Multiplex Recombinase Polymerase Amplification Assay for the Simultaneous Detection of Three Foodborne Pathogens in Seafood


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and DNA Template Preparation
2.2. Design of the RPA Primers
2.3. Multiplex RPA Reactions in Solution
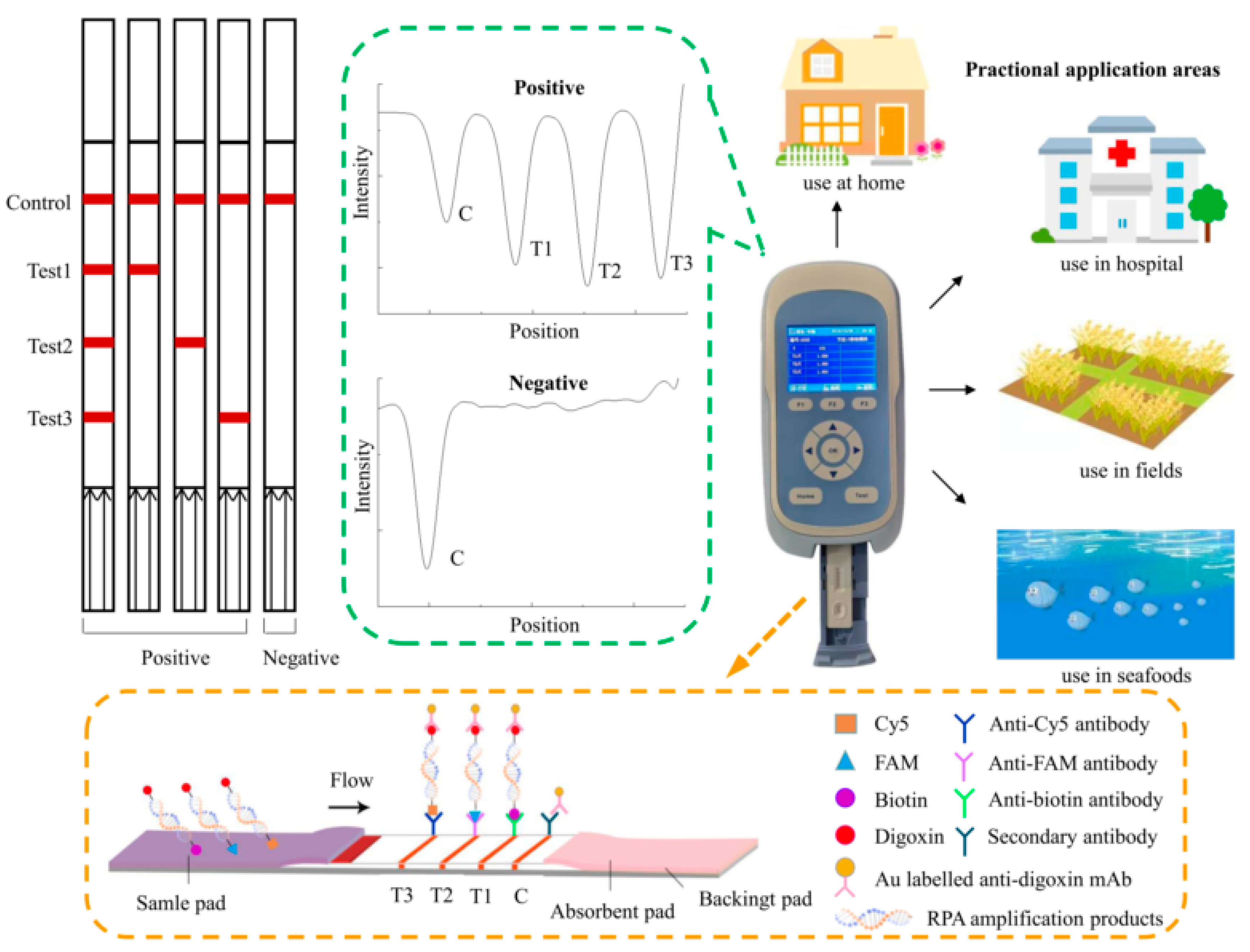
2.4. Visualization and Quantification of Lateral Flow Dipsticks
2.5. RPA-LFD Assay
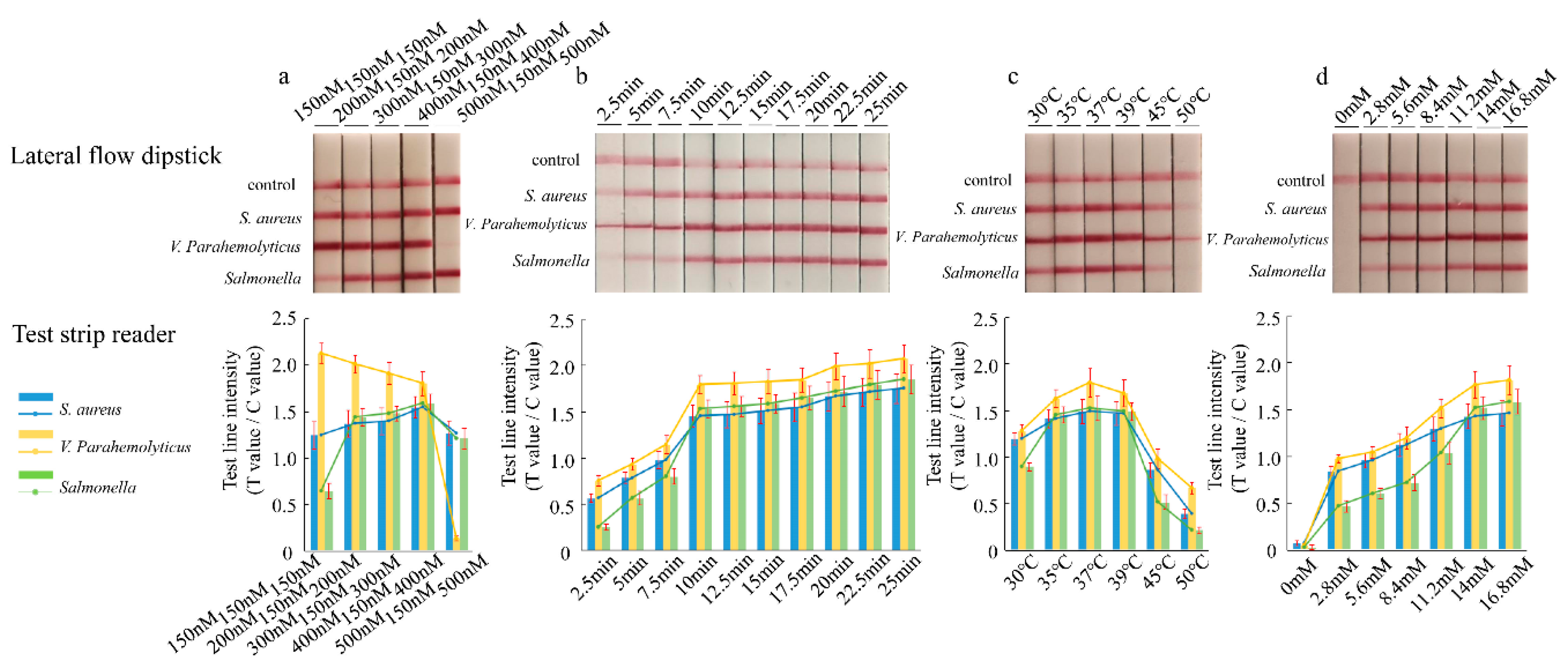
2.6. Optimization of the LFD-RPA Conditions
2.7. Specificity and Sensitivity of Multiplex LFD-RPA
2.8. Application of Multiplex LFD-RPA in Artificially Contaminated Food Samples
2.9. Field Sample Testing
2.10. Data Analysis
3. Results
3.1. Establishment and Optimization of the Multiplex LFD-RPA Assay
3.2. Sensitivity and Specificity of the Multiplex LFD-RPA Assay
3.3. Application of the Multiplex LFD-RPA Assay in Food Samples
3.4. Detection of Field Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, H.B.; Du, X.J.; Zang, Y.X.; Li, P.; Wang, S. SERS-based lateral flow strip biosensor for simultaneous detection of Listeria monocytogenes and Salmonella enterica serotype enteritidis. J. Agric. Food Chem. 2017, 65, 10290–10299. [Google Scholar] [CrossRef]
- Simon, S.S.; Sanjeev, S. Prevalence of enterotoxigenic staphylococcus aureus in fishery products and fish processing factory workers. Food Control 2007, 18, 1565–1568. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Xiao, L.L.; Lou, Y.; Jin, M.T.; Liao, C.; Malakar, P.K.; Pan, Y.J.; Zhao, Y. Development of a multiplex real-time PCR method for simultaneous detection of Vibrio parahaemolyticus, Listeria monocytogenes and Salmonella spp. in raw shrimp. Food Control 2015, 51, 31–36. [Google Scholar] [CrossRef]
- Geng, Y.Y.; Liu, S.Y.; Wang, J.F.; Nan, H.Z.; Liu, L.B.; Sun, X.X.; Li, D.Y.; Liu, M.; Wang, J.C.; Tan, K. Rapid detection of Staphylococcus aureus in food using a recombinase polymerase amplification-based assay. Food Anal. Methods 2018, 11, 2847–2856. [Google Scholar] [CrossRef]
- Du, X.J.; Zang, Y.X.; Liu, H.B.; Li, P.; Wang, S. Rapid Detection of Staphylococcus aureus via recombinase polymerase amplification combined with lateral flow strip. Food Anal. Methods 2018, 11, 2296–2306. [Google Scholar] [CrossRef]
- Butt, A.A.; Aldridge, K.E.; Sanders, C.V. Infections related to the ingestion of seafood part I: viral and bacterial infections. Lancet Infect. Dis. 2004, 4, 201–212. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Z.; Wang, Y.; Zhao, Y.; Lu, Y.; Xu, X.; Yan, J.; Pan, Y. A highly sensitive and flexible magnetic nanoprobe labeled immunochromatographic assay platform for pathogen Vibrio parahaemolyticus. Int. J. Food Microbiol. 2015, 211, 109–116. [Google Scholar] [CrossRef]
- Mąka, Ł.; Popowska, M. Antimicrobial resistance of Salmonella spp. isolated from food. Rocz. Państwowego Zakładu Hig. 2016, 67, 343–358. [Google Scholar]
- Li, F.; Li, F.L.; Chen, B.L.; Zhou, B.Q.; Yu, P.; Yu, S.; Lai, W.H.; Xu, H.Y. Sextuplex PCR combined with immunomagnetic separation and PMA treatment for rapid detection and specific identification of viable Salmonella spp., Salmonella Enterica serovars Paratyphi B, Salmonella Typhimurium, and Salmonella Enteritidis in raw meat. Food Control 2017, 73, 587–594. [Google Scholar] [CrossRef]
- Gao, W.; Huang, H.; Zhu, P.; Yan, X.; Fan, J.; Jiang, J.; Xu, J. Recombinase polymerase amplification combined with lateral flow dipstick for equipment-free detection of Salmonella in shellfish. Bioprocess Biosyst. Eng. 2018, 41, 603–611. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, H.; Zhang, P.; Sun, C.; Wang, X.; Wang, X.; Yang, R.; Wang, C.; Zhou, L. Rapid multiplex detection of 10 foodborne pathogens with an up-converting phosphor technology-based 10-channel lateral flow assay. Sci. Rep. 2016, 6, 21342. [Google Scholar] [CrossRef] [PubMed]
- Whiley, H.; Taylor, M.; Bentham, R. Detection of Legionella species in potting mixes using fluorescent in situ hybridisation (FISH). J. Microbiol. Methods 2011, 86, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, H.; Wang, Y.; Zhang, L.; Xu, J.; Ye, C. Loop-Mediated isothermal amplification label-based gold nanoparticles lateral flow biosensor for detection of Enterococcus faecalis and Staphylococcus aureus. Front. Microbiol. 2017, 8, 192. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Kim, M.J.; Kim, H.J.; Jekong, K.C.; Kim, H.Y. Simultaneous detection of four foodborne viruses in food samples using a one-step multiplex reverse transcription PCR. J. Microbiol. Biotechnol. 2018, 28, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Tao, T.T.; Zhu, X.Y.; Bie, X.M.; Lv, F.X.; Zhao, H.Z.; Lu, Z.X. A multiplex PCR detection method for milk based on novel primers specific for Listeria monocytogenes 1/2a serotype. Food Control 2018, 86, 183–190. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4, 204. [Google Scholar] [CrossRef]
- Dao, T.N.T.; Lee, E.Y.; Koo, B.; Jin, C.E.; Lee, T.Y.; Shin, Y. A microfluidic enrichment platform with a recombinase polymerase amplification sensor for pathogen diagnosis. Anal. Biochem. 2018, 544, 87–92. [Google Scholar] [CrossRef]
- Ma, B.; Fang, J.; Wang, Y.; He, H.; Dai, M.; Lin, W.; Su, W.; Zhang, M. Isothermal method of a recombinase polymerase amplification assay for the detection of most common high-risk human papillomavirus type 16 and type 18 DNA. Clin. Lab. 2017, 63, 27–38. [Google Scholar] [CrossRef]
- Dzantiev, B.B.; Byzova, N.A.; Urusov, A.E.; Zherdev, A.V. Immunochromatographic methods in food analysis. Trends Anal. Chem. 2014, 55, 81–93. [Google Scholar] [CrossRef]
- Kersting, S.; Rausch, V.; Bier, F.F.; von Nickisch-Rosenegk, M. Multiplex isothermal solid-phase recombinase polymerase amplification for the specific and fast DNA-based detection of three bacterial pathogens. Mikrochim. Acta 2014, 181, 1715–1723. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Yu, X.Z.; Wen, K.; Li, C.L.; Marti, G.M.; Jiang, H.Y.; Shi, W.M.; Shen, J.Z.; Wang, Z.H. Multiplex Lateral Flow Immunoassays based on amorphous carbon nanoparticles for detecting three Fusarium Mycotoxins in Maize. J. Agric. Food Chem. 2017, 65, 8063–8071. [Google Scholar] [CrossRef]
- Zhao, G.; Wang, H.; Hou, P.; He, C.; He, H. Rapid visual detection of Mycobacterium avium subsp. paratuberculosis by recombinase polymerase amplification combined with a lateral flow dipstick. J. Vet. Sci. 2018, 19, 242–250. [Google Scholar] [CrossRef]
- Fuller, S.L.; Savory, E.A.; Weisberg, A.J.; Buser, J.Z.; Gordon, M.I.; Putnam, M.L.; Chang, J.H. Isothermal amplification and lateral flow assay for detecting crown gall-causing Agrobacterium spp. Phytopathology 2017, 107, 1062–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, X.J.; Zang, Y.X.; Liu, H.B.; Li, P.; Wang, S. Recombinase polymerase amplification combined with lateral flow strip for Listeria monocytogenes detection in food. J. Food Sci. 2018, 83, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Kersting, S.; Rausch, V.; Bier, F.F.; von Nickisch-Rosenegk, M. A recombinase polymerase amplification assay for the diagnosis of atypical pneumonia. Anal. Biochem. 2018, 550, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Bacteriological Analytical Manual. Available online: https://www.fda.gov/food/laboratory-methods-food/bacteriological-analytical-manual-bam (accessed on 1 February 2020).
- Xu, D.; Ji, L.; Wu, X.; Yan, W.; Chen, L. Detection and differentiation of Vibrio parahaemolyticus by multiplexed real-time PCR. Can. J. Microbiol 2018, 64, 809–815. [Google Scholar] [CrossRef]
- Lv, X.; Huang, Y.; Liu, D.; Liu, C.; Shan, S.; Li, G.; Duan, M.; Lai, W. Multicolor and ultrasensitive enzyme-linked immunosorbent assay based on the fluorescence hybrid chain reaction for simultaneous detection of pathogens. J. Agric. Food Chem. 2019, 67, 9390–9398. [Google Scholar] [CrossRef]
- Ward, A.C.; Hannah, A.J.; Kendrick, S.L.; Tucker, N.P.; MacGregor, G.; Connolly, P. Identification and characterisation of Staphylococcus aureus on lowcost screen printed carbon electrodes using impedance spectroscopy. Biosens. Bioelectron. 2018, 110, 65–70. [Google Scholar] [CrossRef]
- Compton, J. Nucleic acid sequence-based amplification. Nature 1991, 350, 91–92. [Google Scholar] [CrossRef]
- Vincent, M.; Xu, Y.; Kong, H. Helicase-dependent isothermal DNA amplification. EMBO Rep. 2004, 5, 795–800. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Felipe, S.; Tortajada-Genaro, L.A.; Morais, S.; Puchades, R.; Maquieira, Á. Isothermal DNA amplification strategies for duplex microorganism detection. Food Chem. 2015, 174, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Cha, S.Y.; Kang, M.; Park, I.J.; Moon, O.K.; Park, C.K.; Jang, H.K. Development and application of a multiplex PCR assay for rapid detection of 4 major bacterial pathogens in ducks. Poult. Sci. 2013, 92, 1164–1170. [Google Scholar] [CrossRef] [PubMed]
- Nanayakkara, I.A.; White, I.M. Demonstration of a quantitative triplex LAMP assay with an improved probe-based readout for the detection of MRSA. Analyst 2019, 144, 3878–3885. [Google Scholar] [CrossRef] [PubMed]
- Reboud, J.; Xu, G.; Garrett, A.; Adriko, M.; Yang, Z.; Tukahebwa, E.M.; Rowell, C.; Cooper, J.M. Paper-based microfluidics for DNA diagnostics of malaria in low resource underserved rural communities. Proc. Natl. Acad. Sci. USA 2019, 116, 4834–4842. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Lee, J.L. Development of a multiplex real-time recombinase polymerase amplification (RPA) assay for rapid quantitative detection of Campylobacter coli and jejuni from eggs and chicken products. Food Control 2017, 73, 1247–1255. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, J.B.; Li, P.; Bai, L.; Jia, J.W.; Pan, A.H.; Long, X.Q.; Cui, W.D.; Tang, X.M. Rapid detection of P-35S and T-nos in genetically modified organisms by recombinase polymerase amplification combined with a lateral flow strip. Food Control 2020, 107, 106775. [Google Scholar] [CrossRef]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: basics, applications and recent advances. Trends Anal. Chem. 2018, 98, 19–35. [Google Scholar] [CrossRef]
- Croci, L.; Suffredini, E.; Cozzi, L.; Paniconi, M.; Ciccaglioni, G.; Colombo, M.M. Evaluation of different polymerase chain reaction methods for the identification of Vibrio parahaemolyticus strains isolated by cultural methods. J. AOAC Int. 2007, 90, 1588–1597. [Google Scholar] [CrossRef] [Green Version]
- Yeh, K.S.; Chen, T.H.; Liao, C.W.; Chang, C.S.; Lo, H.C. PCR amplification of the Salmonella typhimurium fimY gene sequence to detect the Salmonella species. Int. J. Food Microbiol. 2002, 78, 227–234. [Google Scholar] [CrossRef]
- Li, J.; Macdonald, J.; von Stetten, F. Review: A comprehensive summary of a decade development of the recombinase polymerase amplification. Analyst 2018, 144, 31–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauset-Rubio, M.; Tomaso, H.; El-Shahawi, M.S.; Bashammakh, A.S.; Al-Youbi, A.O.; O’Sullivan, C.K. Duplex lateral flow assay for the simultaneous detection of Yersinia pestis and Francisella tularensis. Anal. Chem. 2018, 90, 12745–12751. [Google Scholar] [CrossRef] [PubMed]
- Lillis, L.; Lehman, D.; Singhal, M.C.; Cantera, J.; Singleton, J.; Labarre, P.; Toyama, A.; Piepenburg, O.; Parker, M.; Wood, R.; et al. Non-instrumented incubation of a recombinase polymerase amplification assay for the rapid and sensitive detection of proviral HIV-1 DNA. PLoS ONE 2014, 9, e108189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Zhang, F.; Wang, L.; Qian, W.; Qian, C.; Wu, J.; Ying, Y. Instant, visual, and instrument-free method for on-site screening of GTS 40-3-2 soybean based on body-heat triggered recombinase polymerase amplification. Anal. Chem. 2017, 89, 4413–4418. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.M.; Wee, E.J.; Mainwaring, P.N.; Trau, M. A simple, rapid, low-cost technique for naked-eye detection of urine-isolated TMPRSS2: ERG gene fusion RNA. Sci. Rep. 2016, 6, 30722. [Google Scholar] [CrossRef] [Green Version]
- Kunze, A.; Dilcher, M.; Abd, E.; Wahed, A.; Hufert, F.; Niessner, R.; Seidel, M. On-chip isothermal nucleic acid amplification on flow-based chemiluminescence microarray analysis platform for the detection of viruses and bacteria. Anal. Chem. 2016, 88, 898–905. [Google Scholar] [CrossRef]
- Sabaté Del Río, J.; Steylaerts, T.; Henry, O.Y.F.; Bienstman, P.; Stakenborg, T.; Van Roy, W.; O’Sullivan, C.K. Real-time and label-free ring-resonator monitoring of solid-phase recombinase polymerase amplification. Biosens. Bioelectron. 2015, 73, 130–137. [Google Scholar] [CrossRef]
- Crannell, Z.; Castellanos-Gonzalez, A.; Nair, G.; Mejia, R.; White, A.C.; Richards-Kortum, R. Multiplexed recombinase polymerase amplification assay to detect intestinal protozoa. Anal. Chem. 2016, 88, 1610–1616. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | ID of Strains | Multiple LFD-RPA Results | ||
|---|---|---|---|---|
| nuc | toxR | fimY | ||
| Staphylococcus aureus | ATCC 25923 | + | - | - |
| Staphylococcus aureus | GIMCC 1.142 | + | - | - |
| Staphylococcus aureus | CICC 10001 | + | - | - |
| Staphylococcus aureus | CICC 21648 | + | - | - |
| Vibrio parahaemolyticus | ATCC 17802 | - | + | - |
| Vibrio parahaemolyticus | ATCC 33847 | - | + | - |
| Vibrio parahaemolyticus | H4-3 * | - | + | - |
| Vibrio parahaemolyticus | FJ14A * | - | + | - |
| Salmonella Enteritidis | ATCC 13076 | - | - | + |
| Salmonella Enteritidis | GIMCC 1.345 | - | - | + |
| Salmonella Enteritidis | CMCC 50041 | - | - | + |
| Salmonella enterica subsp. enterica | CICC 10982 | - | - | + |
| Legionella pneumophila | ATCC 33152 | - | - | - |
| Legionella pneumophila | 07 * | - | - | - |
| Vibrio cholera | GIMCC 1.449 | - | - | - |
| Vibrio cholera | 007zs0902-2009a | - | - | - |
| Escherichia coli O157:H7 | ATCC 35150 | - | - | - |
| Escherichia coli O157:H7 | 61 * | - | - | - |
| Shigella flexneri | CICC 10865 | - | - | - |
| Shigella sonnei | GIMCC 1.424 | - | - | - |
| Listeria monocytogenes | ATCC 19115 | - | - | - |
| Listeria monocytogene | CICC 21633 | - | - | - |
| Cronobacter Sakazakii | GIMCC 1.296 | - | - | - |
| Cronobacter Sakazakii | CS-3 * | - | - | - |
| Campylobacter jejuni subsp. jejuni | ATCC 33560 | - | - | - |
| Enterobacter aerogenes | CICC 10293 | - | - | - |
| Yersinia enterocolitica | ATCC 23715 | - | - | - |
| Target Name | Primer Name | Sequence (5′–3′) | Fragment Length |
|---|---|---|---|
| Staphylococcus aureus (nuc) | Forward primer | 5′-Biotin-CTTATAGGGATGGCTATCAGTAATGTTTCG-3′ | 158bp |
| Reverse primer | 5′-Digoxin-CCACTTCTATTTACGCCGTTATCTGTTTGT-3′ | ||
| Vibrio parahaemolyticus (toxR) | Forward primer | 5′-FAM-TTTGTTTGGCGTGAGCAAGGTTTTGAGGTG-3′ | 230bp |
| Reverse primer | 5′-Digoxin-GCAGAGGCGTCATTGTTATCAGAAGCAGGT-3′ | ||
| Salmonella enterica (fimY) | Forward primer | 5′-Cy5-TATCAGATAAAACCTCCGCTATAACACAGT-3′ | 133 bp |
| Reverse primer | 5′-Digoxin-CTTTCCGATAAGCGAGGTTTGGAGGCTGAT-3′ |
| Sample (n = 6 Each) | Staphylococcus aureus | Vibrio parahaemolyticus | Salmonella Enteritidis | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Inoculation Level (CFU/mL) | LFD-RPA Detected Concentration (CFU/mL) | Recovery (%) | BAM | Inoculation Level (CFU/mL) | LFD-RPA Detected Concentration (CFU/mL) | Recovery (%) | BAM | Inoculation Level (CFU/mL) | LFD-RPA Detected Concentration(CFU/mL) | Recovery (%) | BAM | |
| Sleevefish | 4.5 × 104 | 4.29 × 104 | 95.33 | + | 8.3 × 104 | 8.93 × 104 | 107.59 | + | 2.7 × 104 | 2.59 × 104 | 95.92 | + |
| 4.5 × 103 | 4.35 ×103 | 96.66 | + | 8.3 × 103 | 8.51 × 103 | 102.53 | + | 2.7 × 103 | 2.56 × 103 | 94.81 | + | |
| 4.5 × 102 | 4.66 × 102 | 103.56 | + | 8.3 × 102 | 8.13 × 102 | 98.95 | + | 2.7 × 102 | 2.61 × 102 | 96.6 | + | |
| 4.5 × 101 | 4.14 × 101 | 92.00 | - | 8.3 × 101 | 7.91 × 101 | 95.30 | - | 2.7 × 101 | 2.75 × 101 | 101.85 | - | |
| Shrimp | 4.5 × 104 | 4.72 × 104 | 104.89 | + | 8.3 × 104 | 8.15 × 104 | 98.19 | + | 2.7 × 104 | 2.78 × 104 | 102.96 | + |
| 4.5 × 103 | 4.42 × 103 | 98.22 | + | 8.3 × 103 | 8.67 × 103 | 104.46 | + | 2.7 × 103 | 2.53 × 103 | 93.70 | + | |
| 4.5 × 102 | 4.21 × 102 | 93.56 | + | 8.3 × 102 | 8.96 × 102 | 107.95 | + | 2.7 × 102 | 2.82 × 102 | 104.44 | + | |
| 4.5 × 101 | 4.15 × 101 | 92.22 | - | 8.3 × 101 | 7.96 × 101 | 95.90 | - | 2.7 × 101 | 2.56 × 101 | 94.81 | - | |
| Cod | 4.5 × 104 | 4.78 × 104 | 106.22 | + | 8.3 × 104 | 8.72 × 104 | 105.06 | + | 2.7 × 104 | 2.84 × 104 | 105.19 | + |
| 4.5 × 103 | 4.67 × 103 | 103.78 | + | 8.3 × 103 | 8.61 × 103 | 103.73 | + | 2.7 × 103 | 2.88 × 103 | 106.66 | + | |
| 4.5 × 102 | 4.36 × 102 | 96.89 | + | 8.3 × 102 | 8.06 × 102 | 97.11 | + | 2.7 × 102 | 2.67 × 102 | 98.88 | + | |
| 4.5 × 101 | 4.16 × 101 | 92.44 | - | 8.3 × 101 | 7.78 × 101 | 93.73 | - | 2.7 × 101 | 2.51 × 101 | 92.96 | - | |
| Resource | Samples | No. of Samples | Positive Number of Triplex LFD-RPA and BAM Method | |||||
|---|---|---|---|---|---|---|---|---|
| Staphylococcus aureus | Vibrio parahaemolyticus | Salmonella Enteritidis | ||||||
| Triplex LFD-RPA | BAM | Triplex LFD-RPA | BAM | Triplex LFD-RPA | BAM | |||
| Local market | Shrimp | 24 | 0 | 0 | 2 | 2 | 2 | 2 |
| Sleevefish | 21 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Trichiurus lepturus | 11 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Cod | 15 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Meretrix | 8 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Toasted Muraenesox | 6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Grilled fish | 5 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Grilled yellow croaker | 12 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Dried squid | 4 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total | 106 | 0 | 0 | 2 | 2 | 2 | 2 | |
| Positive detection rate (%) | / | 0% | 0% | 1.9% | 1.9% | 1.9% | 1.9% | |
| Zhoushan Entry-Exit Inspection and Quarantine Bureau | Dried squid | 17 | 0 | 0 | 0 | 0 | 0 | 0 |
| Positive detection rate (%) | / | 0% | 0% | 0% | 0% | 0% | 0% | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, B.; Li, J.; Chen, K.; Yu, X.; Sun, C.; Zhang, M. Multiplex Recombinase Polymerase Amplification Assay for the Simultaneous Detection of Three Foodborne Pathogens in Seafood. Foods 2020, 9, 278. https://doi.org/10.3390/foods9030278
Ma B, Li J, Chen K, Yu X, Sun C, Zhang M. Multiplex Recombinase Polymerase Amplification Assay for the Simultaneous Detection of Three Foodborne Pathogens in Seafood. Foods. 2020; 9(3):278. https://doi.org/10.3390/foods9030278
Chicago/Turabian StyleMa, Biao, Jiali Li, Kai Chen, Xiaoping Yu, Chuanxin Sun, and Mingzhou Zhang. 2020. "Multiplex Recombinase Polymerase Amplification Assay for the Simultaneous Detection of Three Foodborne Pathogens in Seafood" Foods 9, no. 3: 278. https://doi.org/10.3390/foods9030278