Quantification and Distribution of Omega-3 Fatty Acids in South Pacific Fish and Shellfish Species

 ,
,
Abstract
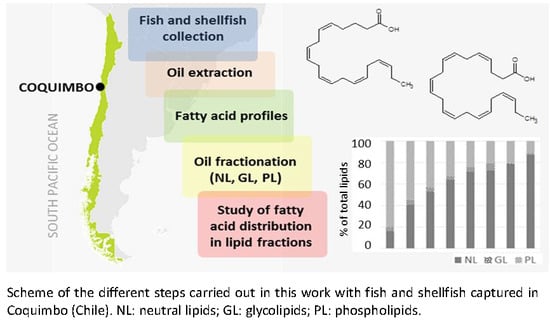
:
1. Introduction
2. Materials and Methods
2.1. Solvents and Reagents
2.2. Samples
2.3. Determination of Total Fat and Fatty Acid Quantification
2.4. Nutritional Quality Indexes of Lipids
2.5. Fatty Acid Distribution among Lipid Classes
2.6. Statistical Analysis
3. Results and Discussion
3.1. Lipid Amount and Fatty Acid Content in Fish
3.2. Lipid Amount and Fatty Acid Content in Shellfish
3.3. Lipid Classes in Extracted Oils from Fish and Shellfish
3.4. Fatty Acid Distribution among Lipid Classes
3.5. Nutritional Relevance for the Chilean Population
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Simonyi, A.; Fritsche, K.L.; Chuang, D.Y.; Hannink, M.; Gu, Z.; Greenlief, C.M.; Yao, J.K.; Lee, J.C.; Beversdorf, D.Q. Docosahexaenoic acid (DHA): An essential nutrient and a nutraceutical for brain health and diseases. Prostaglandins Leukot. Essent. Fatty Acids 2018, 136, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Micek, A.; Marventano, S.; Castellano, S.; Mistretta, A.; Pajak, A.; Galvano, F. Dietary n-3 PUFA, fish consumption and depression: A systematic review and meta-analysis of observational studies. J. Affect. Disord. 2016, 205, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Chiu, C.Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 fatty acids and their lipid mediators: Towards an understanding of resolvin and protectin formation. Prostaglandins Other Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef]
- Nakamura, M.T.; Nara, T.Y. Essential fatty acid synthesis and its regulation in mammals. Prostaglandins Leukot. Essent. Fatty Acids 2003, 68, 145–150. [Google Scholar] [CrossRef]
- Martins, D.A.; Custodio, L.; Barreira, L.; Pereira, H.; Ben-Hamadou, R.; Varela, J.; Abu-Salah, K.M. Alternative sources of n-3 long-chain polyunsaturated fatty acids in marine microalgae. Mar. Drugs 2013, 11, 2259–2281. [Google Scholar] [CrossRef] [Green Version]
- Rincón-Cervera, M.Á.; González-Barriga, V.; Valenzuela, R.; López-Arana, S.; Romero, J.; Valenzuela, A. Profile and distribution of fatty acids in edible parts of commonly consumed marine fishes in Chile. Food Chem. 2019, 274, 123–129. [Google Scholar] [CrossRef]
- Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C. Fatty acids from fish: The anti-inflammatory potential of long-chain omega-3 fatty acids. Nutr. Rev. 2010, 68, 280–289. [Google Scholar] [CrossRef]
- Burlingame, B.; Nishida, C.; Uauy, R.; Weisell, R. Fats and fatty acids in human nutrition: Introduction. Ann. Nutr. Metab. 2009, 55, 5–7. [Google Scholar] [CrossRef]
- Cook, C.M.; Hallaraker, H.; Saebo, P.C.; Innis, S.M.; Kelley, K.M.; Sanoshy, K.D.; Berger, A.; Maki, K.C. Bioavailability of long chain omega-3 polyunsaturated fatty acids from phospholipid-rich herring roe oil in men and women with mildly elevated triacylglycerols. Prostaglandins Leukot. Essent. Fatty Acids 2016, 111, 17–24. [Google Scholar] [CrossRef]
- Michalski, M.C.; Genot, C.; Gayet, C.; Lopez, C.; Fine, F.; Joffre, F.; Vendeuvre, J.L.; Bouvier, J.; Chardigny, J.M.; Raynal-Ljutovak, K.; et al. Multiscale structures of lipids in foods as parameters affecting fatty acid bioavailability and lipid metabolism. Prog. Lipid Res. 2013, 52, 354–373. [Google Scholar] [CrossRef] [Green Version]
- Boran, G.; Karaçam, H.; Boran, M. Changes in the quality of fish oils due to storage temperature and time. Food Chem. 2006, 98, 693–698. [Google Scholar] [CrossRef]
- Prato, E.; Biandolino, F. Total lipid content and fatty acid composition of commercially important fish species from the Mediterranean, Mar Grande Sea. Food Chem. 2012, 131, 1233–1239. [Google Scholar] [CrossRef]
- Cladis, D.P.; Kleiner, A.C.; Freiser, H.H.; Santerre, C.R. Fatty acid profiles of commercially available finfish fillets in the United States. Lipids 2014, 49, 1005–1018. [Google Scholar] [CrossRef]
- Zhang, X.; Ning, X.; He, X.; Sun, X.; Yu, X.; Cheng, Y.; Yu, R.Q.; Wu, Y. Fatty acid composition analyses of commercially important fish species from the Pearl River Estuary, China. PLoS ONE 2020, 15, e0228276. [Google Scholar] [CrossRef] [Green Version]
- Huynh, M.D.; Kitts, D.D. Evaluating nutritional quality of pacific fish species from fatty acid signatures. Food Chem. 2009, 114, 912–918. [Google Scholar] [CrossRef]
- Cerna, F.; Plaza, G. Life history parameters of chub mackerel (Scomber japonicus) from two areas off Chile. Bull. Mar. Sci. 2014, 90, 833–848. [Google Scholar] [CrossRef]
- Parada, C.; Gretchina, A.; Vásquez, S.; Belmadani, A.; Combes, V.; Ernst, B.; Di Lorenzo, E.; Porobic, J.; Sepúlveda, A. Expanding the conceptual framework of the spatial population structure and life history of jack mackerel in the eastern South Pacific: An oceanic seamount region as potential spawning/nursery habitat. ICES J. Mar. Sci. 2017, 74, 2398–2414. [Google Scholar] [CrossRef]
- Huynh, M.D.; Kitts, D.D.; Hu, C.; Trites, A.W. Comparison of fatty acid profiles of spawning and non-spawning Pacific herring, Clupea harengus pallasi. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 146, 504–511. [Google Scholar] [CrossRef]
- Soriguer, F.; Serna, S.; Valverde, E.; Hernando, J.; Martín-Reyes, A.; Soriguer, M.; Pareja, A.; Tinahones, F.; Esteva, I. Lipid, protein, and calorie content of different Atlantic and Mediterranean fish, shellfish, and molluscs commonly eaten in the south of Spain. Eur. J. Epidemiol. 1997, 13, 451–463. [Google Scholar] [CrossRef]
- Chouinard-Watkins, R.; Lacombe, R.J.S.; Metherel, A.H.; Masoodi, M.; Bazinet, R.P. DHA Esterified to Phosphatidylserine or Phosphatidylcholine is More Efficient at Targeting the Brain than DHA Esterified to Triacylglycerol. Mol. Nutr. Food Res. 2019, 63, e1801224. [Google Scholar] [CrossRef]
- Halver, J.E.; Hardy, R.W. Fish nutrition. In The Lipids, 3rd ed.; Sargent, J.R., Tocher, D.R., Bell, G., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 182–257. [Google Scholar]
- Devadason, C.; Jayasinghe, C.; Sivakanesan, R.; Senarath, S.; Beppu, F.; Gotoh, N. Comparative analysis of lipid content and fatty acid composition of commercially important fish and shellfish from Sri Lanka and Japan. J. Oleo Sci. 2016, 65, 543–556. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, C.E.; Vasconcelos, M.A.; Ribeiro, M.A.; Sarubbo, L.A.; Andrade, S.A.; Filho, A.B. Nutritional and lipid profiles in marine fish species from Brazil. Food Chem. 2014, 160, 67–71. [Google Scholar] [CrossRef]
- Li, G.; Sinclair, A.J.; Li, D. Comparison of lipid content and fatty acid composition in the edible meat of wild and cultured freshwater and marine fish and shrimps from China. J. Agric. Food Chem. 2011, 59, 1871–1881. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Ganguly, S.; Mahanty, A.; Sankar, T.V.; Anandan, R.; Chakraborty, K.; Paul, B.N.; Sarma, D.; Syama Dayal, J.; Venkateshwarlu, G.; et al. DHA and EPA content and fatty acid profile of 39 food fishes from India. BioMed. Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Ospina-E, J.C.; Sierra-C, A.; Ochoa, O.; Pérez-Álvarez, J.A.; Fernández-López, J. substitution of saturated fat in processed meat products: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 113–122. [Google Scholar] [CrossRef]
- Hosseini, H.; Mahmoudzadeh, M.; Rezaei, M.; Mahmoudzadeh, L.; Khaksar, R.; Khosroshahi, N.K.; Babakhani, A. Effect of different cooking methods on minerals, vitamins and nutritional quality indices of kutum roach (Rutilus frisii kutum). Food Chem. 2014, 148, 86–91. [Google Scholar] [CrossRef]
- Zhu, B.W.; Qin, L.; Zhou, D.Y.; Wu, H.T.; Wu, J.; Yang, J.F.; Li, D.M.; Dong, X.P.; Murata, Y. Extraction of lipid from sea urchin (Strongylocentrotus nudus) gonad by enzyme-assisted aqueous and supercritical carbon dioxide methods. Eur. Food Res. Technol. 2010, 230, 737–743. [Google Scholar] [CrossRef]
- Abd Aziz, N.; Azlan, A.; Ismail, A.; Alinafiah, S.M.; Razman, M.R. Quantitative determination of fatty acids in marine fish and shellfish from warm water of Straits of Malacca for nutritional purposes. BioMed. Res. Int. 2012, 2013, 1–12. [Google Scholar] [CrossRef]
- Mahaffey, K.R. Fish and shellfish as dietary sources of methylmercury and the ω-3 fatty acids, eicosahexaenoic acid and docosahexaenoic acid: Risks and benefits. Environ. Res. 2004, 95, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Archana, A.; Babu, K.R. Nutrient composition and antioxidant activity of gonads of sea urchin Stomopneustes variolaris. Food Chem. 2016, 197, 597–602. [Google Scholar] [CrossRef]
- Martínez-Pita, I.; García, F.J.; Pita, M.L. Males and females gonad fatty acids of the sea urchins Paracentrotus lividus and Arbacia lixula (Echinodermata). Helgol. Mar. Res. 2010, 64, 135–142. [Google Scholar] [CrossRef] [Green Version]
- González, S.J.; Cáceres, C.W.; Ojeda, F.P. Ecología nutricional y alimenticia del erizo comestible Loxechinus albus en el norte de Chile. Rev. Chil. Hist. Nat. 2008, 81, 575–584. [Google Scholar]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; López-Hernández, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Ríos, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Resek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.R.; Scheibling, R.E.; Iverson, S.J.; Gagnon, P. Fatty acid profiles in the gonads of the sea urchin Strongylocentrotus droebachiensis on natural algal diets. Mar. Ecol. Prog. Ser. 2008, 373, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Byun, H.G.; Eom, T.K.; Jung, W.K.; Kim, S.K. Characterization of fish oil extracted from fish processing by-products. Prev. Nutr. Food Sci. 2008, 13, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.Y.; Park, W.K.; Suzuki, H. Analyses of glycolipids from fish, shellfish, and sea snake lipids by high-performance liquid chromatography. J. Agric. Food Chem. 1999, 47, 960–963. [Google Scholar] [CrossRef]
- Da Costa, E.; Silva, J.; Mendonça, S.H.; Abreu, M.H.; Domingues, M.R. Lipidomic approaches towards deciphering glycolipids from microalgae as a reservoir of bioactive lipids. Mar. Drugs 2016, 14, 101. [Google Scholar] [CrossRef] [Green Version]
- Echeverría, F.; Valenzuela, R.; Hernández-Rodas, C.; Valenzuela, A. Docosahexaenoic acid (DHA), a fundamental fatty acid for the brain: New dietary sources. Prostaglandins Leukot. Essent. Fatty Acids 2017, 124, 1–10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name in English | Common Name in Chile | Scientific Name | |
|---|---|---|---|
| Fish | Yellowtail amberjack | Palometa | Seriola lalandi |
| Palm ruff | Cojinoba del Norte | Seriolella violacea | |
| Red cusk-eel | Congrio colorado | Genypterus chilensis | |
| Croaker | Corvina | Cilus gilberti | |
| Mackerel | Caballa | Scomber japonicus | |
| Chilean hake | Merluza común | Merluccius gayi gayi | |
| Jack mackerel | Jurel | Trachurus murphyi | |
| Chilean sandperch | Rollizo | Pinguipes chilensis | |
| Shellfish | Clam | Almeja | Venus antiqua |
| Red squat lobster | Langostino colorado | Pleuroncodes monodon | |
| Yellow squat lobster | Langostino amarillo | Cervimunida johni | |
| Deep water shrimp | Camarón nailon | Heterocarpus reedi | |
| Sea squirt | Piure | Pyura chilensis | |
| Razor clam | Macha | Mesodesma donacium | |
| Chilean abalone | Loco | Concholepas concholepas | |
| Stone crab | Jaiba marmola | Cancer edwardsi | |
| Sea urchin | Erizo | Loxechinus albus |
| Yellowtail Amberjack | Palm Ruff | Red Cusk-eel | Croaker | Mackerel | Chilean Hake | Jack Mackerel | Chilean Sandperch | |
|---|---|---|---|---|---|---|---|---|
| Total lipids (g/100 g fillet) | 4.59 ± 0.30 | 1.71 ± 0.07 | 0.69 ± 0.06 | 1.44 ± 0.05 | 6.44 ± 0.21 | 1.29 ± 0.08 | 3.77 ± 0.22 | 2.72 ± 0.11 |
| FA content (mg/100 g fillet) | ||||||||
| 14:0 | 99.89 ± 3.63 | 52.42 ± 2.88 | 2.23 ± 0.02 | 37.16 ± 1.18 | 262.13 ± 20.52 | 24.96 ± 0.61 | 193.70 ± 6.41 | 93.14 ± 9.00 |
| 15:0 | 25.87 ± 1.00 | 13.90 ± 0.71 | 0.86 ± 0.06 | 5.21 ± 0.25 | 62.67 ± 5.37 | 3.77 ± 0.09 | 20.58 ± 0.64 | 14.49 ± 1.51 |
| 16:0 | 794.74 ± 22.98 | 262.49 ± 11.19 | 55.60 ± 2.99 | 208.88 ± 7.02 | 1248.20 ± 75.94 | 166.52 ± 3.65 | 688.38 ± 17.62 | 593.53 ± 54.78 |
| 17:0 | 29.51 ± 1.07 | 10.39 ± 0.41 | 1.17 ± 0.10 | 4.95 ± 0.25 | 53.24 ± 3.22 | 3.60 ± 0.11 | 13.55 ± 1.25 | 11.48 ± 1.15 |
| 18:0 | 295.55 ± 8.64 | 117.31 ± 4.77 | 25.81 ± 1.43 | 65.85 ± 2.41 | 416.72 ± 22.62 | 41.94 ± 0.57 | 201.07 ± 3.74 | 138.33 ± 11.11 |
| 20:0 | 9.72 ± 0.38 | 5.18 ± 0.18 | 0.36 ± 0.01 | 2.53 ± 0.09 | 19.57 ± 1.35 | 1.77 ± 0.07 | 6.80 ± 0.60 | 6.65 ± 1.08 |
| 24:0 | 4.52 ± 0.21 | 0.00 ± 0.00 | 0.34 ± 0.04 | 1.41 ± 0.11 | 3.00 ± 0.38 | 1.34 ± 0.18 | 2.06 ± 0.27 | 1.94 ± 0.17 |
| Σ SFA | 1259.78 ± 24.87 | 461.69 ± 12.53 | 86.36 ± 3.32 | 326.00 ± 7.53 | 2065.53 ± 82.10 | 243.91 ± 3.76 | 1126.14 ± 19.18 | 859.56 ± 56.56 |
| 16:1n-7 | 165.81 ± 7.83 | 35.97 ± 1.07 | 5.61 ± 0.24 | 61.76 ± 3.29 | 198.98 ± 17.29 | 33.68 ± 0.76 | 169.38 ± 5.68 | 319.91 ± 35.98 |
| 18:1n-9 | 803.83 ± 23.33 | 235.10 ± 10.49 | 32.67 ± 1.66 | 97.50 ± 4.61 | 1255.30 ± 78.65 | 70.94 ± 1.44 | 533.70 ± 15.89 | 382.75 ± 38.68 |
| 18:1n-7 | 102.16 ± 3.07 | 47.09 ± 2.44 | 9.68 ± 0.53 | 34.69 ± 1.46 | 257.38 ± 16.41 | 34.66 ± 1.04 | 115.04 ± 2.57 | 127.39 ± 12.64 |
| 20:1n-9 | 41.84 ± 1.12 | 27.90 ± 1.34 | 1.17 ± 0.03 | 8.86 ± 0.47 | 155.43 ± 9.58 | 5.94 ± 0.43 | 60.77 ± 2.57 | 25.27 ± 2.50 |
| 24:1n-9 | 33.41 ± 0.80 | 12.30 ± 0.50 | 1.78 ± 0.08 | 5.96 ± 0.08 | 30.41 ± 1.49 | 6.58 ± 0.94 | 20.75 ± 0.64 | 8.20 ± 0.91 |
| Σ MUFA | 1147.04± 24.84 | 358.35 ± 10.92 | 50.92 ± 1.76 | 208.77 ± 5.87 | 1897.50 ± 82.75 | 151.79 ± 2.19 | 899.65 ± 17.27 | 863.50 ± 54.39 |
| 18:2n-6 | 55.89 ± 6.52 | 8.87 ± 0.45 | 2.76 ± 0.11 | 15.94 ± 0.99 | 129.64 ± 8.13 | 17.94 ± 0.80 | 71.03 ± 1.90 | 32.11 ± 3.24 |
| 20:4n-6 | 31.98 ± 3.06 | 70.63 ± 2.79 | 10.95 ± 0.99 | 15.52 ± 0.46 | 99.85 ± 4.69 | 11.66 ± 0.28 | 29.26 ± 0.83 | 70.02 ± 4.94 |
| 18:3n-3 | 22.41 ± 4.00 | 4.13 ± 0.16 | 0.36 ± 0.05 | 3.19 ± 0.29 | 42.79 ± 3.41 | 5.52 ± 0.14 | 15.47 ± 0.44 | 5.91 ± 0.56 |
| 18:4n-3 | 35.25 ± 5.87 | 9.02 ± 0.51 | 0.37 ± 0.03 | 10.45 ± 0.67 | 97.78 ± 8.48 | 10.51 ± 0.31 | 48.75 ± 1.77 | 8.59 ± 0.90 |
| 20:4n-3 | 22.24 ± 0.78 | 6.69 ± 0.43 | 0.61 ± 0.08 | 5.82 ± 0.30 | 37.66 ± 3.04 | 3.89 ± 0.10 | 23.97 ± 0.57 | 8.07 ± 0.58 |
| 20:5n-3 (EPA) | 154.41 ± 6.53 | 84.01 ± 5.11 | 40.76 ± 2.01 | 106.78 ± 5.10 | 414.72 ± 30.21 | 121.38 ± 5.92 | 282.01 ± 8.58 | 285.87 ± 22.97 |
| 22:5n-3 | 80.63 ± 3.10 | 37.25 ± 2.16 | 7.61 ± 0.52 | 30.65 ± 1.28 | 128.06 ± 7.67 | 10.92 ± 0.43 | 92.57 ± 1.80 | 58.00 ± 3.96 |
| 22:6n-3 (DHA) | 761.35 ± 18.56 | 220.02 ± 13.20 | 74.39 ± 5.82 | 187.80 ± 7.13 | 955.95 ± 46.90 | 188.00 ± 3.37 | 504.88 ± 7.57 | 221.73 ± 10.66 |
| Σ PUFA | 1164.17 ± 22.36 | 440.63 ± 14.61 | 137.81 ± 6.26 | 376.15 ± 8.96 | 1906.45 ± 57.90 | 369.81 ± 6.89 | 1067.96 ± 11.92 | 690.30 ± 26.33 |
| Σ n-6 PUFA | 87.87 ± 7.20 | 79.50 ± 2.83 | 13.71 ± 0.99 | 31.46 ± 1.09 | 229.49 ± 9.39 | 29.60 ± 0.84 | 100.30 ± 2.07 | 102.13 ± 5.90 |
| Σ n-3 PUFA | 1076.29 ± 21.17 | 361.13 ± 14.33 | 124.10 ± 6.18 | 344.69 ± 8.89 | 1676.95 ± 57.13 | 340.21 ± 6.83 | 967.66 ± 11.74 | 588.17 ± 25.66 |
| Σ EPA + DHA | 915.76 ± 19.68 | 304.04 ± 14.15 | 115.15 ± 6.16 | 294.57 ± 8.76 | 1370.67 ± 55.79 | 309.38 ± 6.81 | 786.90 ± 11.44 | 507.60 ± 25.32 |
| PUFA/SFA | 0.92 | 0.95 | 1.60 | 1.15 | 0.92 | 1.52 | 0.95 | 0.80 |
| n-6/n-3 ratio | 0.08 | 0.22 | 0.11 | 0.09 | 0.14 | 0.09 | 0.10 | 0.17 |
| HH | 2.14 | 2.10 | 2.93 | 1.86 | 2.00 | 2.23 | 1.73 | 1.54 |
| Amount of flesh (g) to supply 250 mg EPA + DHA | 27.3 | 82.2 | 217.1 | 84.9 | 18.2 | 80.8 | 31.8 | 49.3 |
| Clam | Red Squat Lobster | Yellow Squat Lobster | Deep Water Shrimp | Sea Squirt | Razor Clam | Chilean Abalone | Stone Crab | Sea Urchin | |
|---|---|---|---|---|---|---|---|---|---|
| Total lipids (g/100 g meat) | 1.47 ± 0.12 | 1.23 ± 0.08 | 1.32 ± 0.07 | 1.35 ± 0.06 | 2.96 ± 0.25 | 1.94 ± 0.07 | 0.87 ± 0.04 | 1.20 ± 0.16 | 7.23 ± 0.41 |
| FA content (mg/100 g meat) | |||||||||
| 14:0 | 19.70 ± 0.65 | 3.66 ± 0.17 | 2.86 ± 0.16 | 7.75 ± 0.23 | 96.76 ± 7.13 | 20.63 ± 1.23 | 5.94 ± 0.30 | 2.02 ± 0.27 | 1762.43 ± 74.47 |
| 15:0 | 5.60 ± 0.24 | 12.09 ± 0.18 | 2.42 ± 0.02 | 5.58 ± 0.25 | 23.98 ± 3.25 | 3.97 ± 0.34 | 0.29 ± 0.03 | 1.45 ± 0.07 | 22.12 ± 1.83 |
| 16:0 | 130.30 ± 5.89 | 73.52 ± 2.01 | 72.34 ± 1.21 | 93.06 ± 2.17 | 314.73 ± 19.17 | 113.71 ± 7.57 | 43.60 ± 3.69 | 62.19 ± 2.21 | 1022.63 ± 51.13 |
| 17:0 | 50.57 ± 2.06 | 9.69 ± 0.26 | 4.67 ± 0.03 | 5.05 ± 0.26 | 12.66 ± 0.80 | 2.79 ± 0.21 | 25.63 ± 2.17 | 8.68 ± 0.33 | 35.49 ± 1.45 |
| 18:0 | 56.03 ± 3.03 | 28.90 ± 0.74 | 23.50 ± 0.06 | 29.67 ± 0.53 | 77.97 ± 4.03 | 62.99 ± 4.74 | 25.31 ± 1.07 | 39.06 ± 1.92 | 91.87 ± 4.72 |
| 20:0 | 1.11 ± 0.03 | 2.96 ± 0.17 | 4.66 ± 0.17 | 1.85 ± 0.14 | 7.85 ± 0.20 | 1.64 ± 0.08 | 0.59 ± 0.03 | 1.01 ± 0.10 | 27.94 ± 1.54 |
| 24:0 | 4.41 ± 0.31 | 0.79 ± 0.09 | 0.50 ± 0.08 | 1.04 ± 0.11 | 0.00 ± 0.00 | 0.67 ± 0.06 | 0.25 ± 0.02 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Σ SFA | 267.73 ± 6.98 | 131.61 ± 2.18 | 110.96 ± 1.23 | 144.00 ± 2.28 | 533.94 ± 21.22 | 206.40 ± 9.02 | 101.62 ± 4.43 | 114.40 ± 2.96 | 2962.48 ± 90.51 |
| 16:1n-7 | 52.13 ± 2.14 | 4.90 ± 0.14 | 0.79 ± 0.04 | 3.57 ± 0.07 | 152.64 ± 11.13 | 26.16 ± 2.11 | 3.76 ± 0.27 | 21.81 ± 1.07 | 138.93 ± 6.46 |
| 18:1n-9 | 17.47 ± 0.86 | 65.51 ± 2.04 | 62.92 ± 0.96 | 82.66 ± 1.22 | 81.74 ± 3.02 | 24.41 ± 1.24 | 8.69 ± 0.50 | 66.37 ± 2.37 | 71.71 ± 2.79 |
| 18:1n-7 | 24.35 ± 1.32 | 20.43 ± 0.66 | 21.60 ± 0.34 | 33.46 ± 0.98 | 147.60 ± 9.35 | 19.28 ± 1.53 | 5.56 ± 0.58 | 24.91 ± 1.10 | 571.99 ± 30.34 |
| 20:1n-9 | 11.75 ± 0.57 | 2.20 ± 0.22 | 1.48 ± 0.07 | 0.50 ± 0.04 | 9.40 ± 0.62 | 14.49 ± 0.97 | 7.46 ± 0.34 | 1.62 ± 0.06 | 444.43 ± 33.93 |
| 24:1n-9 | 0.00 ± 0.00 | 0.86 ± 0.14 | 0.44 ± 0.02 | 1.77 ± 0.16 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.89 ± 0.30 |
| Σ MUFA | 105.70 ± 2.72 | 93.91 ± 2.17 | 87.23 ± 1.02 | 121.96 ± 1.57 | 391.38 ± 15.01 | 84.35 ± 3.05 | 25.47 ± 0.88 | 114.71 ± 2.83 | 1229.95 ± 46.05 |
| 18:2n-6 | 2.57 ± 0.06 | 5.02 ± 0.20 | 4.55 ± 0.26 | 5.42 ± 0.07 | 51.11 ± 2.23 | 1.66 ± 0.03 | 7.31 ± 0.93 | 7.60 ± 0.41 | 10.60 ± 0.39 |
| 20:4n-6 | 19.82 ± 0.66 | 20.19 ± 0.54 | 24.93 ± 0.23 | 13.33 ± 0.38 | 18.25 ± 1.50 | 10.03 ± 0.71 | 6.50 ± 0.30 | 15.12 ± 0.93 | 269.72 ± 13.32 |
| 18:3n-3 | 5.81 ± 0.80 | 0.17 ± 0.04 | 1.42 ± 0.05 | 1.93 ± 0.07 | 25.60 ± 1.80 | 1.71 ± 0.20 | 0.24 ± 0.01 | 0.76 ± 0.04 | 5.15 ± 0.57 |
| 18:4n-3 | 10.32 ± 0.42 | 1.97 ± 0.06 | 1.64 ± 0.04 | 0.91 ± 0.09 | 71.44 ± 5.43 | 7.25 ± 0.57 | 1.60 ± 0.15 | 0.66 ± 0.03 | 69.15 ± 2.88 |
| 20:4n-3 | 6.08 ± 0.41 | 0.85 ± 0.15 | 0.88 ± 0.07 | 0.94 ± 0.09 | 0.00 ± 0.00 | 4.54 ± 0.40 | 0.48 ± 0.01 | 1.23 ± 0.03 | 27.08 ± 1.34 |
| 20:5n-3 (EPA) | 130.44 ± 5.74 | 106.31 ± 3.00 | 90.58 ± 1.23 | 119.53 ± 3.43 | 356.97 ± 24.71 | 102.12 ± 7.55 | 46.70 ± 0.12 | 140.85 ± 5.80 | 199.01 ± 10.19 |
| 22:5n-3 | 25.13 ± 1.33 | 3.55 ± 0.14 | 4.88 ± 0.24 | 2.77 ± 0.08 | 12.29 ± 0.87 | 33.86 ± 1.55 | 38.31 ± 0.41 | 9.70 ± 0.56 | 8.40 ± 0.53 |
| 22:6n-3 (DHA) | 83.91 ± 4.87 | 83.53 ± 2.23 | 72.33 ± 2.54 | 67.45 ± 1.80 | 165.71 ± 13.21 | 114.84 ± 6.18 | 16.91 ± 0.40 | 64.77 ± 2.16 | 9.54 ± 1.40 |
| Σ PUFA | 283.27 ± 7.73 | 221.59 ± 3.79 | 201.22 ± 2.86 | 212.28 ± 3.90 | 701.37 ± 28.74 | 276.01 ± 9.93 | 118.06 ± 1.15 | 240.69 ± 6.30 | 598.66 ± 17.15 |
| Σ n-6 PUFA | 22.39 ± 0.66 | 25.21 ± 0.58 | 29.48 ± 0.35 | 18.75 ± 0.39 | 69.36 ± 2.69 | 11.69 ± 0.72 | 13.81 ± 0.97 | 22.72 ± 1.02 | 280.32 ± 13.33 |
| Σ n-3 PUFA | 260.89 ± 7.70 | 196.38 ± 3.74 | 171.73 ± 2.84 | 193.53 ± 3.88 | 632.00 ± 28.61 | 264.32 ± 9.91 | 104.25 ± 0.61 | 217.97 ± 6.22 | 318.34 ± 10.79 |
| Σ EPA + DHA | 214.34 ± 7.52 | 189.83 ± 3.74 | 162.90 ± 2.83 | 186.98 ± 3.88 | 522.68 ± 28.02 | 216.96 ± 9.76 | 63.61 ± 0.42 | 205.62 ± 6.19 | 208.55 ± 10.28 |
| PUFA/SFA | 1.06 | 1.68 | 1.81 | 1.47 | 1.31 | 1.34 | 1.16 | 2.10 | 0.20 |
| n-6/n-3 ratio | 0.09 | 0.13 | 0.17 | 0.10 | 0.11 | 0.04 | 0.13 | 0.10 | 0.88 |
| HH | 1.90 | 3.68 | 3.48 | 2.91 | 1.73 | 2.15 | 2.52 | 4.75 | 0.21 |
| Amount of meat (g) to supply 250 mg EPA + DHA | 116.6 | 131.7 | 153.5 | 133.7 | 47.8 | 115.2 | 393.0 | 121.6 | 119.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rincón-Cervera, M.Á.; González-Barriga, V.; Romero, J.; Rojas, R.; López-Arana, S. Quantification and Distribution of Omega-3 Fatty Acids in South Pacific Fish and Shellfish Species. Foods 2020, 9, 233. https://doi.org/10.3390/foods9020233
Rincón-Cervera MÁ, González-Barriga V, Romero J, Rojas R, López-Arana S. Quantification and Distribution of Omega-3 Fatty Acids in South Pacific Fish and Shellfish Species. Foods. 2020; 9(2):233. https://doi.org/10.3390/foods9020233
Chicago/Turabian StyleRincón-Cervera, Miguel Ángel, Valeria González-Barriga, Jaime Romero, Rodrigo Rojas, and Sandra López-Arana. 2020. "Quantification and Distribution of Omega-3 Fatty Acids in South Pacific Fish and Shellfish Species" Foods 9, no. 2: 233. https://doi.org/10.3390/foods9020233