
K252a Prevents Microglial Activation Induced by Anoxic Stimulation of Carotid Bodies in Rats
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Substances and Compounds
2.3. Surgery
2.4. Intracerebroventricular Injection in the Left Lateral Ventricle (LV)
2.5. Microglia (Griffonia simplicifolia)
2.6. Statistical Analysis
3. Results
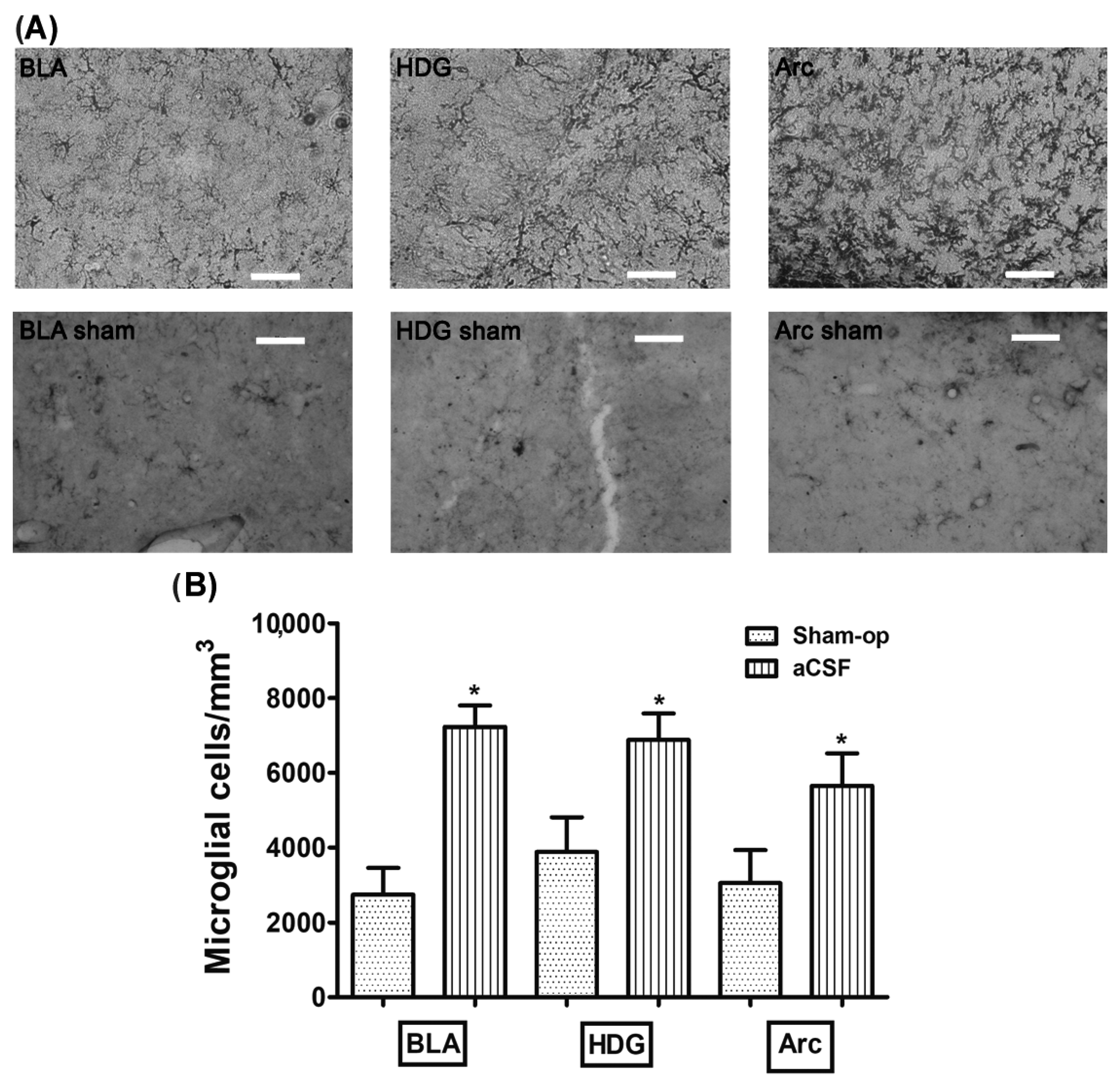
3.1. Stimulation of the CBC with NaCN Increases the Density of Microglia in the Amygdala, Dentate Gyrus Hilus, and Arcuate
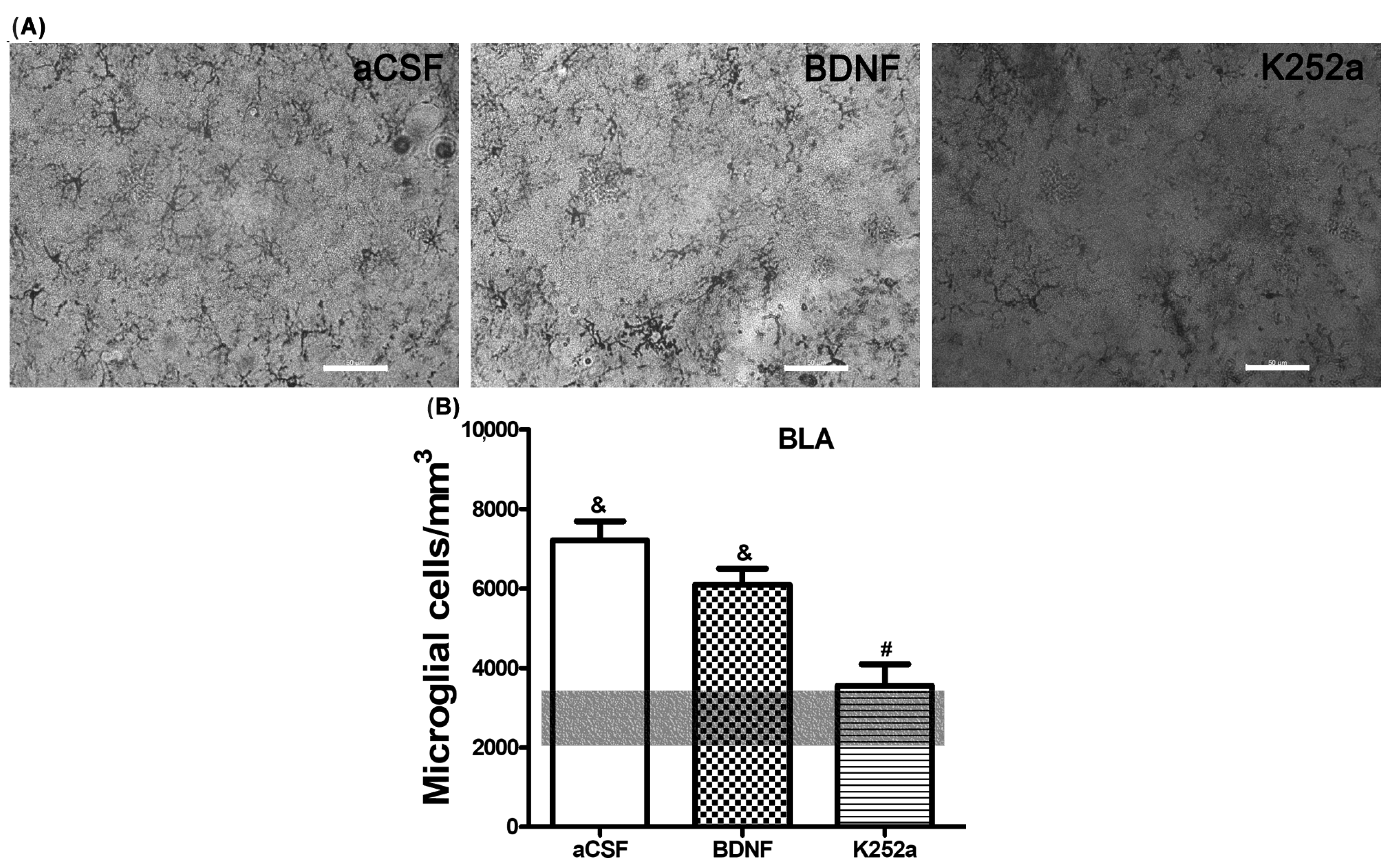
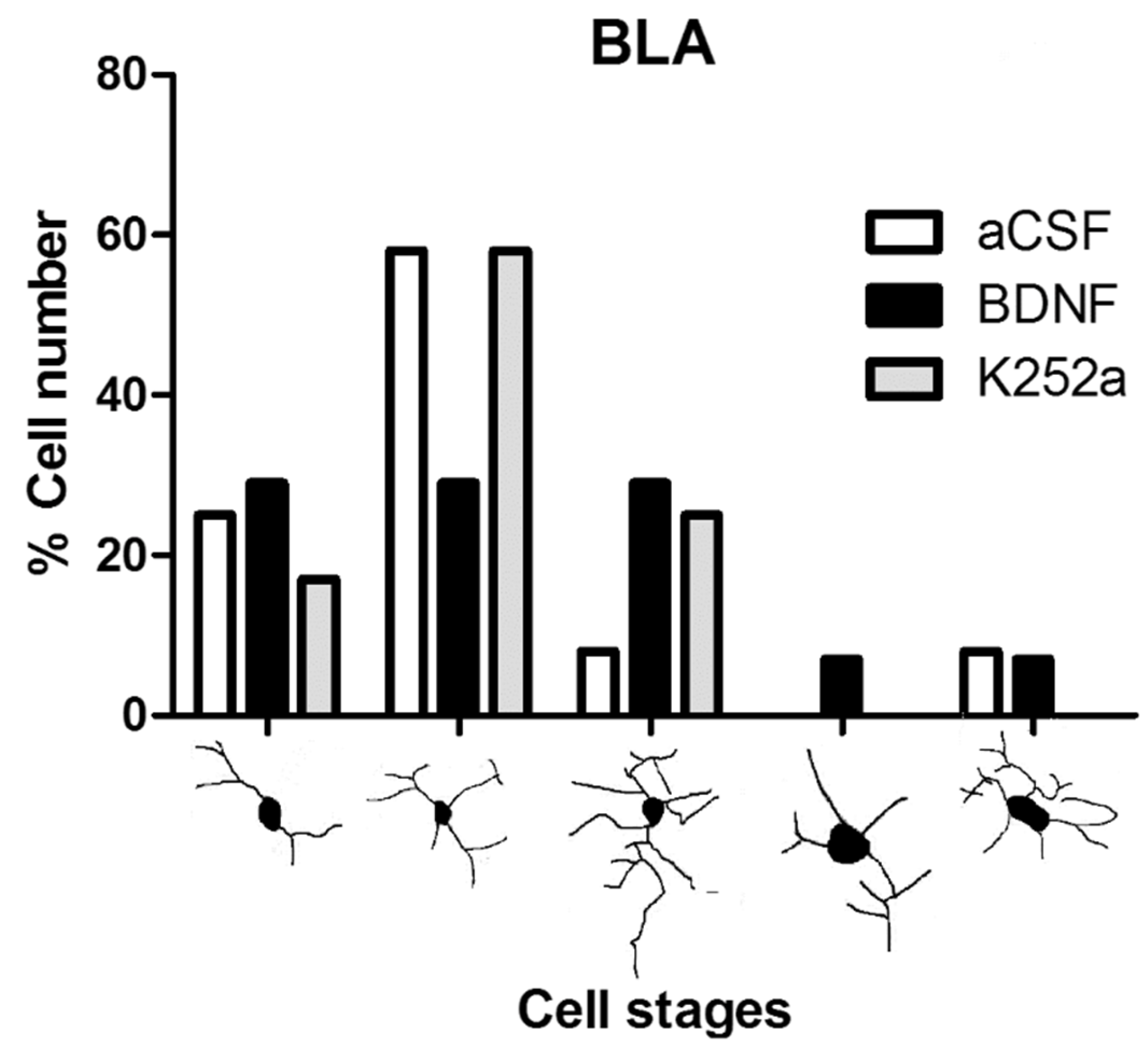
3.2. The Infusion of K252a in the Left LV after the Injection of NaCN in the CBC Prevented Microglial Reactivity in the Basolateral Amygdala
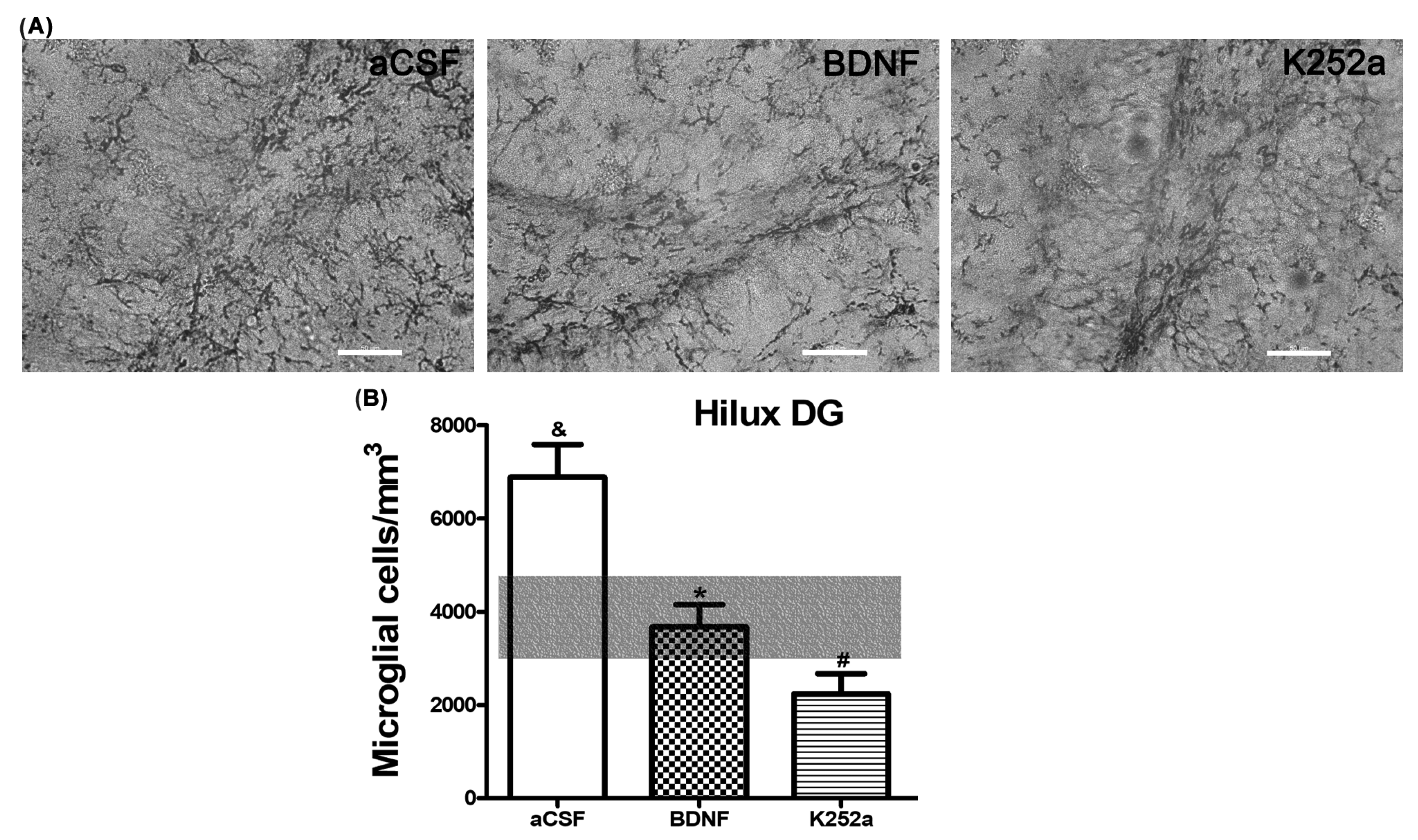
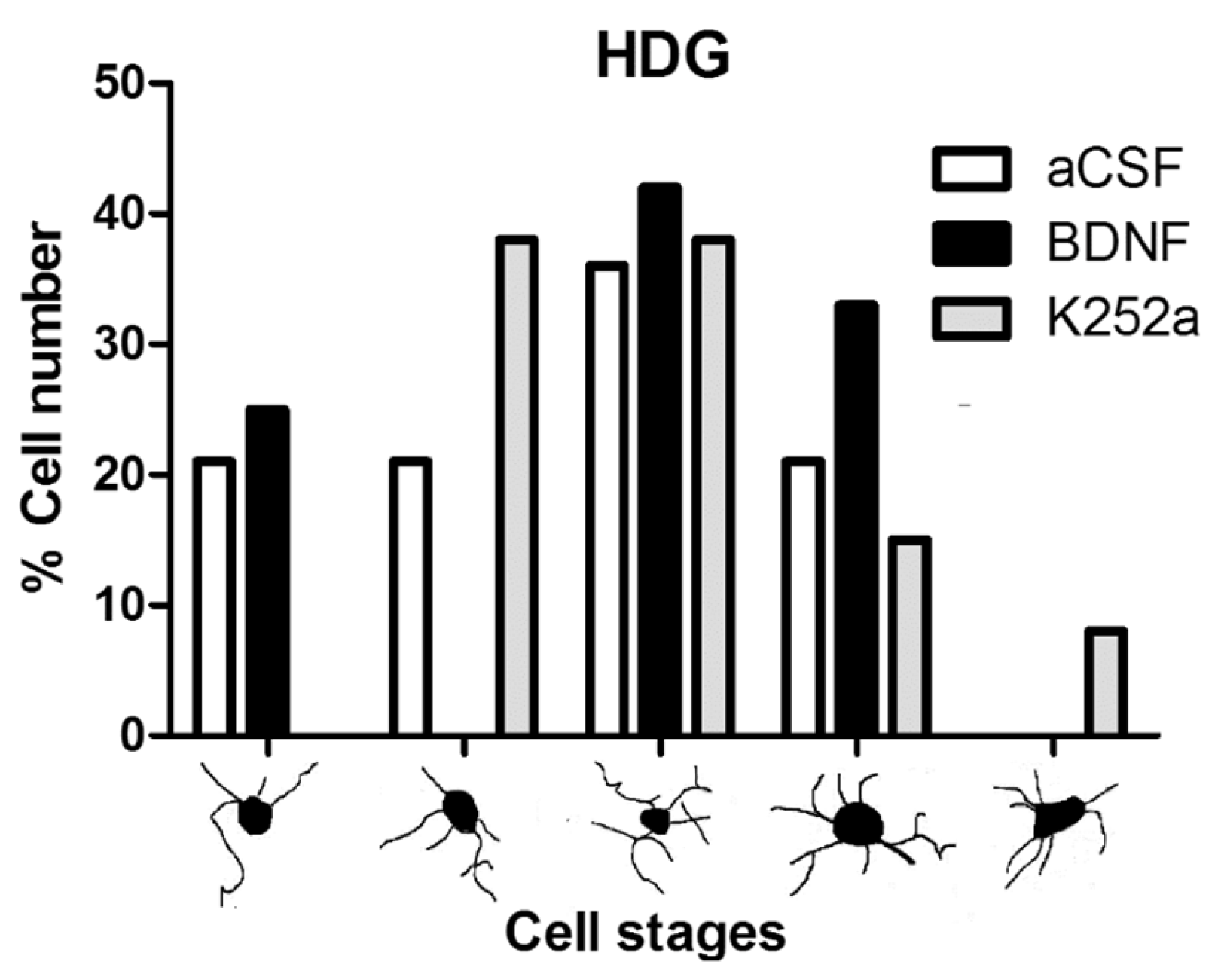
3.3. Infusion of BDNF or K252a in the Left LV Prior to the CBC Stimulation Prevented the Increase in Microglial Cell Density in the Hilus of the Dentate Gyrus
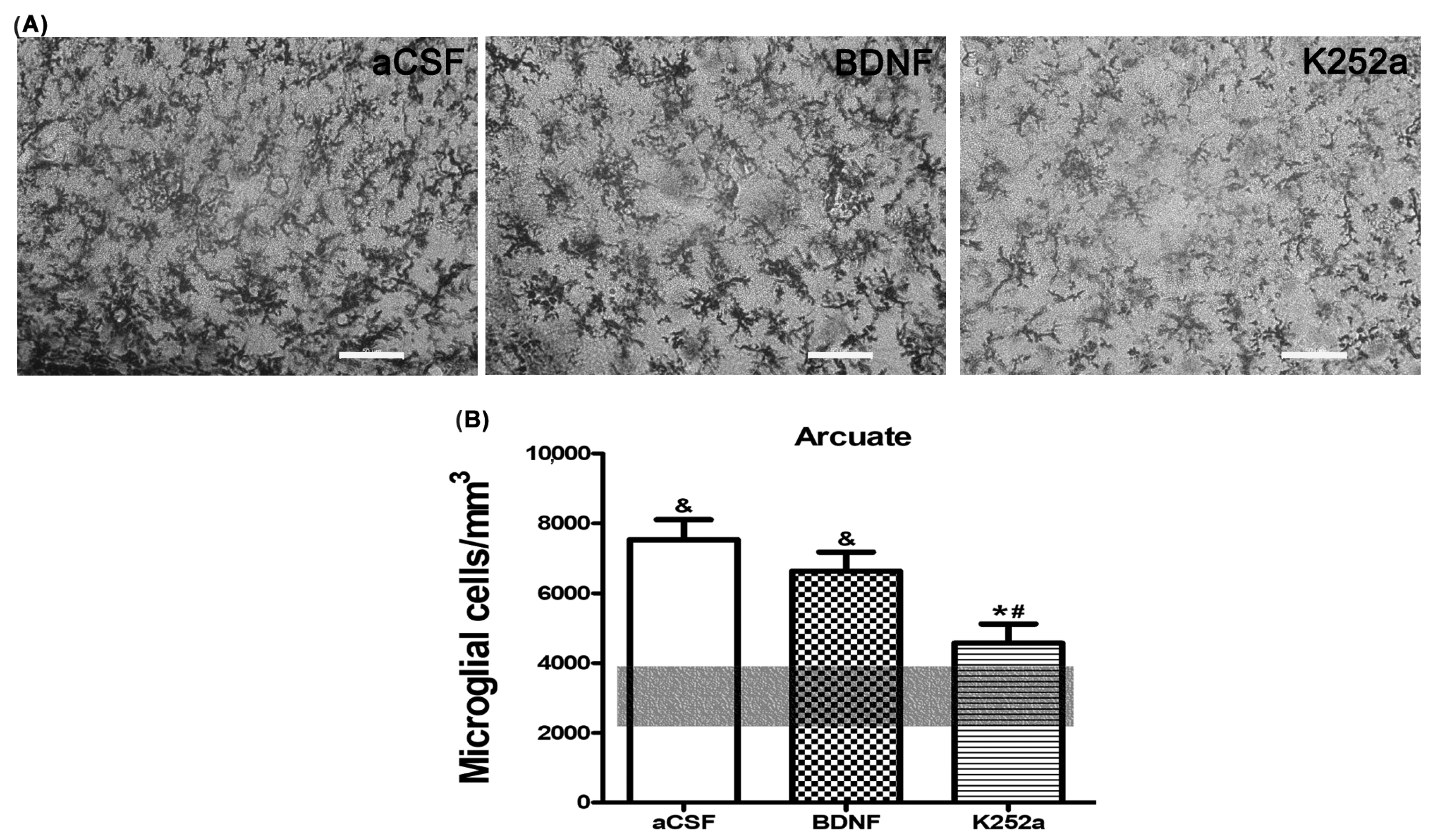
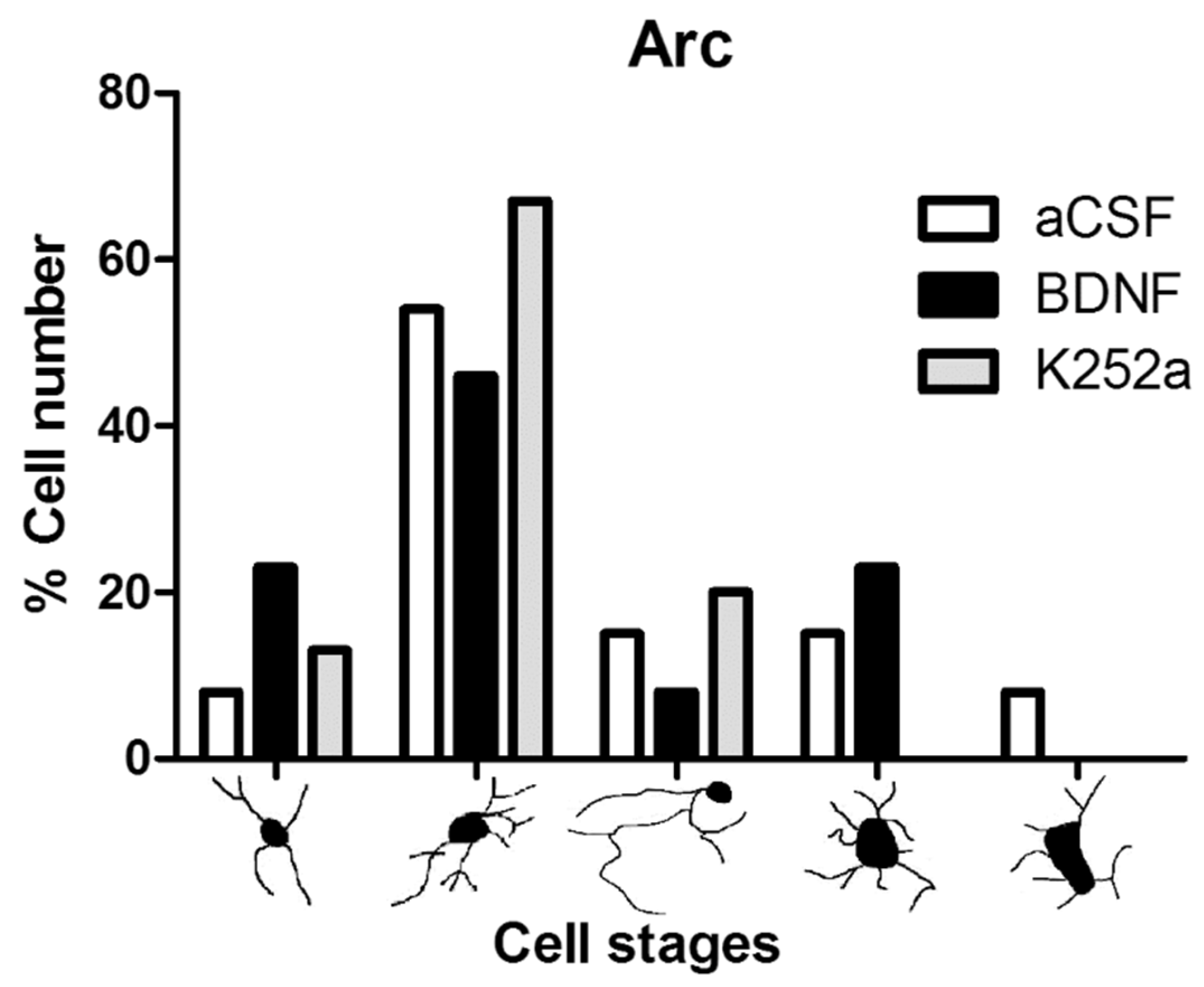
3.4. Blocking the TrkB Receptor Prior to CBC Stimulation Prevented the Increase in Microglia Density in the Arcuate Nucleus of the Hypothalamus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Joyner, M.J.; Limberg, J.K.; Wehrwein, E.A.; Johnson, B.D. Role of the carotid body chemoreceptors in glucose homeostasis and thermoregulation in humans. J. Physiol. 2018, 596, 3079–3085. [Google Scholar] [CrossRef]
- Limberg, J.K.; Taylor, J.L.; Mozer, M.T.; Dube, S.; Basu, A.; Basu, R.; Rizza, R.A.; Curry, T.B.; Joyner, M.J.; Wehrwein, E.A. Effect of bilateral carotid body resection on cardiac baroreflex control of blood pressure during hypoglycemia. Hypertension 2015, 65, 1365–1371. [Google Scholar] [CrossRef] [PubMed]
- Iturriaga, R.; Alcayaga, J.; Chapleau, M.W.; Somers, V.K. Carotid body chemoreceptors: Physiology, pathology, and implications for health and disease. Physiol. Rev. 2021, 101, 1177–1235. [Google Scholar] [CrossRef]
- Prabhakar, N.R.; Peng, Y.J.; Kumar, G.K.; Nanduri, J. Peripheral chemoreception and arterial pressure responses to intermittent hypoxia. Comprehe. Physiol. 2015, 5, 561–577. [Google Scholar]
- Alvarez-Buylla, R.; de Alvarez-Buylla, E.R. Carotid sinus receptors participate in glucose homeostasis. Respir. Physiol. 1988, 72, 347–359. [Google Scholar] [CrossRef]
- Pardal, R.; López-Barneo, J. Low glucose-sensing cells in the carotid body. Nat. Neurosci. 2002, 5, 197–198. [Google Scholar] [CrossRef] [PubMed]
- Isom, G.E.; Way, J.L. Effects of oxygen on the antagonism of cyanide intoxication: Cytochrome oxidase, in vitro. Toxicol. Appl. Pharmacol. 1984, 74, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Housley, G.D.; Sinclair, J.D. Localization by kainic acid lesions of neurones transmitting the carotid chemoreceptor stimulus for respiration in rat. J. Physiol. 1988, 406, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Ruyle, B.C.; Klutho, P.J.; Baines, C.P.; Heesch, C.M.; Hasser, E.M. Hypoxia activates a neuropeptidergic pathway from the paraventricular nucleus of the hypothalamus to the nucleus tractus solitarii. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R1167–R1182. [Google Scholar] [CrossRef]
- Luo, Y.; Kaur, C.; Ling, E.A. Hypobaric hypoxia induces fos and neuronal nitric oxide synthase expression in the paraventricular and supraoptic nucleus in rats. Neurosci. Lett. 2000, 296, 145–148. [Google Scholar] [CrossRef]
- Babović, S.S.; Žigić, S.; Šakić, B. c-Fos protein expression in the anterior amygdaloid area and nc. accumbens in the hypoxic rat brain. Med. Pregl. 2014, 67, 379–384. [Google Scholar] [CrossRef]
- Tadmouri, A.; Champagnat, J.; Morin-Surun, M.P. Activation of microglia and astrocytes in the nucleus tractus solitarius during ventilatory acclimatization to 10% hypoxia in unanesthetized mice. J. Neurosci. Res. 2014, 92, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Liu, Z.; Yan, X.; Luo, C. Effect of intermittent hypoxia on neuro-functional recovery post brain ischemia in mice. J. Mol. Neurosci. 2015, 55, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.J. The role of microglia in neurotoxicity. In The Role of Glia in Neurotoxicity; Aschner, M., Costa, L.G., Eds.; CRC Press: Boca Raton, FL, USA, 1996; pp. 29–40. [Google Scholar]
- Graeber, M.B.; Li, W.; Rodriguez, M.L. Role of microglia in CNS inflammation. FEBS Lett. 2011, 585, 3798–3805. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A.; et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef]
- Rodriguez, J.I.; Kern, J.K. Evidence of microglial activation in autism and its possible role in brain underconnectivity. Neuron. Glia. Biol. 2011, 7, 205–213. [Google Scholar] [CrossRef]
- Lyman, M.; Lloyd, D.G.; Ji, X.; Vizcaychipi, M.P.; Ma, D. Neuroinflammation: The role and consequences. Neurosci. Res. 2014, 79, 1–12. [Google Scholar] [CrossRef]
- Bachstetter, A.D.; Xing, B.; de Almeida, L.; Dimayuga, E.R.; Watterson, D.M.; Van Eldik, L.J. Microglial p38α MAPK is a key regulator of proinflammatory cytokine up-regulation induced by toll-like receptor (TLR) ligands or beta-amyloid (Aβ). J. Neuroinflammation 2011, 8, 79. [Google Scholar] [CrossRef]
- Hirsch, E.C.; Vyas, S.; Hunot, S. Neuroinflammation in Parkinson’s disease. Parkinsonism Relat. Disord. 2012, 18 (Suppl. 1), S210–S212. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 2005, 308, 1314–8131. [Google Scholar] [CrossRef]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Chen, Z.; Wang, P.; Zhao, M.; Fujino, M.; Zhang, C.; Zhou, W.; Hirano, S.I.; Li, X.K.; Zhao, L. Hydrogen inhalation protects hypoxic-ischemic brain damage by attenuating inflammation and apoptosis in neonatal rats. Exp. Biol. Med. 2019, 244, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Rong, P.; Zhang, L.; He, H.; Zhou, T.; Fan, Y.; Mo, L.; Zhao, Q.; Han, Y.; Li, S.; et al. IL4-driven microglia modulate stress resilience through BDNF-dependent neurogenesis. Sci. Adv. 2021, 7, eabb9888. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Barhwal, K.K.; Biswal, S.N.; Srivastava, A.K.; Bhardwaj, P.; Kumar, A.; Chaurasia, O.P.; Hota, S.K. Hypoxia-mediated alteration in cholesterol oxidation and raft dynamics regulates BDNF signalling and neurodegeneration in hippocampus. J. Neurochem. 2019, 148, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Kerschensteiner, M.; Stadelmann, C.; Dechant, G.; Wekerle, H.; Hohlfeld, R. Neurotrophic cross-talk between the nervous and immune systems: Implications for neurological diseases. Ann. Neurol. 2003, 53, 292–304. [Google Scholar] [CrossRef]
- Madinier, A.; Bertrand, N.; Mossiat, C.; Prigent-Tessier, A.; Beley, A.; Marie, C.; Garnier, P. Microglial involvement in neuroplastic changes following focal brain ischemia in rats. PLoS ONE 2009, 4, e8101. [Google Scholar] [CrossRef]
- Sun, P.; Li, X.; Chen, C.; Chen, Q.; Ouyang, Q.; Liu, F.; Xiang, Z.; Yuan, H. Activating transcription factor 4 modulates BDNF release from microglial cells. J. Mol. Neurosci. 2014, 52, 225–230. [Google Scholar] [CrossRef]
- Huang, Y.; Cheung, L.; Rowe, D.; Halliday, G. Genetic contributions to Parkinson’s disease. Brain Res. Brain Res. Rev. 2004, 46, 44–70. [Google Scholar] [CrossRef]
- Imai, F.; Suzuki, H.; Oda, J.; Ninomiya, T.; Ono, K.; Sano, H.; Sawada, M. Neuroprotective effect of exogenous microglia in global brain ischemia. J. Cereb. Blood. Flow. Metab. 2007, 27, 488–500. [Google Scholar] [CrossRef]
- Tansey, M.G.; Goldberg, M.S. Neuroinflammation in Parkinson’s disease: Its role in neuronal death and implications for therapeutic intervention. Neurobiol. Dis. 2010, 37, 510–518. [Google Scholar] [CrossRef]
- Chavez-Valdez, R.; Mason, A.; Nunes, A.R.; Northington, F.J.; Tankersley, C.; Ahlawat, R.; Johnson, S.M.; Gauda, E.B. Effect of hyperoxic exposure during early development on neurotrophin expression in the carotid body and nucleus tractus solitarii. J. Appl. Physiol. 2012, 112, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Montero, S.; Cuéllar, R.; Lemus, M.; Avalos, R.; Ramírez, G.; de Álvarez-Buylla, E.R. Brain-derived neurotrophic factor in the nucleus tractus solitarii modulates glucose homeostasis after carotid chemoreceptor stimulation in rats. Adv. Exp. Med. Biol. 2012, 758, 233–239. [Google Scholar] [PubMed]
- Thibault, K.; Lin, W.K.; Rancillac, A.; Fan, M.; Snollaerts, T.; Sordoillet, V.; Hamon, M.; Smith, G.M.; Lenkei, Z.; Pezet, S. BDNF-dependent plasticity induced by peripheral inflammation in the primary sensory and the cingulate cortex triggers cold allodynia and reveals a major role for endogenous BDNF as a tuner of the affective aspect of pain. J. Neurosci. 2014, 34, 14739–14751. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.J.; Yang, T.; Wei, X.; Liu, Y.; Xin, W.J.; Chen, Y.; Pang, R.P.; Zang, Y.; Li, Y.Y.; Liu, X.G. Brain-derived neurotrophic factor contributes to spinal long-term potentiation and mechanical hypersensitivity by activation of spinal microglia in rat. Brain Behav. Immun. 2011, 25, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Chen, J.; Su, M.; Lin, Z.; Zhan, H.; Yang, F.; Li, W.; Xie, J.; Huang, Y.; Liu, X.; et al. BDNF promotes activation of astrocytes and microglia contributing to neuroinflammation and mechanical allodynia in cyclophosphamide-induced cystitis. J. Neuroinflamm. 2020, 17, 19. [Google Scholar] [CrossRef]
- Infantino, R.; Schiano, C.; Luongo, L.; Paino, S.; Mansueto, G.; Boccella, S.; Guida, F.; Ricciardi, F.; Iannotta, M.; Belardo, C.; et al. MED1/BDNF/TrkB pathway is involved in thalamic hemorrhage-induced pain and depression by regulating microglia. Neurobiol. Dis. 2022, 164, 105611. [Google Scholar] [CrossRef]
- Nakajima, K.; Kikuchi, Y.; Ikoma, E.; Honda, S.; Ishikawa, M.; Liu, Y.; Kohsaka, S. Neurotrophins regulate the function of cultured microglia. Glia 1998, 24, 272–289. [Google Scholar] [CrossRef]
- Tanaka, K.; Okugawa, Y.; Toiyama, Y.; Inoue, Y.; Saigusa, S.; Kawamura, M.; Araki, T.; Uchida, K.; Mohri, Y.; Kusunoki, M. Brain-derived neurotrophic factor (BDNF)-induced tropomyosin-related kinase B (Trk B) signaling is a potential therapeutic target for peritoneal carcinomatosis arising from colorectal cancer. PLoS ONE 2014, 9, e96410. [Google Scholar] [CrossRef]
- Thielen, R.J.; Engleman, E.A.; Rodd, Z.A.; Murphy, J.M.; Lumeng, L.; Li, T.K.; McBride, W.J. Ethanol drinking and deprivation alter dopaminergic and serotonergic function in the nucleus accumbens of alcohol-preferring rats. J. Pharmacol. Exp. Ther. 2004, 309, 216–225. [Google Scholar] [CrossRef]
- Valvassori, S.S.; Arent, C.O.; Steckert, A.V.; Varela, R.B.; Jornada, L.K.; Tonin, P.T.; Budni, J.; Mariot, E.; Kapczinski, F.; Quevedo, J. Intracerebral Administration of BDNF Protects Rat Brain against Oxidative Stress Induced by Ouabain in an Animal Model of Mania. Mol. Neurobiol. 2015, 52, 353–362. [Google Scholar] [CrossRef]
- Fukuhara, K.; Ishikawa, K.; Yasuda, S.; Kishishita, Y.; Kim, H.K.; Kakeda, T.; Yamamoto, M.; Norii, T.; Ishikawa, T. Intracerebroventricular 4-methylcatechol (4-MC) ameliorates chronic pain associated with depression-like behavior via induction of brain-derived neurotrophic factor (BDNF). Cell. Mol. Neurobiol. 2012, 32, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic Press: New York, NY, USA, 1986. [Google Scholar]
- Streit, W.J.; Schulte, B.A.; Balentine, D.J.; Spicer, S.S. Histochemical localization of galactose-containing glycoconjugates in sensory neurons and their processes in the central and peripheral nervous system of the rat. J. Histochem. Cytochem. 1985, 33, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.J.; Kreutzberg, G.W. Lectin binding by resting and reactive microglia. J. Neurocytol. 1987, 16, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Hatton, W.J.; von Bartheld, C.S. Analysis of cell death in the trochlear nucleus of the chick embryo: Calibration of the optical disector counting method reveals systematic bias. J. Comp. Neurol. 1999, 409, 169–186. [Google Scholar] [CrossRef]
- Kaplan, S.; Odaci, E.; Canan, S.; Onger, M.E.; Aslan, H.; Unal, B. The Disector counting technique. NeuroQuantology 2012, 10, 44–53. [Google Scholar] [CrossRef]
- Diz-Chaves, Y.; Pernía, O.; Carrero, P.; Garcia-Segura, L.M. Prenatal stress causes alterations in the morphology of microglia and the inflammatory response of the hippocampus of adult female mice. J. Neuroinflamm. 2012, 9, 71. [Google Scholar] [CrossRef]
- Clark, C.G.; Hasser, E.M.; Kunze, D.L.; Katz, D.M.; Kline, D.D. Endogenous brain-derived neurotrophic factor in the nucleus tractus solitarius tonically regulates synaptic and autonomic function. J. Neurosci. 2011, 31, 12318–12329. [Google Scholar] [CrossRef]
- Mizoguchi, Y.; Kato, T.A.; Seki, Y.; Ohgidani, M.; Sagata, N.; Horikawa, H.; Yamauchi, Y.; Sato-Kasai, M.; Hayakawa, K.; Inoue, R.; et al. Brain-derived neurotrophic factor (BDNF) induces sustained intracellular Ca2+ elevation through the up-regulation of surface transient receptor potential 3 (TRPC3) channels in rodent microglia. J. Biol. Chem. 2014, 289, 18549–18555. [Google Scholar] [CrossRef]
- Barbacid, M. The Trk family of neurotrophin receptors. J. Neurobiol. 1994, 25, 1386–1403. [Google Scholar] [CrossRef]
- Wang, Y.; Hagel, C.; Hamel, W.; Müller, S.; Kluwe, L.; Westphal, M. Trk A, B, and C are commonly expressed in human astrocytes and astrocytic gliomas but not by human oligodendrocytes and oligodendroglioma. Acta. Neuropathol. 1998, 96, 357–364. [Google Scholar] [CrossRef]
- Spencer-Segal, J.L.; Waters, E.M.; Bath, K.G.; Chao, M.V.; McEwen, B.S.; Milner, T.A. Distribution of phosphorylated TrkB receptor in the mouse hippocampal formation depends on sex and estrous cycle stage. J. Neurosci. 2011, 31, 6780–6790. [Google Scholar] [CrossRef] [PubMed]
- Geller, R.J.; Barthold, C.; Saiers, J.A.; Hall, A.H. Pediatric cyanide poisoning: Causes, manifestations, management, and unmet needs. Pediatrics 2006, 118, 2146–2158. [Google Scholar] [CrossRef] [PubMed]
- Hawk, M.A.; Ritchie, G.D.; Henderson, K.A.; Knostman, K.A.; Roche, B.M.; Ma, Z.J.; Matthews, C.M.; Sabourin, C.L.; Wakayama, E.J.; Sabourin, P.J. Neurobehavioral and Cardiovascular Effects of Potassium Cyanide Administered Orally to Mice. Int. J. Toxicol. 2016, 35, 604–615. [Google Scholar] [CrossRef]
- Cuéllar, R.; Montero, S.; Luquín, S.; García-Estrada, J.; Dobrovinskaya, O.; Melnikov, V.; Lemus, M.; de Álvarez-Buylla, E.R. Glutamatergic Receptor Activation in the Commisural Nucleus Tractus Solitarii (cNTS) Mediates Brain Glucose Retention (BGR) Response to Anoxic Carotid Chemoreceptor (CChr) Stimulation in Rats. Adv. Exp. Med. Biol. 2015, 860, 301–308. [Google Scholar] [PubMed]
- Cuéllar, R.; Montero, S.; Luquín, S.; García-Estrada, J.; Melnikov, V.; Virgen-Ortiz, A.; Lemus, M.; Pineda-Lemus, M.; de Álvarez-Buylla, E. BDNF and AMPA receptors in the cNTS modulate the hyperglycemic reflex after local carotid body NaCN stimulation. Auton. Neurosci. 2017, 205, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Merlio, J.P.; Ernfors, P.; Jaber, M.; Persson, H. Molecular cloning of rat trkC and distribution of cells expressing messenger RNAs for members of the trk family in the rat central nervous system. Neuroscience 1992, 51, 513–532. [Google Scholar] [CrossRef]
- Yan, Q.; Matheson, C.; Sun, J.; Radeke, M.J.; Feinstein, S.C.; Miller, J.A. Distribution of intracerebral ventricularly administered neurotrophins in rat brain and its correlation with trk receptor expression. Exp. Neurol. 1994, 127, 23–36. [Google Scholar] [CrossRef]
- Croll, S.D.; Chesnutt, C.R.; Rudge, J.S.; Acheson, A.; Ryan, T.E.; Siuciak, J.A.; DiStefano, P.S.; Wiegand, S.J.; Lindsay, R.M. Co-infusion with a TrkB-Fc receptor body carrier enhances BDNF distribution in the adult rat brain. Exp. Neurol. 1998, 152, 20–33. [Google Scholar] [CrossRef]
- Rudge, J.S.; Li, Y.; Pasnikowski, E.M.; Mattsson, K.; Pan, L.; Yancopoulos, G.D.; Wiegand, S.J.; Lindsay, R.M.; Ip, N.Y. Neurotrophic factor receptors and their signal transduction capabilities in rat astrocytes. Eur. J. Neurosci. 1994, 6, 693–705. [Google Scholar] [CrossRef]
- Ju, J.; Chen, R.; Li, T.; Wang, J.; Xiao, Y.; Liu, C. Modulation of hippocampal synaptic transmission in a mouse model of chronic intermittent hypoxia: Implications for neurocognitive impairments. Zhonghua Yi Xue Za Zhi. 2016, 96, 610–614. [Google Scholar]
- Fanselow, M.S.; Dong, H.W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Gozal, D.; Daniel, J.M.; Dohanich, G.P. Behavioral and anatomical correlates of chronic episodic hypoxia during sleep in the rat. J. Neurosci. 2001, 21, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Bucks, R.S.; Olaithe, M.; Eastwood, P. Neurocognitive function in obstructive sleep apnoea: A meta-review. Respirology 2013, 18, 61–70. [Google Scholar] [CrossRef]
- Torelli, F.; Moscufo, N.; Garreffa, G.; Placidi, F.; Romigi, A.; Zannino, S.; Bozzali, M.; Fasano, F.; Giulietti, G.; Djonlagic, I.; et al. Cognitive profile and brain morphological changes in obstructive sleep apnea. Neuroimage 2011, 54, 787–793. [Google Scholar] [CrossRef]
- Ahn, J.H.; Shin, B.N.; Park, J.H.; Kim, I.H.; Cho, J.H.; Chen, B.; Lee, T.K.; Tae, H.J.; Lee, J.C.; Cho, J.H.; et al. Long-term observation of neuronal degeneration and microgliosis in the gerbil dentate gyrus after transient cerebral ischemia. J. Neurol. Sci. 2016, 363, 21–26. [Google Scholar] [CrossRef]
- Dougherty, K.D.; Dreyfus, C.F.; Black, I.B. Brain-derived neurotrophic factor in astrocytes, oligodendrocytes, and microglia/macrophages after spinal cord injury. Neurobiol. Dis. 2000, 7, 574–585. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, S.R.; Choi, S.S.; Yeo, H.G.; Chang, K.T.; Lee, H.J. Therapeutically targeting neuroinflammation and microglia after acute ischemic stroke. Biomed. Res. Int. 2014, 2014, 297241. [Google Scholar] [CrossRef]
- Chamberland, S.; Topolnik, L. Inhibitory control of hippocampal inhibitory neurons. Front. Neurosci. 2012, 6, 165. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Kunimatsu, T.; Yamada, K.; Kojo, A.; Yamamoto, T.; Sato, S.; Onozuka, M. Hypothermic and normothermic ischemia-reperfusion activate microglia differently in hippocampal formation. Arch. Histol. Cytol. 2010, 73, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Cervós-Navarro, J.; Diemer, N.H. Selective vulnerability in brain hypoxia. Crit. Rev. Neurobiol. 1991, 6, 149–182. [Google Scholar] [PubMed]
- Blanquet, P.R.; Mariani, J.; Fournier, B. Identification of a biphasic signaling pathway involved in ischemic resistance of the hippocampal dentate gyrus. Exp. Neurol. 2006, 202, 357–372. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Waterhouse, B. Locus coeruleus: From global projection system to adaptive regulation of behavior. Brain Res. 2016, 1645, 75–78. [Google Scholar] [CrossRef]
- Bae, E.; Hwang, I.K.; Yoo, K.Y.; Han, T.H.; Lee, C.H.; Choi, J.H.; Yi, S.S.; Lee, S.Y.; Ryu, P.D.; Yoon, Y.S.; et al. Gliosis in the amygdala following myocardial infarction in the rat. J. Vet. Med. Sci. 2010, 72, 1041–1045. [Google Scholar] [CrossRef]
- Lee, T.H.; Kato, H.; Chen, S.T.; Kogure, K.; Itoyama, Y. Expression disparity of brain-derived neurotrophic factor immunoreactivity and mRNA in ischemic hippocampal neurons. Neuroreport 2002, 13, 2271–2275. [Google Scholar] [CrossRef]
- Coldren, K.M.; Li, D.P.; Kline, D.D.; Hasser, E.M.; Heesch, C.M. Acute hypoxia activates neuroendocrine, but not presympathetic, neurons in the paraventricular nucleus of the hypothalamus: Differential role of nitric oxide. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R982–R995. [Google Scholar] [CrossRef]
- Wang, C.; Bomberg, E.; Billington, C.; Levine, A.; Kotz, C.M. Brain-derived neurotrophic factor in the hypothalamic paraventricular nucleus reduces energy intake. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1003–R1012. [Google Scholar] [CrossRef]
- Teubner, B.J.; Leitner, C.; Thomas, M.A.; Ryu, V.; Bartness, T.J. An intact dorsomedial posterior arcuate nucleus is not necessary for photoperiodic responses in Siberian hamsters. Horm. Behav. 2015, 70, 22–29. [Google Scholar] [CrossRef]
- Streit, W.J.; Mrak, R.E.; Griffin, W.S. Microglia and neuroinflammation: A pathological perspective. J. Neuroinflammation 2004, 1, 14. [Google Scholar] [CrossRef]
- Kreutzberg, G.W. Microglia: A sensor for pathological events in the CNS. Trends Neurosci. 1996, 19, 312–318. [Google Scholar] [CrossRef]
- Miwa, T.; Furukawa, S.; Nakajima, K.; Furukawa, Y.; Kohsaka, S. Lipopolysaccharide enhances synthesis of brain-derived neurotrophic factor in cultured rat microglia. J. Neurosci. Res. 1997, 50, 1023–1029. [Google Scholar] [CrossRef]
- Ulmann, L.; Hatcher, J.P.; Hughes, J.P.; Chaumont, S.; Green, P.J.; Conquet, F.; Buell, G.N.; Reeve, A.J.; Chessell, I.P.; Rassendren, F. Up-regulation of P2X4 receptors in spinal microglia after peripheral nerve injury mediates BDNF release and neuropathic pain. J. Neurosci. 2008, 28, 11263–11268. [Google Scholar] [CrossRef]
- Fujita, R.; Ma, Y.; Ueda, H. Lysophosphatidic acid-induced membrane ruffling and brain-derived neurotrophic factor gene expression are mediated by ATP release in primary microglia. J. Neurochem. 2008, 107, 152–160. [Google Scholar] [CrossRef]
- Trang, T.; Beggs, S.; Wan, X.; Salter, M.W. P2X4-receptor-mediated synthesis and release of brain-derived neurotrophic factor in microglia is dependent on calcium and p38-mitogen-activated protein kinase activation. J. Neurosci. 2009, 29, 3518–3528. [Google Scholar] [CrossRef]
- Sastre, M.; Richardson, J.C.; Gentleman, S.M.; Brooks, D.J. Inflammatory risk factors and pathologies associated with Alzheimer’s disease. Curr. Alzheimer. Res. 2011, 8, 132–141. [Google Scholar] [CrossRef]
- Merlo, S.; Luaces, J.P.; Spampinato, S.F.; Toro-Urrego, N.; Caruso, G.I.; D’Amico, F.; Capani, F.; Sortino, M.A. SIRT1 Mediates Melatonin’s Effects on Microglial Activation in Hypoxia: In Vitro and In Vivo Evidence. Biomolecules 2020, 10, 364. [Google Scholar] [CrossRef]
- Seo, J.W.; Kim, J.H.; Kim, J.H.; Seo, M.; Han, H.S.; Park, J.; Suk, K. Time-dependent effects of hypothermia on microglial activation and migration. J. Neuroinflammation 2012, 9, 164. [Google Scholar] [CrossRef]
- Nakajima, K.; Honda, S.; Tohyama, Y.; Imai, Y.; Kohsaka, S.; Kurihara, T. Neurotrophin secretion from cultured microglia. J. Neurosci. Res. 2001, 65, 322–331. [Google Scholar] [CrossRef]
- Mizoguchi, Y.; Monji, A.; Kato, T.; Seki, Y.; Gotoh, L.; Horikawa, H.; Suzuki, S.O.; Iwaki, T.; Yonaha, M.; Hashioka, S.; et al. Brain-derived neurotrophic factor induces sustained elevation of intracellular Ca2+ in rodent microglia. J. Immunol. 2009, 183, 7778–7786. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (1) Chemical Union of GRIFFONIA to Glycocalix. | (2) Incubation with Primary Antibody (Ab). | (3) Incubation with Secondary Ab. | (4) Incubation with Avidin–Biotin Complex (ABC) |
|---|---|---|---|
| Griffonia simplicifolia Lectin I non-conjugated goat isolectin B4 1:100 | Anti-griffonia (Bandeiraea) simplicifolia lectin goat 1:100 | Biotinylated rabbit anti-goat IgG 1:150 | Sandwich-type reaction and color development in chromogenic solution |
| Area | |||
|---|---|---|---|
| Drug | BLA | HDG | Arc |
| aCSF | 7209 ± 488 * | 6882 ± 706 * | 5647 ± 873 * |
| BDNF | 6095 ± 408 * | 3694 ± 491 # | 5012 ± 621 |
| K252a | 3556 ± 536 | 2241 ± 433 # | 3288 ± 450 # |
| Sham-operated | 2743 ± 711 | 3879 ± 927 | 3048 ± 880 * |
| Area | |||
|---|---|---|---|
| Drug | BLA | HDG | Arc |
| aCSF | II (58%) | III (36%) | II (54%) |
| BDNF | I, II, III (29% each) | III (42%) | II (46%) |
| K252a | II (58%) | II, III (38% each) | II (67%) |
| Sham-operated | I (89%) | I (57%) | I (57%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuéllar-Pérez, R.; Jauregui-Huerta, F.; Ruvalcaba-Delgadillo, Y.; Montero, S.; Lemus, M.; Roces de Álvarez-Buylla, E.; García-Estrada, J.; Luquín, S. K252a Prevents Microglial Activation Induced by Anoxic Stimulation of Carotid Bodies in Rats. Toxics 2023, 11, 871. https://doi.org/10.3390/toxics11100871
Cuéllar-Pérez R, Jauregui-Huerta F, Ruvalcaba-Delgadillo Y, Montero S, Lemus M, Roces de Álvarez-Buylla E, García-Estrada J, Luquín S. K252a Prevents Microglial Activation Induced by Anoxic Stimulation of Carotid Bodies in Rats. Toxics. 2023; 11(10):871. https://doi.org/10.3390/toxics11100871
Chicago/Turabian StyleCuéllar-Pérez, Ricardo, Fernando Jauregui-Huerta, Yaveth Ruvalcaba-Delgadillo, Sergio Montero, Mónica Lemus, Elena Roces de Álvarez-Buylla, Joaquín García-Estrada, and Sonia Luquín. 2023. "K252a Prevents Microglial Activation Induced by Anoxic Stimulation of Carotid Bodies in Rats" Toxics 11, no. 10: 871. https://doi.org/10.3390/toxics11100871