

Spleen Toxicity of Organophosphorus Flame Retardant TDCPP in Mice and the Related Mechanisms

Abstract
:
1. Introduction
2. Materials and Methods
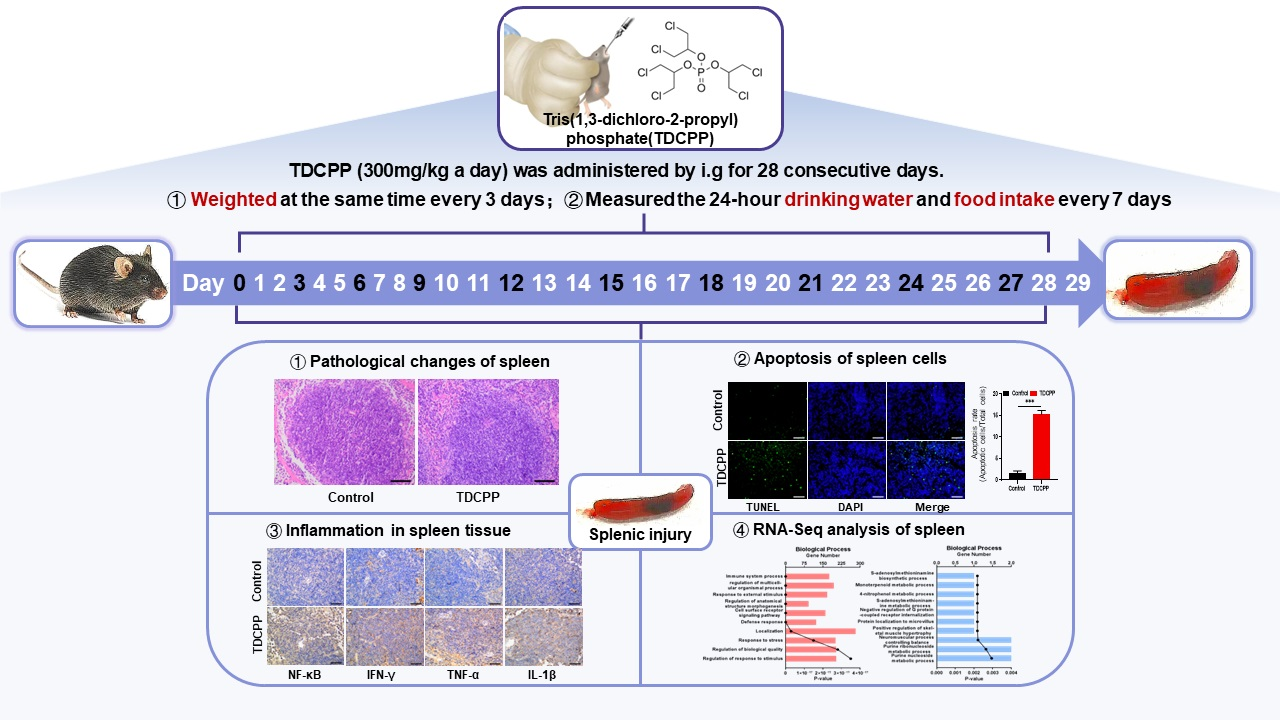
2.1. Animals and Exposure
2.2. Tissue Collection
2.3. Hematoxylin-Eosin (H&E) Staining
2.4. Immunohistochemistry (IHC) Staining
2.5. Terminal dUTP Nick-End Labeling (TUNEL) Assay
2.6. Western Blot (WB) Analysis
2.7. RNA Extraction and cDNA Library Generation
2.8. RNA-Seq Data Analysis
2.8.1. Quality Control
2.8.2. Mapping
2.8.3. Quantification of Gene Expression Level
2.8.4. Differential Expression Analysis
2.8.5. Enrichment Analysis
2.9. Quantitative Reverse-Transcription PCR(RT-qPCR) Analysis
2.10. Statistical Analysis
3. Results
3.1. Oral Exposure to TDCPP Caused Systematic Response and Induced Splenic Damage
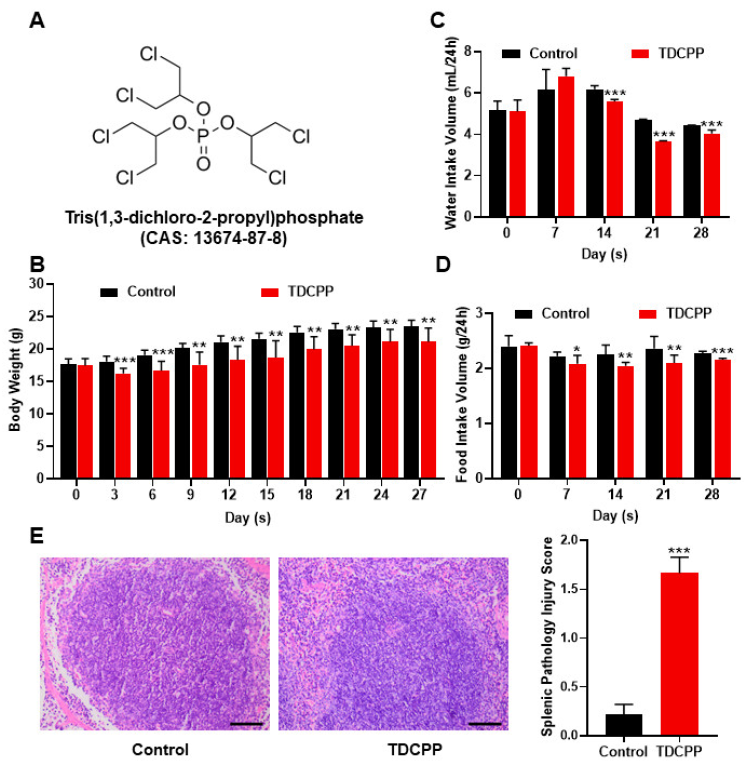
3.2. Exposure to TDCPP Activated the NF-kB Pathway and Induced Inflammation in the Spleen
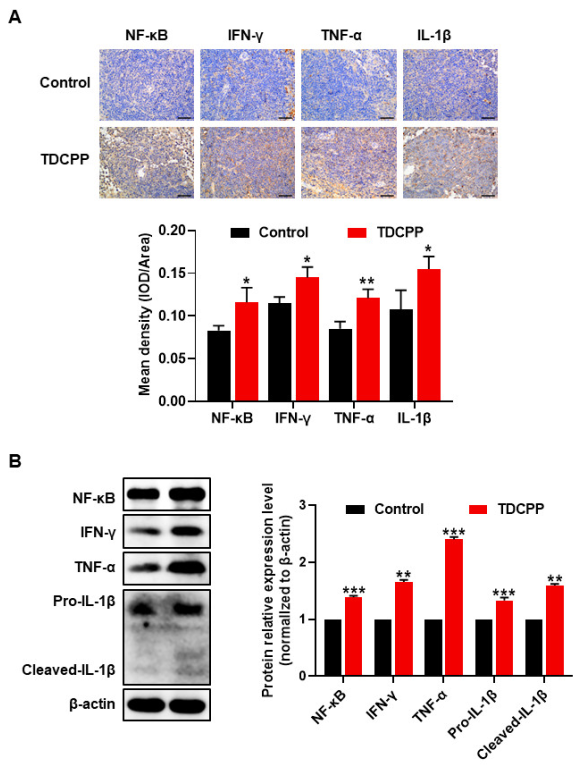
3.3. TDCPP-Induced Apoptotic Cell Death in the Spleen
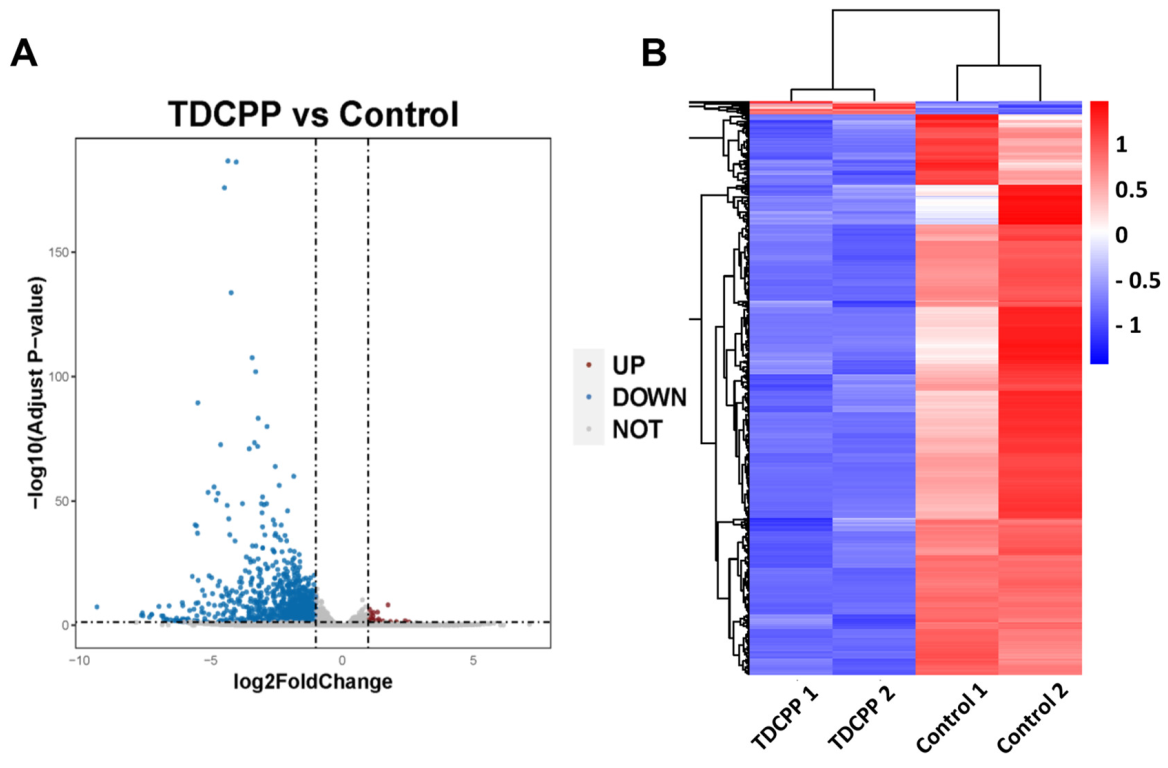
3.4. RNA-Seq Analysis of Mice Spleen Tissue
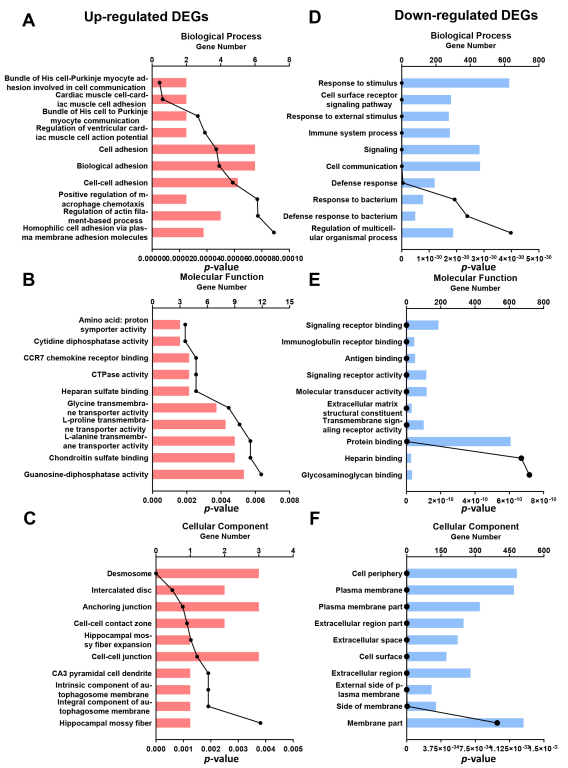
3.5. GO Analysis
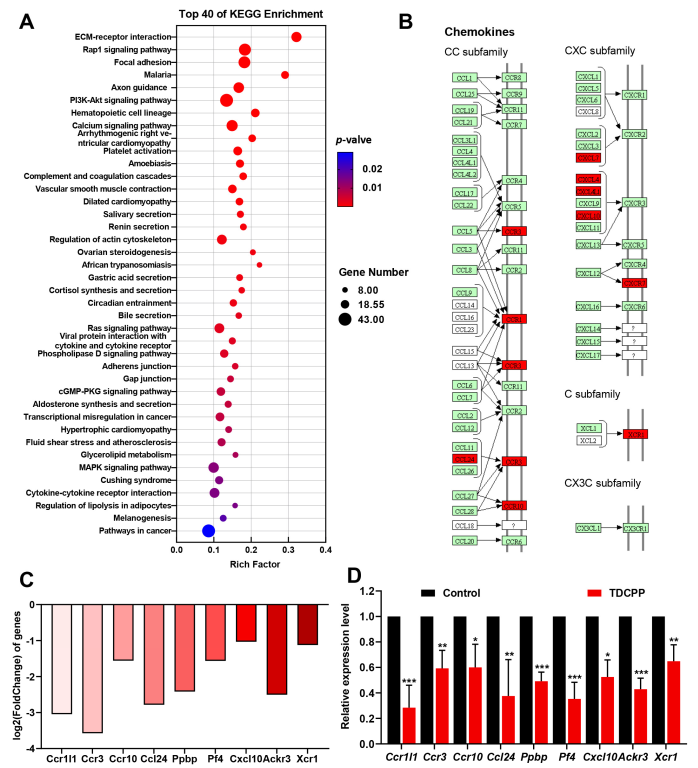
3.6. KEGG Enrichment Analysis
3.7. Validation of Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Chen, H.; Li, H.; Yu, J.; Wang, X.; Liu, Y. Review of emerging contaminant tris(1,3-dichloro-2-propyl)phosphate: Environmental occurrence, exposure, and risks to organisms and human health. Environ. Int. 2020, 143, 105946. [Google Scholar] [CrossRef]
- Sharkey, M.; Harrad, S.; Abdallah, M.A.-E.; Drage, D.S.; Berresheim, H. Phasing-out of legacy brominated flame retardants: The UNEP Stockholm Convention and other legislative action worldwide. Environ. Int. 2020, 144, 106041. [Google Scholar] [CrossRef]
- Akortia, E.; Okonkwo, J.O.; Lupankwa, M.; Osae, S.D.; Daso, A.P.; Olukunle, O.I.; Chaudhary, A. A review of sources, levels, and toxicity of polybrominated diphenyl ethers (PBDEs) and their transformation and transport in various environmental compartments. Environ. Rev. 2016, 24, 253–273. [Google Scholar] [CrossRef]
- Alcock, R.E.; Sweetman, A.J.; Prevedouros, K.; Jones, K.C. Understanding levels and trends of BDE-47 in the UK and North America: An assessment of principal reservoirs and source inputs. Environ. Int. 2003, 29, 691–698. [Google Scholar] [CrossRef]
- Rodgers, T.F.M.; Truong, J.; Jantunen, L.M.; Helm, P.A.; Diamond, M.L. Organophosphate Ester Transport, Fate, and Emissions in Toronto, Canada, Estimated Using an Updated Multimedia Urban Model. Environ. Sci. Technol. 2018, 52, 12465–12474. [Google Scholar] [CrossRef]
- Ding, J.; Xu, Z.; Huang, W.; Feng, L.; Yang, F. Organophosphate ester flame retardants and plasticizers in human placenta in Eastern China. Sci. Total. Environ. 2016, 554–555, 211–217. [Google Scholar] [CrossRef]
- Wang, X.L.; Liu, Q.; Zhong, W.J.; Yang, L.P.; Yang, J.; Covaci, A.; Zhu, L. Estimating renal and hepatic clearance rates of organophosphate esters in humans: Impacts of intrinsic metabolism and binding affinity with plasma proteins. Environ. Int. 2020, 134, 105321. [Google Scholar] [CrossRef]
- Carignan, C.C.; Mínguez-Alarcón, L.; Butt, C.M.; Williams, P.L.; Meeker, J.D.; Stapleton, H.M.; Toth, T.L.; Ford, J.B.; Hauser, R. Erratum: “Urinary Concentrations of Organophosphate Flame Retardant Metabolites and Pregnancy Outcomes among Women Undergoing in Vitro Fertilization”. Environ. Health Perspect. 2017, 125, 119001. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Han, J.; Zhou, B.; Gong, Z.; Santos, E.M.; Huo, X.; Zheng, W.; Liu, H.; Yu, H.; Liu, C. Toxicogenomic Responses of Zebrafish Embryos/Larvae to Tris(1,3-dichloro-2-propyl) Phosphate (TDCPP) Reveal Possible Molecular Mechanisms of Developmental Toxicity. Environ. Sci. Technol. 2013, 47, 10574–10582. [Google Scholar] [CrossRef]
- Carignan, C.C.; McClean, M.D.; Cooper, E.M.; Watkins, D.J.; Fraser, A.J.; Heiger-Bernays, W.; Stapleton, H.M.; Webster, T.F. Predictors of tris(1,3-dichloro-2-propyl) phosphate metabolite in the urine of office workers. Environ. Int. 2013, 55, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Saquib, Q.; Al-Salem, A.M.; Siddiqui, M.A.; Ansari, S.M.; Zhang, X.; Al-Khedhairy, A.A. Organophosphorus Flame Retardant TDCPP Displays Genotoxic and Carcinogenic Risks in Human Liver Cells. Cells 2022, 11, 195. [Google Scholar] [CrossRef]
- Li, R.; Zhou, P.; Guo, Y.; Lee, J.-S.; Zhou, B. Tris (1,3-dichloro-2-propyl) phosphate-induced apoptotic signaling pathways in SH-SY5Y neuroblastoma cells. Neurotoxicology 2017, 58, 1–10. [Google Scholar] [CrossRef]
- Zhong, X.; Wu, J.; Ke, W.; Yu, Y.; Ji, D.; Kang, J.; Qiu, J.; Wang, C.; Yu, P.; Wei, Y. Neonatal exposure to organophosphorus flame retardant TDCPP elicits neurotoxicity in mouse hippocampus via microglia-mediated inflammation in vivo and in vitro. Arch. Toxicol. 2020, 94, 541–552. [Google Scholar] [CrossRef]
- Liu, Y.; Le, Y.; Xu, M.; Wang, W.; Chen, H.; Zhang, Q.; Wang, C. Remodeling on adipocytic physiology of organophosphorus esters in mature adipocytes. Environ. Pollut. 2022, 305, 119287. [Google Scholar] [CrossRef]
- Li, X.; Li, N.; Rao, K.; Huang, Q.; Ma, M. In Vitro Immunotoxicity of Organophosphate Flame Retardants in Human THP-1-Derived Macrophages. Environ. Sci. Technol. 2020, 54, 8900–8908. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, eaau6085. [Google Scholar] [CrossRef]
- Bronte, V.; Pittet, M.J. The Spleen in Local and Systemic Regulation of Immunity. Immunity 2013, 39, 806–818. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, Y.; Han, Q.; Yao, Y.; Xing, H.; Teng, X. Immunosuppression, oxidative stress, and glycometabolism disorder caused by cadmium in common carp (Cyprinus carpio L.): Application of transcriptome analysis in risk assessment of environmental contaminant cadmium. J. Hazard. Mater. 2019, 366, 386–394. [Google Scholar] [CrossRef]
- Kamata, E.; Naito, K.; Nakaji, Y.; Ogawa, Y.; Suzuki, S.; Kaneko, T.; Takada, K.; Kurokawa, Y.; Tobe, M. Acute and subacute toxicity studies of tris (1,3-dichloro-2-propyl) phosphate on mice. Eisei Shikenjo Hokoku. Bull. Natl. Inst. Hyg. Sci. 1989, 107, 36–43. [Google Scholar]
- Momtazi-Borojeni, A.A.; Banach, M.; Tabatabaei, S.A.; Sahebkar, A. Preclinical toxicity assessment of a peptide-based antiPCSK9 vaccine in healthy mice. Biomed. Pharmacother. 2023, 158, 114170. [Google Scholar] [CrossRef]
- Fernández-Carrobles, M.M.; Bueno, G.; García-Rojo, M.; González-López, L.; López, C.; Déniz, O. Automatic quantification of IHC stain in breast TMA using colour analysis. Comput. Med. Imaging Graph. 2017, 61, 14–27. [Google Scholar] [CrossRef]
- Wang, H.; Yang, L.-L.; Ji, Y.-L.; Chen, Y.-H.; Hu, J.; Zhang, C.; Zhang, J.; Xu, D.-X. Different fixative methods influence histological morphology and TUNEL staining in mouse testes. Reprod. Toxicol. 2016, 60, 53–61. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Luo, H.; Wang, J.; Qiao, C.; Ma, N.; Liu, D.; Zhang, W. Pycnogenol attenuates atherosclerosis by regulating lipid metabolism through the TLR4–NF-κB pathway. Exp. Mol. Med. 2015, 47, e191. [Google Scholar] [CrossRef] [Green Version]
- Van Herreweghe, F.; Festjens, N.; Declercq, W.; Vandenabeele, P. Tumor necrosis factor-mediated cell death: To break or to burst, that’s the question. Cell. Mol. Life Sci. 2010, 67, 1567–1579. [Google Scholar] [CrossRef]
- Vilgelm, A.E.; Richmond, A. Chemokines Modulate Immune Surveillance in Tumorigenesis, Metastasis, and Response to Immunotherapy. Front. Immunol. 2019, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Poma, G.; Sales, C.; Bruyland, B.; Christia, C.; Goscinny, S.; Van Loco, J.; Covaci, A. Occurrence of Organophosphorus Flame Retardants and Plasticizers (PFRs) in Belgian Foodstuffs and Estimation of the Dietary Exposure of the Adult Population. Environ. Sci. Technol. 2018, 52, 2331–2338. [Google Scholar] [CrossRef]
- Liu, X.; Ji, K.; Jo, A.; Moon, H.-B.; Choi, K. Effects of TDCPP or TPP on gene transcriptions and hormones of HPG axis, and their consequences on reproduction in adult zebrafish (Danio rerio). Aquat. Toxicol. 2013, 134–135, 104–111. [Google Scholar] [CrossRef]
- McGee, S.P.; Cooper, E.M.; Stapleton, H.M.; Volz, D.C. Early Zebrafish Embryogenesis Is Susceptible to Developmental TDCPP Exposure. Environ. Health Perspect. 2012, 120, 1585–1591. [Google Scholar] [CrossRef]
- Moser, G.; McDaniel, K.; Phillips, P.; Hedge, J. Neurobehavioral and thyroid evaluations of rats developmentally exposed to tris(1,3-dichloro-2-propyl)phosphate (TDCPP). Neurotoxicol. Teratol. 2014, 43, 97. [Google Scholar] [CrossRef]
- Liu, X.; Cai, Y.; Wang, Y.; Xu, S.; Ji, K.; Choi, K. Corrigendum to “Effects of tris(1,3-dichloro-2-propyl) phosphate (TDCPP) and triphenyl phosphate (TPP) on sex dependent alterations of thyroid hormones in adult zebrafish” [Ecotoxicol. Environ. Saf. 170 (2019) 25–32]. Ecotoxicol. Environ. Saf. 2020, 201, 110867. [Google Scholar] [CrossRef]
- Cheng, L.; Rao, Q.; Zhang, Q.; Song, W.; Guan, S.; Jiang, Z.; Wu, T.; Zhao, Z.; Song, W. The immunotoxicity of decabromodiphenyl ether (BDE-209) on broiler chicks by transcriptome profiling analysis. Ecotoxicol. Environ. Saf. 2022, 232, 113284. [Google Scholar] [CrossRef]
- Hao, R.; Jiang, Y.; Li, F.; Sun-Waterhouse, D.; Li, D. MiR-182–5p/TLR4/NF-κB axis contributes to the protective effect of caffeic acid phenethyl ester against cadmium-induced spleen toxicity and associated damage in mice. Food Chem. Toxicol. 2021, 158, 112654. [Google Scholar] [CrossRef]
- Sun, H.D.; Malabunga, M.; Tonra, J.R.; DiRenzo, R.; Carrick, F.E.; Zheng, H.; Berthoud, H.-R.; McGuinness, O.P.; Shen, J.; Bohlen, P.; et al. Monoclonal antibody antagonists of hypothalamic FGFR1 cause potent but reversible hypophagia and weight loss in rodents and monkeys. Am. J. Physiol. Metab. 2007, 292, E964–E976. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Li, Y.; He, C.; Yan, F.; Li, J.-R.; Xu, H.-Z.; Zhuang, J.-F.; Zhou, H.; Peng, Y.-C.; Fu, X.-J.; et al. Selective Ferroptosis Inhibitor Liproxstatin-1 Attenuates Neurological Deficits and Neuroinflammation After Subarachnoid Hemorrhage. Neurosci. Bull. 2021, 37, 535–549. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Regulation of NF-κB by TNF family cytokines. Semin. Immunol. 2014, 26, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Napetschnig, J.; Wu, H. Molecular Basis of NF-κB Signaling. Annu. Rev. Biophys. 2013, 42, 443–468. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Frankel, T.L.; Green, M.; Zou, W. IFNγ signaling integrity in colorectal cancer immunity and immunotherapy. Cell. Mol. Immunol. 2022, 19, 23–32. [Google Scholar] [CrossRef]
- Ni, R.; Fan, L.; Zhang, L.; Song, Y.; Wang, H.; Wang, A.; Liu, B. A mouse model of irradiation and spleen-thymus lymphocyte infusion induced aplastic anemia. Hematology 2022, 27, 932–945. [Google Scholar] [CrossRef]
- Cui, D.; Bi, J.; Zhang, Z.-N.; Li, M.-Y.; Qin, Y.-S.; Xiang, P.; Ma, L.Q. Organophosphorus flame retardant TDCPP-induced cytotoxicity and associated mechanisms in normal human skin keratinocytes. Sci. Total. Environ. 2020, 726, 138526. [Google Scholar] [CrossRef]
- Wan, F.; Tang, L.; Rao, G.; Zhong, G.; Jiang, X.; Wu, S.; Huang, R.; Tang, Z.; Ruan, Z.; Chen, Z.; et al. Curcumin activates the Nrf2 Pathway to alleviate AFB1-induced immunosuppression in the spleen of ducklings. Toxicon 2022, 209, 18–27. [Google Scholar] [CrossRef]
- Yan, N.; Xu, G.; Zhang, C.; Liu, X.; Li, X.; Sun, L.; Wang, D.; Duan, X.; Li, B. Chronic arsenic exposure induces the time-dependent modulation of inflammation and immunosuppression in spleen. Cell Biosci. 2020, 10, 91. [Google Scholar] [CrossRef]
- Hughes, C.E.; Nibbs, R.J.B. A guide to chemokines and their receptors. FEBS J. 2018, 285, 2944–2971. [Google Scholar] [CrossRef]
- Korbecki, J.; Grochans, S.; Gutowska, I.; Barczak, K.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of Receptors CCR5, CCR6, CCR7, CCR8, CCR9, and CCR10 Ligands. Int. J. Mol. Sci. 2020, 21, 7619. [Google Scholar] [CrossRef]
- Zhou, S.-F.; Ma, J.; Qu, H.-T.; Liu, Z.-T.; He, W.-D.; Wang, J.-D.; Dou, A.-X.; Zhang, N.; Liu, J.-L.; Guo, C.-S.; et al. Characterization of Th1- and Th2-associated Chemokine Receptor Expression in Spleens of Patients with Immune Thrombocytopenia. J. Clin. Immunol. 2013, 33, 938–946. [Google Scholar] [CrossRef]
- Topham, P.S.; Csizmadia, V.; Soler, D.; Hines, D.; Gerard, C.J.; Salant, D.J.; Hancock, W.W. Lack of chemokine receptor CCR1 enhances Th1 responses and glomerular injury during nephrotoxic nephritis. J. Clin. Investig. 1999, 104, 1549–1557. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Kojder, K.; Simińska, D.; Bohatyrewicz, R.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of the Ligands of Receptors CCR1, CCR2, CCR3, and CCR4. Int. J. Mol. Sci. 2020, 21, 8412. [Google Scholar] [CrossRef]
- Cabrero-de las Heras, S.; Martínez-Balibrea, E. CXC family of chemokines as prognostic or predictive biomarkers and possible drug targets in colorectal cancer. World J. Gastroenterol. 2018, 24, 4738–4749. [Google Scholar] [CrossRef]
- Lei, Y.; Takahama, Y. XCL1 and XCR1 in the immune system. Microbes Infect. 2012, 14, 262–267. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Primer Sequence (5′–3′) | Tm (°C) | Length (bp) |
|---|---|---|---|---|
| Chemokine | ||||
| CC subfamily | ||||
| Ccr1l1 | 12770 | GGCATCATCAGTAGCATCA | 55 | 303 |
| GAGGAAGAAGAGAAGCGTAA | ||||
| Ccr3 | 12771 | GCTTTGAGACCACACCCTATG | 58 | 136 |
| ACCATTATGTTGCCCAGGAG | ||||
| Ccr10 | 12777 | ATGCTTCACTCGGTCTCT | 55 | 127 |
| CACTACTGGATAGCGATAGG | ||||
| Ccl24 | 56221 | GGTTCAGAGGCACATACAA | 55 | 492 |
| AGAGATGGACAGACAGACA | ||||
| CXC subfamily | ||||
| Ppbp | 57349 | TGTGCTGATGTGGAAGTGATAG | 58 | 150 |
| CTGAGCAGGAAAATGGTTTGG | ||||
| Pf4 | 56744 | GAGGTGATCAAGGCAGGAC | 58 | 141 |
| AGCTGATACCTAACTCTCCAGG | ||||
| Cxcl10 | 15945 | GATGGATGGACAGCAGAG | 54 | 417 |
| GGAAGATGGTGGTTAAGTTC | ||||
| Ackr3 | 12778 | TTCATCAACCGCAACTACA | 55 | 260 |
| TCTCCTCTTCATACCACTCA | ||||
| C subfamily | ||||
| Xcr1 | 23832 | ATCTTCTTCCTCCTCTCCAT | 55 | 473 |
| ATCCACTTCTCCTTGTCTTC | ||||
| Glyceraldehyde-3-phosphate dehydrogenase protein family | ||||
| Gapdh | 14433 | TCTCCTGCGACTTCAACA | 56 | 117 |
| TGTAGCCGTATTCATTGTCA | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, L.; Wei, L.; Du, Q.; Su, Y.; Ye, S.; Liu, K. Spleen Toxicity of Organophosphorus Flame Retardant TDCPP in Mice and the Related Mechanisms. Toxics 2023, 11, 231. https://doi.org/10.3390/toxics11030231
Cao L, Wei L, Du Q, Su Y, Ye S, Liu K. Spleen Toxicity of Organophosphorus Flame Retardant TDCPP in Mice and the Related Mechanisms. Toxics. 2023; 11(3):231. https://doi.org/10.3390/toxics11030231
Chicago/Turabian StyleCao, Lanqin, Lai Wei, Qiaoyun Du, Ying Su, Shuzi Ye, and Kaihua Liu. 2023. "Spleen Toxicity of Organophosphorus Flame Retardant TDCPP in Mice and the Related Mechanisms" Toxics 11, no. 3: 231. https://doi.org/10.3390/toxics11030231