Discrimination between Fresh and Frozen-Thawed Fish Involved in Food Safety and Fraud Protection

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
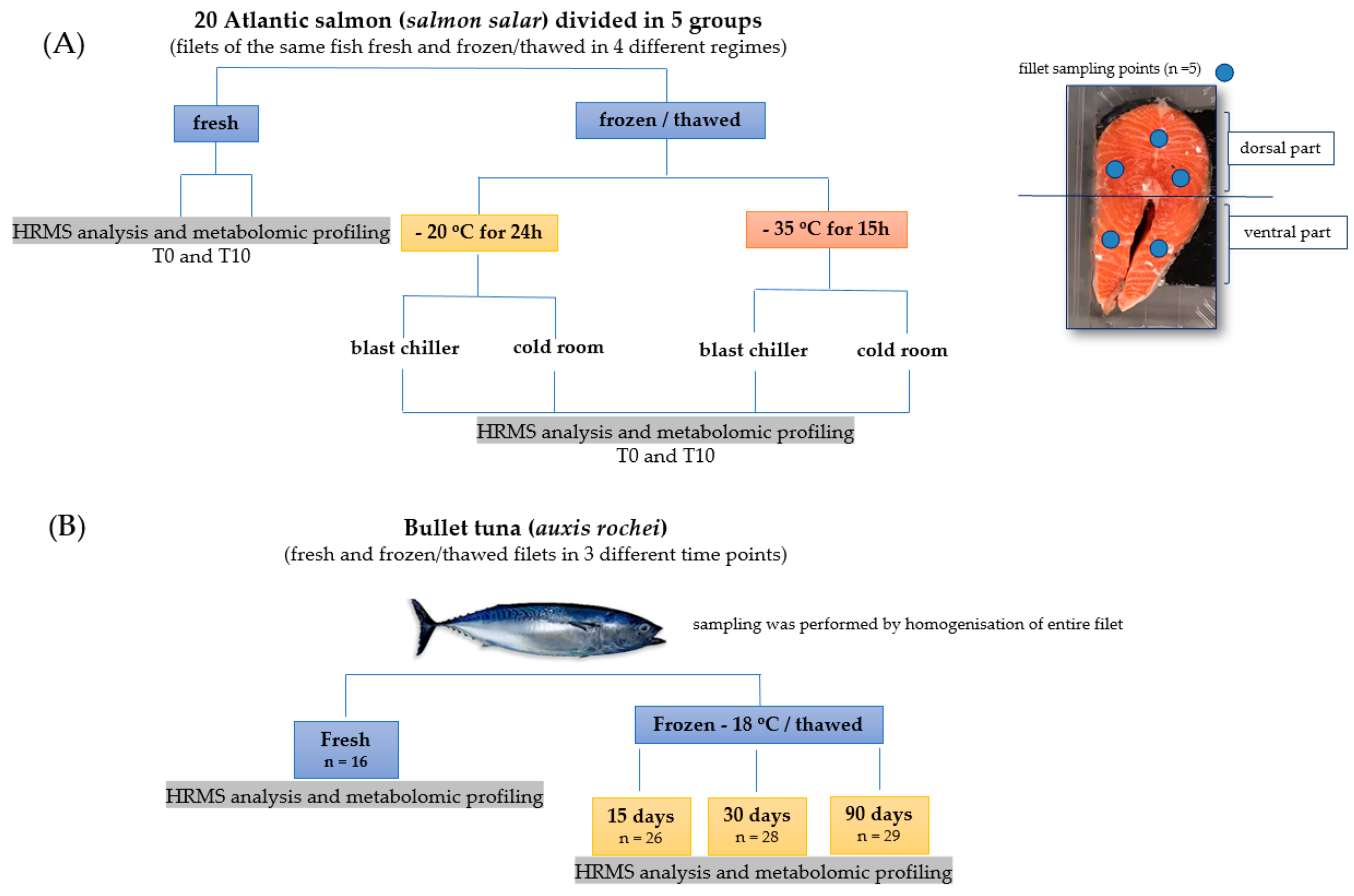
2.2. Experimental Design
2.2.1. Atlantic salmon (Salmo salar) Rapid Freezing/Thawing Processing
2.2.2. Bullet Tuna (Auxis rochei) Long Term Freezing
2.3. Sample Preparation
2.4. HPLC Q-Exactive Orbitrap HRMS Analysis
2.5. Untargeted Metabolomics with Compound Discoverer™ Workflow
3. Results and Discussion
3.1. HRMS Untargeted Metabolomics with Compound Discoverer™ Workflow
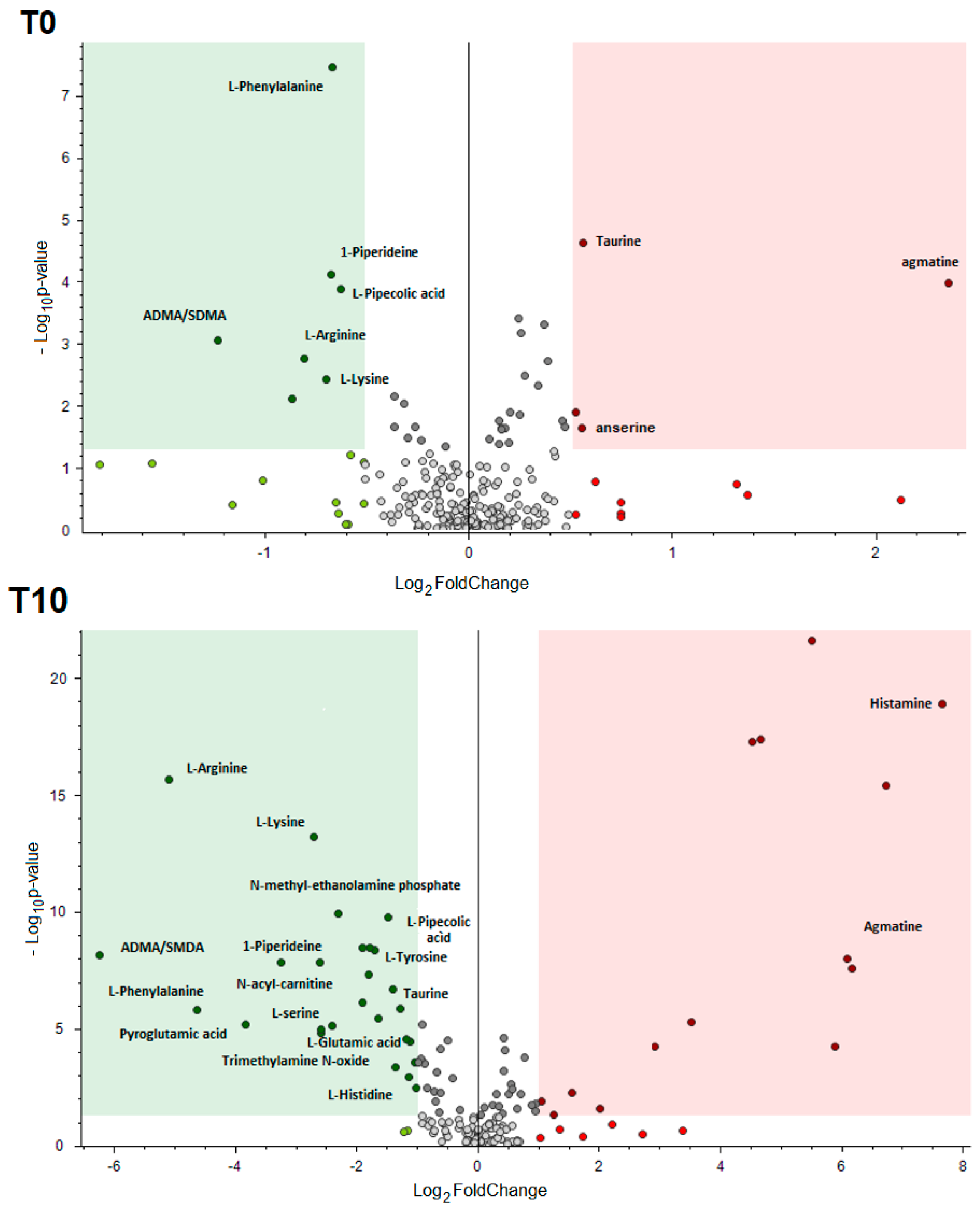
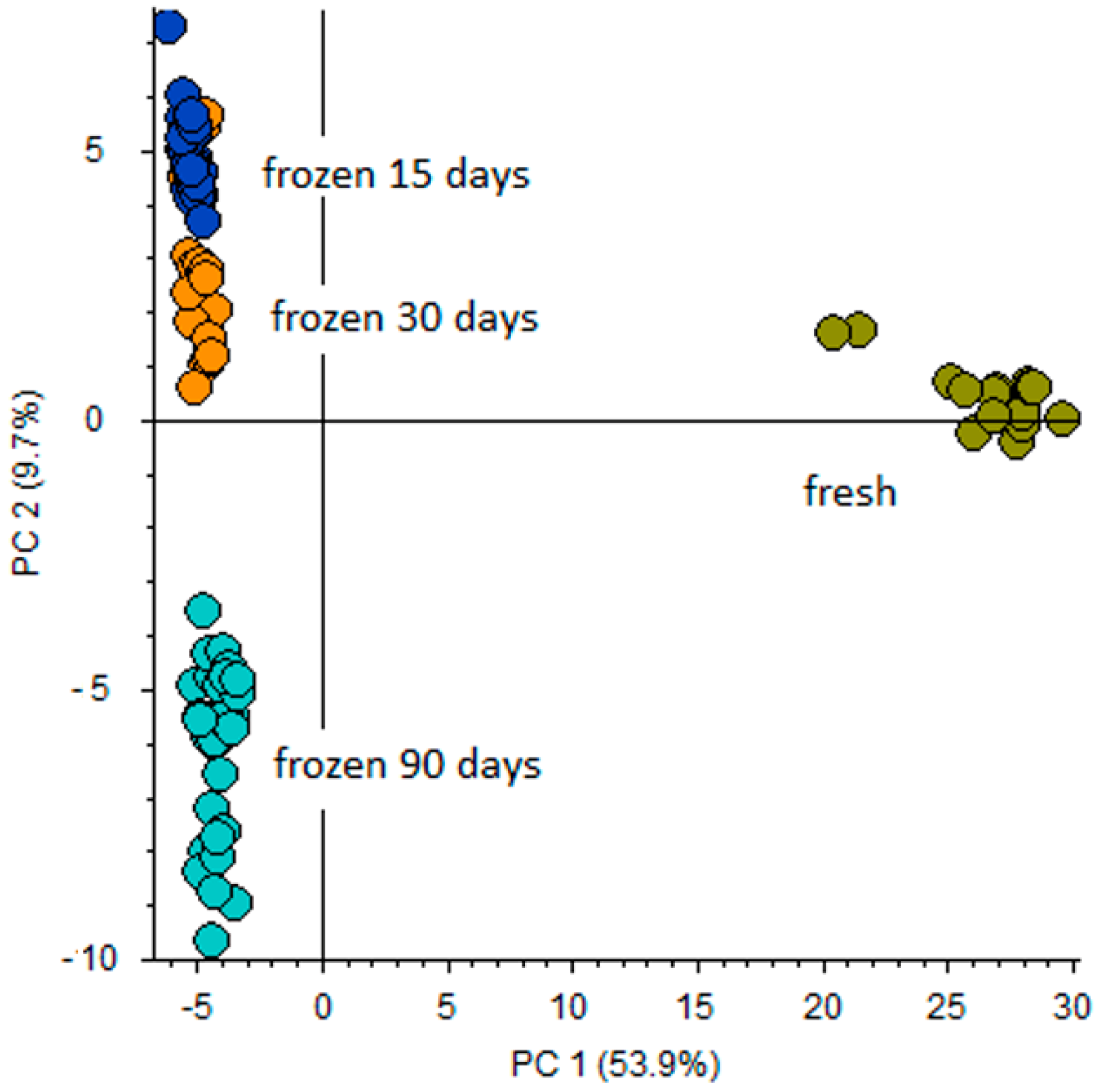
- Full scan (FS) with resolving power of 140,000 identified signals of metabolites; 156 and 148 candidates for Atlantic salmon and bullet tuna, respectively. Applying the Compound Discoverer workflow described above, differential analysis that involved principal component analysis (PCA) and volcano plot (VP) was performed. PCA was used in order to visualize and provide overall differentiation between fresh and frozen/thawed samples. In order to obtain more uniform signals, data were normalized by conversion of the absolute peak areas for each metabolic feature into relative peak intensities. This data was applied to PCA and VP evaluation in order to visualize maximum variation. As preliminary statistical assessment did not give any differences between the freezing regimens used for Atlantic salmon and for bullet tuna, respectively, both PCA and VP were performed comparing the fresh samples to the group of frozen ones, regardless of the freezing/thawing treatment applied. PCA (Figure S2) clearly distinguished the fresh and frozen/thawed sample at T10 for salmon samples, while at T0 there were no differences. Furthermore, VP (Figure 2) analysis gave more precise responses regarding the main differentiators. For the bullet tuna, the PCA score plot clearly distinguished the fresh filets from frozen ones, with notable differences between 90-day samples and the two shorter sampling periods (Figure 3).
- FS-data dependent (FS-DDA) processing was performed on the selected inclusion list of 156 and 148 signals extracted from the FS data collection. This acquisition mode allowed putative identification of the main differentiators and was based on predicted composition according to accurate mass, adduct formation, isotopes ratios and fragmentation patterns. Selection of the best-fit candidates for the signals was performed by software-linked mass fragmentation database libraries (mzCloud and ChemSpider). In some cases, as no satisfactory confirmation was found from existing databases (including external libraries like Human Metabolome Database) presumption of the structural formula was achieved in-house by identification of fragment structure.
3.2. Salmon Metabolite Profile upon Rapid Freezing/Thawing Processing
3.3. Bullet Tuna Metabolites Profile upon Long Freezing/Thawing Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dawson, P.; Al-Jeddawi, W.; Remington, N. Effect of freezing on the shelf life of salmon. Int. J. Food Sci. 2018, 12, 1686121. [Google Scholar] [CrossRef] [Green Version]
- Claret, A.; Guerrero, L.; Aguirre, E.; Rincón, L.; Hernández, M.D.; Martínez, I.; Peleteiro, J.B.; Grau, A.; Rodríguez-Rodríguez, C. Consumer preferences for sea fish using conjoint analysis: Exploratory study of the importance of country of origin, obtaining method, storage conditions and purchasing price. Food Qual. Prefer. 2012, 26, 259–266. [Google Scholar] [CrossRef]
- Pezzolato, M.; Baioni, E.; Maurella, C.; Varello, K.; Meistro, S.; Balsano, A.; Bozzetta, E. Distinguishing between Fresh and Frozen-Thawed Smoked Salmon: Histology to Detect Food Adulteration in High-Value Products. J. Food Prot. 2020, 83, 52–53. [Google Scholar] [CrossRef]
- FAO. GLOBEFISH—Information and Analysis on World Fish Trade. Available online: http://www.fao.org/in-action/globefish/market-reports/resource-detail/en/c/1268636/ (accessed on 16 November 2020).
- Mowi. Salmon Farming Industry Handbook. Available online: https://mowi.com/it/wp-content/uploads/sites/16/2020/06/Mowi-Salmon-Farming-Industry-Handbook-2020.pdf (accessed on 16 November 2020).
- FAO. Fish Finder. Available online: http://www.fao.org/fishery/species/2491/en (accessed on 16 November 2020).
- Gomes, T.L.; Quiazon, K.M.A.; Kotake, M.; Itoh, N.; Yoshinaga, T. Anisakis spp. in fishery products from Japanese waters: Updated insights on host prevalence and human infection risk factors. Parasitol. Int. 2020, 78, 102137. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation (EU) No 1276/2011 of 8 December 2011 amending Annex III to Regulation (EC) No 853/2004 of the European Parliament and of the Council as regards the treatment to kill viable parasites in fishery products for human consumption. Off. J. Eur. Union 2011, 50, 39–41. [Google Scholar]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on risk assessment of parasites in fishery products. EFSA J. 2010, 8, 1543. [Google Scholar] [CrossRef]
- European Parliament. Regulation No 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of food information to consumers. Off. J. Eur. Union 2011, 50, 18–63. [Google Scholar]
- European Parliament. Regulation (EU) No 1379/2013 of the European Parliament and of the Council of 11 December 2013 on the common organisation of the markets in fishery and aquaculture products, amending Council Regulations (EC) No 1184/2006 and (EC) No 1224. Off. J. Eur. Union 2013, 354, 1–21. [Google Scholar]
- Bozzetta, E.; Pezzolato, M.; Cencetti, E.; Varello, K.; ABRAMO, F.; Mutinelli, F.; Ingravalle, F.; Teneggi, E. Histology as a valid and reliable tool to differentiate fresh from frozen-thawed fish. J. Food Prot. 2012, 75, 1536–1541. [Google Scholar] [CrossRef] [PubMed]
- Meistro, S.; Pezzolato, M.; Muscolino, D.; Giarratana, F.; Baioni, E.; Panebianco, A.; Bozzetta, E. Histology as a valid tool to differentiate fresh from frozen-thawed marinated fish. J. Food Prot. 2016, 79, 1457–1459. [Google Scholar] [CrossRef]
- Tinacci, L.; Armani, A.; Guidi, A.; Nucera, D.; Shvartzman, D.; Miragliotta, V.; Coli, A.; Giannessi, E.; Stornelli, M.R.; Fronte, B. Histological discrimination of fresh and frozen/thawed fish meat: European hake (Merluccius merluccius) as a possible model for white meat fish species. Food Control 2018, 92, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Orlova, D.; Kalyuzhnaya, T.; Tokarev, A.; Kuznetsov, Y. New method for veterinary and sanitary control of defrosted meat and fish. Int. J. Vet. Sci. 2020, 9, 317–319. [Google Scholar]
- Watanabe, M.; Suzuki, T.; Ichimaida, K.; Hattori, T.; Ueda, R. Do consumers actually sense that sashimi made from frozen-thawed fish tastes worse than non-frozen one? Int. J. Refrig. 2020, 111, 94–102. [Google Scholar] [CrossRef]
- Kimiya, T.; Sivertsen, A.H.; Heia, K. VIS/NIR spectroscopy for non-destructive freshness assessment of Atlantic salmon (Salmo salar L.) fillets. J. Food Eng. 2013, 116, 758–764. [Google Scholar] [CrossRef]
- Reis, M.M.; Martínez, E.; Saitua, E.; Rodríguez, R.; Perez, I.; Olabarrieta, I. Non-invasive differentiation between fresh and frozen/thawed tuna fillets using near infrared spectroscopy (Vis-NIRS). LWT 2017, 78, 129–137. [Google Scholar] [CrossRef]
- Leduc, F.; Krzewinski, F.; Le Fur, B.; N’Guessan, A.; Malle, P.; Kol, O.; Duflos, G. Differentiation of fresh and frozen/thawed fish, European sea bass (Dicentrarchus labrax), gilthead seabream (Sparus aurata), cod (Gadus morhua) and salmon (Salmo salar), using volatile compounds by SPME/GC/MS. J. Sci. Food Agric. 2012, 92, 2560–2568. [Google Scholar] [CrossRef]
- Iglesias, J.; Medina, I.; Bianchi, F.; Careri, M.; Mangia, A.; Musci, M. Study of the volatile compounds useful for the characterisation of fresh and frozen-thawed cultured gilthead sea bream fish by solid-phase microextraction gas chromatography–mass spectrometry. Food Chem. 2009, 115, 1473–1478. [Google Scholar] [CrossRef]
- Shumilina, E.; Møller, I.A.; Dikiy, A. Differentiation of fresh and thawed Atlantic salmon using NMR metabolomics. Food Chem. 2020, 314, 126227. [Google Scholar] [CrossRef]
- Duflos, G.; Le Fur, B.; Mulak, V.; Becel, P.; Malle, P. Comparison of methods of differentiating between fresh and frozen–thawed fish or fillets. J. Sci. Food Agric. 2002, 82, 1341–1345. [Google Scholar] [CrossRef]
- Popelka, P.; Nagy, J.; Pipová, M.; Marcinčák, S.; Lenhardt, Ľ. Comparison of chemical, microbiological and histological changes in fresh, frozen and double frozen rainbow trout (Oncorhynchus mykiss). Acta Vet. Brno 2014, 83, 157–161. [Google Scholar] [CrossRef]
- Marlard, S.; Doyen, P.; Grard, T. Rapid Multiparameters Approach to Differentiate Fresh Skinless Sea Bass (Dicentrarchus labrax) Fillets from Frozen-Thawed Ones. J. Aquat. Food Prod. Technol. 2019, 28, 253–262. [Google Scholar] [CrossRef]
- Karoui, R.; Thomas, E.; Dufour, E. Utilisation of a rapid technique based on front-face fluorescence spectroscopy for differentiating between fresh and frozen–thawed fish fillets. Food Res. Int. 2006, 39, 349–355. [Google Scholar] [CrossRef]
- Cheng, J.-H.; Sun, D.-W.; Zeng, X.-A.; Liu, D. Recent advances in methods and techniques for freshness quality determination and evaluation of fish and fish fillets: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1012–1225. [Google Scholar] [CrossRef] [PubMed]
- Callao, M.P.; Ruisánchez, I. An overview of multivariate qualitative methods for food fraud detection. Food Control 2018, 86, 283–293. [Google Scholar] [CrossRef]
- Shumilina, E.; Dykyy, A.; Dikiy, A. Development of a statistical model to detect quality and storage conditions of Atlantic salmon. Food Chem. 2018, 258, 381–386. [Google Scholar] [CrossRef]
- Jääskeläinen, E.; Jakobsen, L.M.A.; Hultman, J.; Eggers, N.; Bertram, H.C.; Björkroth, J. Metabolomics and bacterial diversity of packaged yellowfin tuna (Thunnus albacares) and salmon (Salmo salar) show fish species-specific spoilage development during chilled storage. Int. J. Food Microbiol. 2019, 293, 44–52. [Google Scholar] [CrossRef]
- Kaufmann, A.; Maden, K. Easy and fast method for the determination of biogenic amines in fish and fish products with liquid chromatography coupled to orbitrap tandem mass spectrometry. J. AOAC Int. 2018, 101, 336–341. [Google Scholar] [CrossRef]
- Chiesa, L.; Panseri, S.; Pasquale, E.; Malandra, R.; Pavlovic, R.; Arioli, F. Validated multiclass targeted determination of antibiotics in fish with high performance liquid chromatography–benchtop quadrupole orbitrap hybrid mass spectrometry. Food Chem. 2018, 258, 222–230. [Google Scholar] [CrossRef]
- Chiesa, L.M.; Nobile, M.; Malandra, R.; Panseri, S.; Arioli, F. Occurrence of antibiotics in mussels and clams from various FAO areas. Food Chem. 2018, 240, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Zhuang, H.; Zhou, G.; Zhang, J. Investigation of inhibition of lipid oxidation by L-carnosine using an oxidized-myoglobin-mediated washed fish muscle system. LWT 2018, 97, 703–710. [Google Scholar] [CrossRef]
- Aru, V.; Pisano, M.B.; Savorani, F.; Engelsen, S.B.; Cosentino, S.; Marincola, F.C. Metabolomics analysis of shucked mussels’ freshness. Food Chem. 2016, 205, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Liu, Y.; Yu, Q. Metabolomic approach to measuring quality of chilled chicken meat during storage. Poult. Sci. 2020, 99, 2543–2554. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Cao, M.; Yang, Y.; Nagahama, Y.; Zhao, H. Expression pattern of prmt5 in adult fish and embryos of medaka, Oryzias latipes. Fish Physiol. Biochem. 2009, 35, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, L.; Panseri, S.; Pavlovic, R.; Arioli, F. Biogenic amines evaluation in wild Bluefin tuna (Thunnus thynnus) originating from various FAO areas. J. Consum. Prot. Food Saf. 2018, 13, 375–382. [Google Scholar] [CrossRef]
- Ohshima, T.; Wada, S.; Koizumi, C. Accumulation of lyso-form phospholipids in several species of fish flesh during storage at-5 C. Nippon Suisan Gakkaishi 1985, 51, 965–971. [Google Scholar] [CrossRef]
- Chiesa, L.; Panseri, S.; Bonacci, S.; Procopio, A.; Zecconi, A.; Arioli, F.; Cuevas, F.J.; Moreno-Rojas, J.M. Authentication of Italian PDO lard using NIR spectroscopy, volatile profile and fatty acid composition combined with chemometrics. Food Chem. 2016, 12, 296–304. [Google Scholar] [CrossRef]
- Zheng, Z.; Lin, S.; Xue, J.; Shen, Q.; Feng, J.; Jin, R.; Dai, Z. The characterization of myoglobin and myoglobin-induced lipid oxidation in frigate mackerel. J. Food Process. Preserv. 2016, 40, 1438–1447. [Google Scholar] [CrossRef]
- Chaijan, M.; Klomklao, S.; Benjakul, S. Characterisation of muscles from Frigate mackerel (Auxis thazard) and catfish (Clarias macrocephalus). Food Chem. 2013, 139, 414–419. [Google Scholar] [CrossRef]
- Thébault, M.T.; Kervarec, N.; Pichon, R.; Nonnotte, G.; Le Gal, Y. Distribution and potential role of cytosolic water-soluble phosphodiesters in fish. Physiol. Biochem. Zool. 2003, 76, 553–565. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Reference | Fish Species | Method Applied for Differentiation | Identification Criteria/Markers Suggested |
|---|---|---|---|
| Histological methodology | |||
| Bozzetta et al., 2012 [12] | Gilthead, red mullet, swordfish | Specific microscopic changes related to freezing transverse muscle sections with hematoxylin and eosin staining |
|
| Meistro et al., 2016 [13] | Marinated anchovy | ||
| Pezzolato et al., 2020 [3] | Smoked salmon | ||
| Tinacci et al., 2018 [14] | European hake | Standard histological morphological and morphometric parameters |
|
| Orlova et al., 2020 [15] | Not specified | Hematoxylin-eosin staining and microscopy | presence of fibrous areas, tearing of muscle fibers |
| Organoleptic evaluation | |||
| Watanabe et al., 2020 [16] | Sashimi from pacific saury | Consumer panel sensorial evaluation | taste and flavor, fattiness, smooth mouth feel, retro-nasal aroma, redness of meat, overall appearance, texture, springy palatability, wateriness |
| Spectroscopy-based approach | |||
| Kimiya et al., 2013 [17] | Salmon | Visible-Near InfraRed Spectroscopy (Vis-NIRS) | spectral changes due to oxidation of heme proteins |
| Reis et al., 2017 [18] | Tuna | chemometrics based on structural alternations of myoglobin and its oxidative products | |
| Solid phase microextraction gas chromatography/mass spectrometry (SPME-GC/MS) | |||
| Leduc et al., 2012 [19] | European sea bass, gilthead seabream, cod and salmon | SPME-GC/MS analysis of volatile compounds; statistical processing by principal component analysis and ascending hierarchical classification | dimethyl sulfide, 3-methylbutanal, ethyl acetate and 2-methylbutanal |
| Iglesias et al., 2009 [20] | Gilthead sea bream | SPME-GC/MS analysis of volatile compounds; peroxide value; thiobarbituric acid-reactive substances (TBARS) index | 1-octen-3-ol, 1-penten-3-ol, and Z-4-heptenal |
| Metabolomic nuclear magnetic resonance spectroscopy (NMR) approach | |||
| Shumilina et al., 2020 [21] | Salmon | NMR characterization; chemometrics statistical evaluation | Measurement of aspartate in frozen/thawed samples |
| Metabolomic high resolution mass spectrometry (HRMS) approach | |||
| This study | Salmon, bullet tuna | HRMS characterization; multivariate data analysis |
|
| Combination of different analytical strategies | |||
| Duflos et al., 2002 [22] | Plaice, whiting, mackerel | Physical determination (torrymeter); physiological examination (eye lens); enzymatic assays: (α-glucosidase, β-N-acetylglucosaminidase and β-hydroxyacyl-CoA-dehydrogenase) |
|
| Popelka et al., 2014 [23] | Rainbow trout | Chemical composition (water, protein and fat content); expressible drip; total volatile nitrogen (TVN); microbiological analyses; histological examinations |
|
| Marlard et al., 2019 [24] | Sea bass exudate | Proteins amount; α-d-glucosidase specific activity; nucleotides and related compounds (NRCs); free Ca2+ | NRs and Ca2+ concentration |
| Compound | Fresh | Frozen/Thawed | p Values 2, Regulation 3 | |||
|---|---|---|---|---|---|---|
| −20 °C | −35 °C | |||||
| Blast Chiller | Cold Room | Blast Chiller | Cold Room | |||
| T0–hawed samples analyzed immediately | ||||||
| Anserine (β-alanyl-3-metil l-histidine) | 2214.1 ± 228.8 | 1197.3 ± 422.9 | 1312.3 ± 553.3 | 1521.2 ± 541.3 | 1638.2 ± 221.3 | 0.025, ↓ |
| Arginine | 24.3 ± 9.8 | 38.3 ± 6.4 | 39.7 ± 16.4 | 32.3 ± 7.9 | 35.8 ± 17.1 | <0.001, ↑ |
| Agmatine | 12.0 ± 20.8 | 0.3 ± 0.5 | 0.4 ± 0.7 | 0.3 ± 0.3 | 0.1 ± 0.2 | <0.001, ↓ |
| Adma/Sdma (N, N-dimethyl-arginine) | 4.3 ± 2.8 | 8.3 ± 2.1 | 7.3 ± 2.9 | 6.8 ± 1.8 | 10.0 ± 3.9 | 0.028, ↑ |
| Lysine | 35.5 ± 5.2 | 57.1 ± 23.8 | 52.3 ± 17.9 | 51.6 ± 13.4 | 63.2 ± 12.3 | 0.017, ↑ |
| Pipecolic acid | 1.7 ± 0.7 | 3.7 ± 2.7 | 5.2 ± 3.8 | 13.8 ± 8.1 | 25.8 ± 1.1 | 0.011, ↑ |
| 1-Piperideine | 5.5 ± 2.1 | 9.6 ± 4.3 | 10.2 ± 3.4 | 10.3 ± 2.71 | 12.2 ± 1.9 | <0.001, ↑ |
| Phenylalanine | 15.7 ± 7.3 | 25.9 ± 4.2 | 25.6 ± 4.0 | 22.9 ± 1.5 | 28.2 ± 8.4 | 0.008, ↑ |
| Taurine | 47.7 ± 7.1 | 25.9 ± 4.2 | 38.6 ± 4.0 | 24.9 ± 2.5 | 38.2 ± 0.4 | 0.006, ↓ |
| T10–thawed samples analyzed after 10 days refrigerator (4 °C) storage | ||||||
| Histidine | 94.1 ± 41.1 | 124.7 ± 37.2 | 124.2 ± 43.2 | 130.6 ± 32.6 | 158.6 ± 64.6 | 0.005, ↑ |
| Histamine | 38.2 ± 13.3 | 1.6 ± 1.55 | 2.8 ± 1.8 | 1.9 ± 2.9 | 2.5 ± 2.6 | <0.001, ↓ |
| Arginine | 11.4 ± 12.3 | 78.5 ± 9.2 | 113.47 ± 70.3 | 56.3 ± 17.9 | 48.8 ± 31.1 | <0.001, ↑ |
| Agmatine | 124.8 ± 82.5 | 11.6 ± 16.1 | 13.1 ± 10.2 | 19.2 ± 0.3 | 20.2 ± 2.2 | <0.001, ↓ |
| Adma/Sdma (N, N-dimethyl-arginine) | 5.2 ± 3.1 | 11.1 ± 2.0 | 19.6 ± 2.9 | 19.8 ± 7.7 | 20.0 ± 13.9 | 0.021, ↑ |
| N-acyl-carnitine | 34.8 ± 12.5 | 66.4 ± 13.2 | 55.1 ± 27.2 | 53.3 ± 11.2 | 62.2 ± 21.9 | 0.043, ↑ |
| Lysine | 22.8 ± 12.1 | 59.1 ± 26.8 | 93.3 ± 53.9 | 62.0 ± 26.4 | 97.4 ± 24.3 | 0.036, ↑ |
| Cadaverine | 27.8 ± 9.1 | n.d.4 | n.d. | n.d. | n.d. | <0.001, ↓ |
| d-Pipecolic acid | 12.7 ± 12.7 | 33.4 ± 2.7 | 35.2 ± 14.8 | 23.8 ± 11.1 | 45.8 ± 11.1 | <0.001, ↑ |
| 1-Piperideine | 5.7 ± 2.75 | 16.5 ± 4.70 | 19.45 ± 9.97 | 13.33 ± 5.86 | 20.5 ± 11.4 | <0.001, ↑ |
| Serine | 1.6 ± 1.3 | 7.2 ± 1.1 | 7.5 ± 3.7 | 4.9 ± 2.3 | 7.4 ± 1.0 | <0.001, ↑ |
| Glutamic acid | 11.0 ± 7.1 | 19.2 ± 2.5 | 22.0 ± 12.5 | 18.2 ± 7.4 | 22.9 ± 15.6 | 0.039, ↑ |
| Pyroglutamic acid | 4.5 + 1.5 | 9.9 ± 2.1 | 10.8 ± 4.2 | 8.2 ± 3.1 | 11.8 ± 5.2 | 0.006, ↑ |
| Phenylalanine | 3.6 ± 1.5 | 10.7 ± 1.8 | 12.1 ± 8.6 | 10.0 ± 3.6 | 18.2 ± 12.4 | 0.005, ↑ |
| Tyrosine | 23.6 ± 12.8 | 49.9 ± 7.2 | 57.6 ± 19.0 | 36.9 ± 13.5 | 53.2 ± 32.4 | <0.001,↑ |
| Taurine | 8.1 ± 2.1 | 15.9 ± 4.1 | 13.6 ± 24.0 | 14.8 ± 1.5 | 18.2 ± 1.4 | <0.001, ↑ |
| N-methyl-ethanolamine phosphate | 23.8 ± 12.9 | 32.67 ± 5.8 | 54.1 ± 2.5 | 39.82 ± 18.2 | 47.2 ± 12.8 | <0.001, ↑ |
| Trimethylamine N-oxide | 264.6 ± 26.25 | 627.2 ± 123.2 | 590.0 ± 37.12 | 438.1 ± 123.2 | 541.0 ± 54.1 | <0.001, ↑ |
| Compound | Fresh | Frozen/Thawed | p Values 2, Regulation 3 | ||
|---|---|---|---|---|---|
| 15 Days | 30 Days | 90 Days | |||
| Arginine | 22.4 ± 12.3 | 58.5 ± 19.2 | 43.7 ± 20.1 | 66.3 ± 7.9 | 0.0038, ↑ |
| γ-glutamyl-S-methylcysteinyl-β-alanine | 109.2 ± 12.8 | 12.4 ± 4.7 | 10.7 ± 5.3 | 7.1 ± 4.7 | <0.001, ↓ |
| N-methyl-ethanolamine-phosphate | 42.8 ± 22.5 | 100.6 ± 10.2 | 205.1 ± 98.5 | 350.8 ± 10.2 | <0.001, ↑ |
| l-α-Glyceryl-phosphoryl-choline | 7.8 ± 7.6 | 52.2 ± 12.2 | 42.1 ± 12.8 | 20.7 ± 14.9 | <0.001, ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiesa, L.M.; Pavlovic, R.; Nobile, M.; Di Cesare, F.; Malandra, R.; Pessina, D.; Panseri, S. Discrimination between Fresh and Frozen-Thawed Fish Involved in Food Safety and Fraud Protection. Foods 2020, 9, 1896. https://doi.org/10.3390/foods9121896
Chiesa LM, Pavlovic R, Nobile M, Di Cesare F, Malandra R, Pessina D, Panseri S. Discrimination between Fresh and Frozen-Thawed Fish Involved in Food Safety and Fraud Protection. Foods. 2020; 9(12):1896. https://doi.org/10.3390/foods9121896
Chicago/Turabian StyleChiesa, Luca Maria, Radmila Pavlovic, Maria Nobile, Federica Di Cesare, Renato Malandra, Davide Pessina, and Sara Panseri. 2020. "Discrimination between Fresh and Frozen-Thawed Fish Involved in Food Safety and Fraud Protection" Foods 9, no. 12: 1896. https://doi.org/10.3390/foods9121896