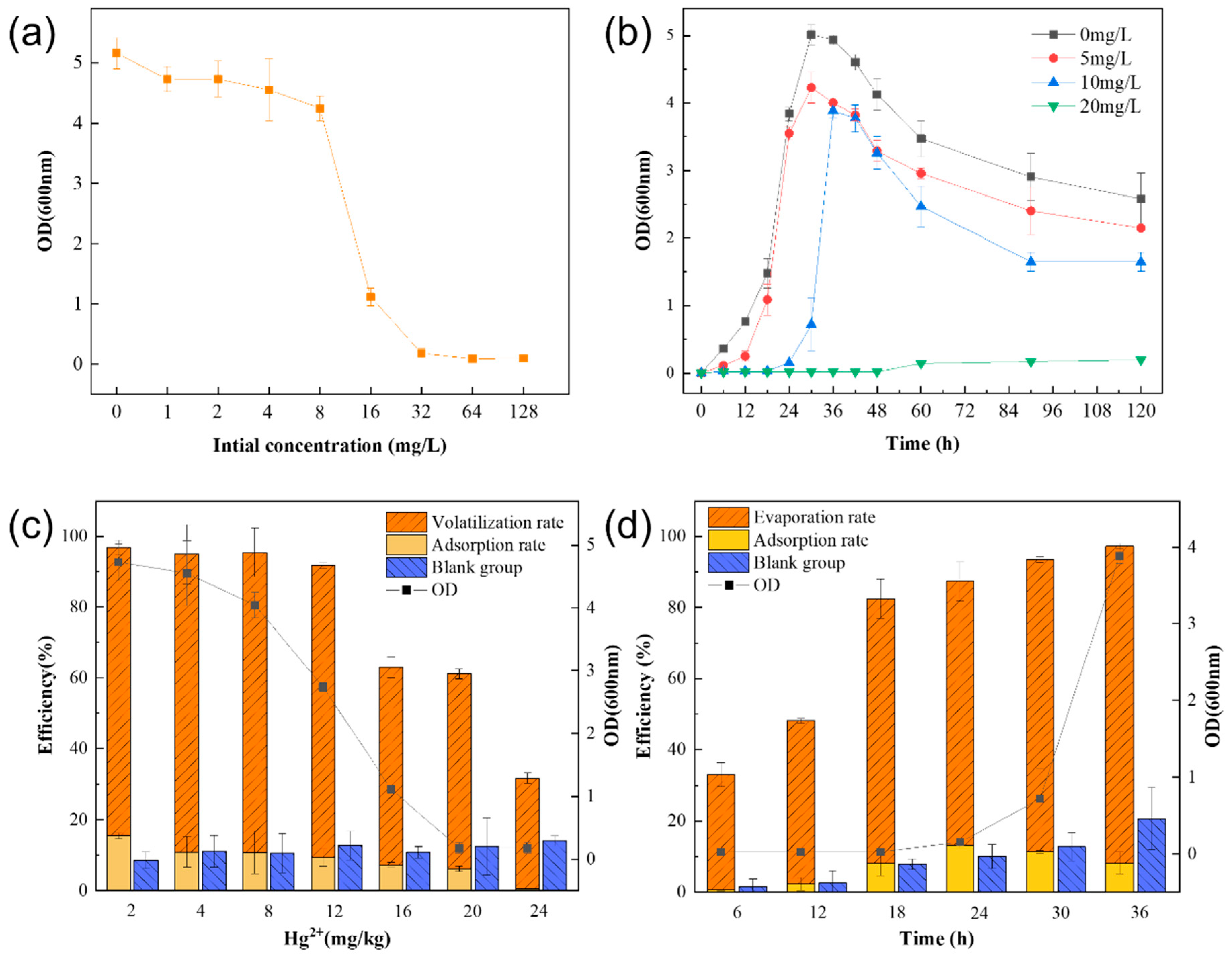
The logarithmic growth period of strain LBA119 was continuously delayed with increasing mercury concentration (
Figure 3b). The growth and removal of mercury by this strain decreased with increasing mercury concentration (
Figure 3a,c). However, mercury removal continuously increased with time (
Figure 3d). Mercuric chloride is an inorganic mercury agent; the higher the mercury concentration, the more toxic it is. High mercury concentrations inhibit bacterial strains’ growth and their bacterial sulfhydrylase activity, causing impairment of bacterial metabolism. At mercury concentrations up to 32 mg/L, the LBA119 strain stopped growing because mercury ion reductase was inhibited at high mercury concentrations, the same as the mercury tolerance of Halomonas zincidurans [
26].
As shown in
Figure 4a, by comparing the IR spectra of strain LBA119 in the LB medium containing mercury (10 mg/L) and that without mercury, it was found that OH, -CH3, C=O in the amide I band, NH in the amide II band, P-O, C-S C-O, C-OH, -COOH, C-O-C, C-O-P, M-O, and O-M-O groups were enhanced. Amide I and amide II are characteristic absorption peaks of proteins. From these peaks, it can be assumed that proteins are involved in the adsorption process to the heavy metal Hg
2+. The positively charged mercury ions combined with protein molecules to form protein salts, and irreversible precipitation of proteins (heavy metal ions precipitated proteins) occurred. The relationship between protein content and heavy metals indicated that specific proteins were critical for binding heavy metals [
27]. The presence of excessive mercury concentrations may inhibit strain growth or even result in strain death.
The main mechanisms of bacterial resistance to heavy metals are biosorption, extracellular precipitation, biotransformation, bioaccumulation, and efflux.
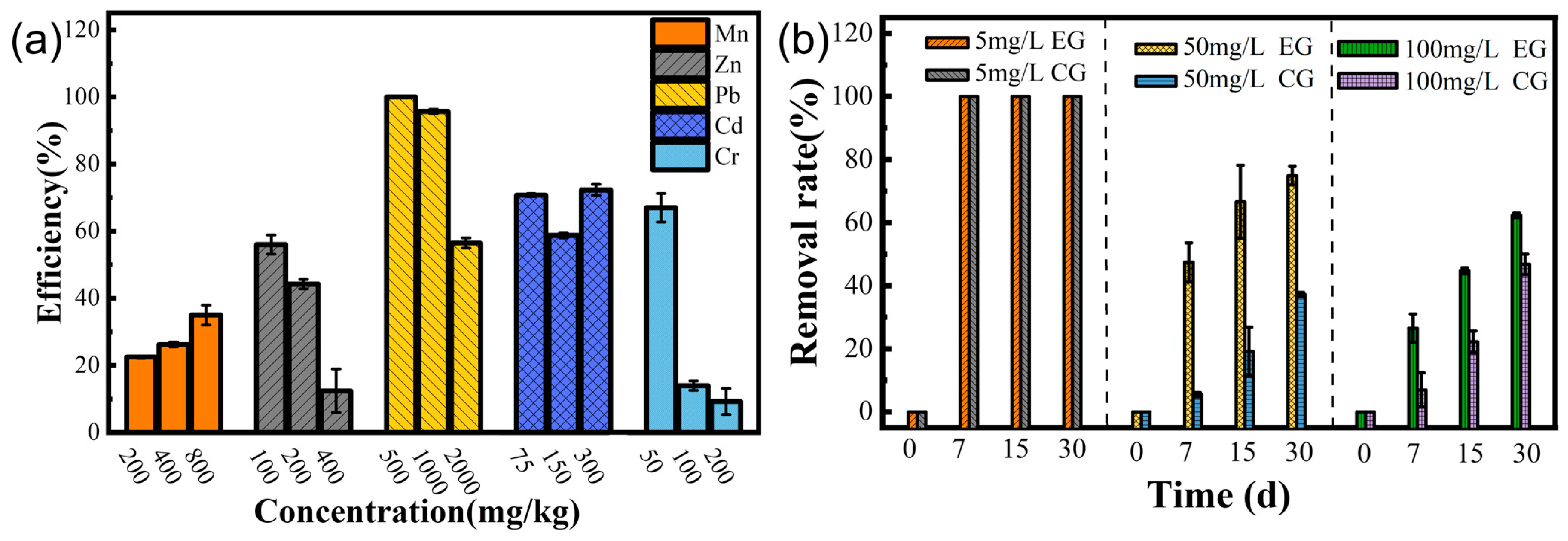
Table 2 and
Figure 4b–f show the extensive resistance of strain LBA119 to other heavy metals, with severe inhibition of growth at the maximum tolerated concentration of heavy metal ions, in agreement with the previously discussed results.
Biosorption depends on many factors, such as surface adsorption, ion exchange, ligand, chelation, and micro-precipitation, as well as the type of functional groups of the bacterial biomass. As shown in
Figure 5a, the strain showed some removal of different heavy metals, with the most effective removal of Pb. According to Hoyle-Gardner, J. et al. [
28], at neutral pH, the electrostatic attraction between Pb ions and receptors on the bacterial cell wall might facilitate the adsorption of Pb by the strain, and Pb precipitates at weak acidic pH 2 and alkaline pH above 7, in the absence of bacteria, which is consistent with our results. The LBA119 strain adjusted the environmental pH (
Figure 1e), and the mechanism of lead removal mainly relied on adsorption and the adjustment of environmental pH to precipitate lead ions.
Microbial soil remediation technology mainly relies on adsorption and transformation to remediate the soil and also has the characteristics of low investment, low pollution, and environmental friendliness. In the Orbetello Lagoon, Italy, Milva Pepi et al. [
11] have isolated mercury-tolerant bacteria that can volatilize both organic and inorganic mercury into elemental mercury and contribute to mercury removal from sediment leachate as in the removal of mercury by the strain LBA119 we studied. Thus, LBA119 is resistant to mercury and can volatilize the ionic form of mercury metal (Hg
2+) into its elemental form (Hg
0). As shown in
Figure 5b, inoculation of the LBA119 strain in high-concentration mercury-contaminated soil could improve the mercury removal rate compared with the control group, thus reducing the heavy metal mercury content in the soil. In conclusion, strain LBA119 has potential applications in the bioremediation of mercury-contaminated soils.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}