Fatty Acid Composition and Volatile Profile of M. longissimus thoracis from Commercial Lambs Reared in Different Forage Systems

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals and Management
2.2. Slaughter and Sampling
2.3. Fatty Acid Composition
2.4. Volatile Compounds Analysis
2.5. Statistical Analysis
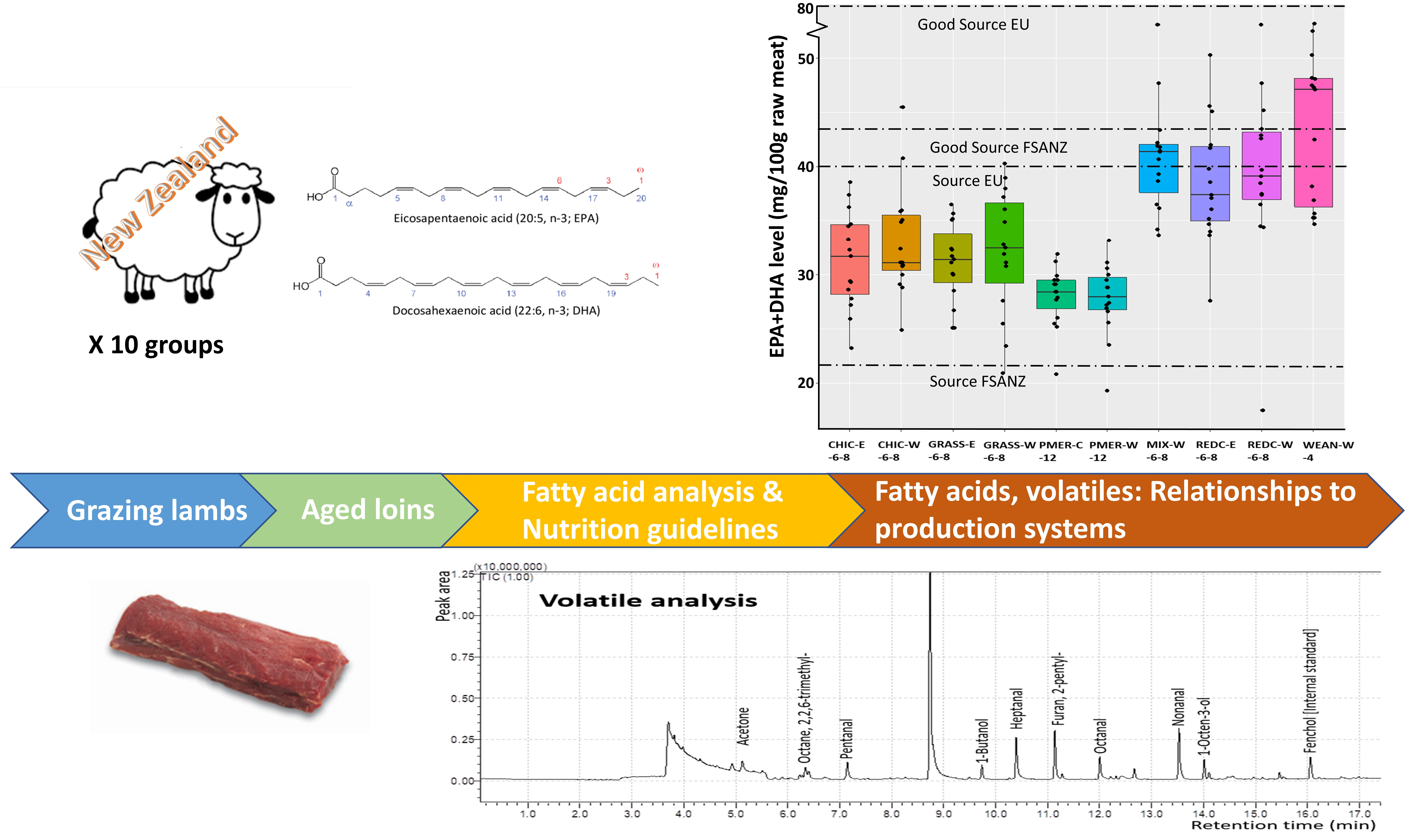
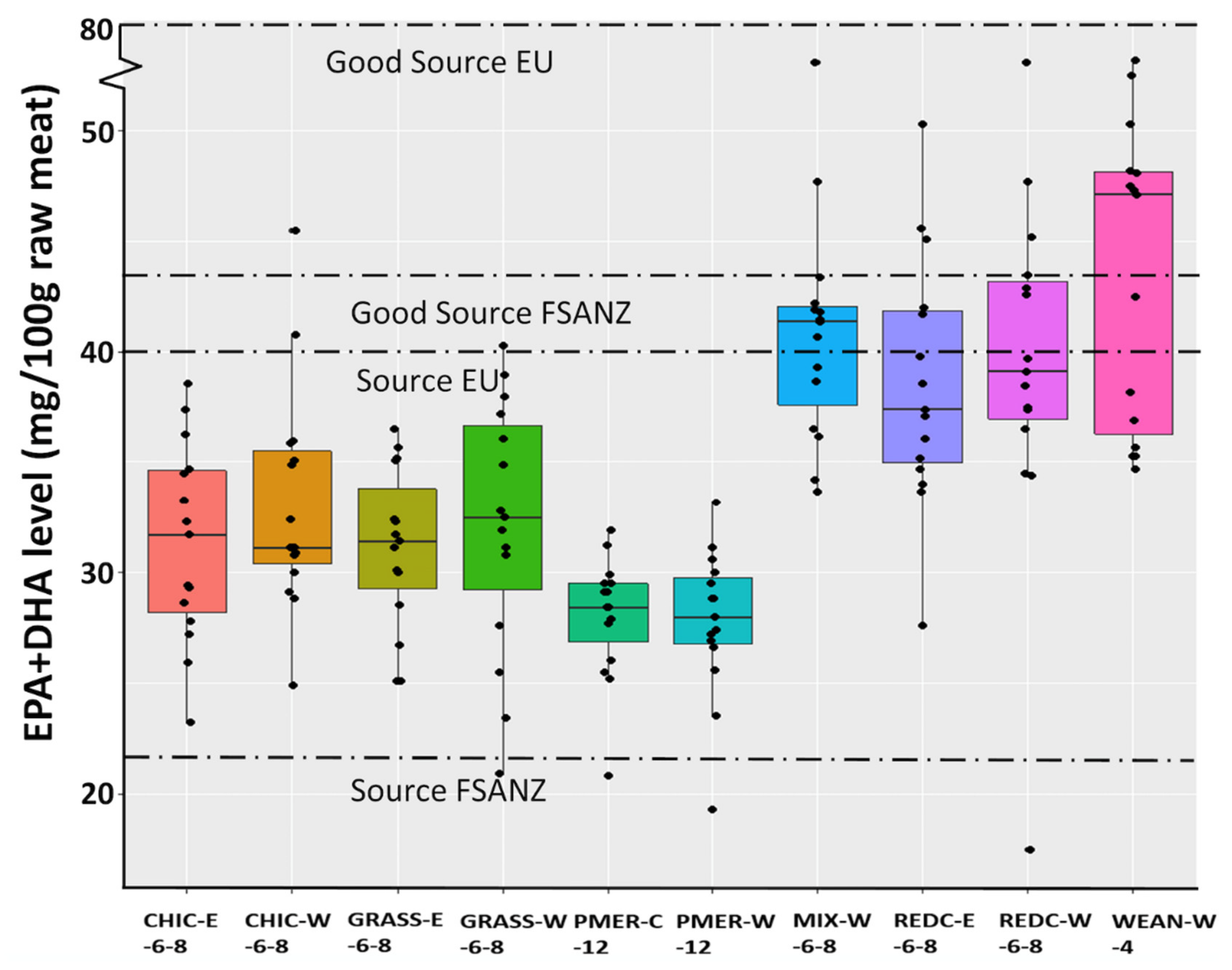
3. Results
3.1. Quantitative Fatty Acids
3.2. Fatty Acid as a Proportion of Total Fatty Acids
3.3. Nutrition Indexes
3.4. Volatiles from Raw Meat
3.4.1. Acids
3.4.2. Alcohols
3.4.3. Aldehydes
3.4.4. Ketones
3.4.5. Hydrocarbons and Furans
3.4.6. Sulphur Compounds
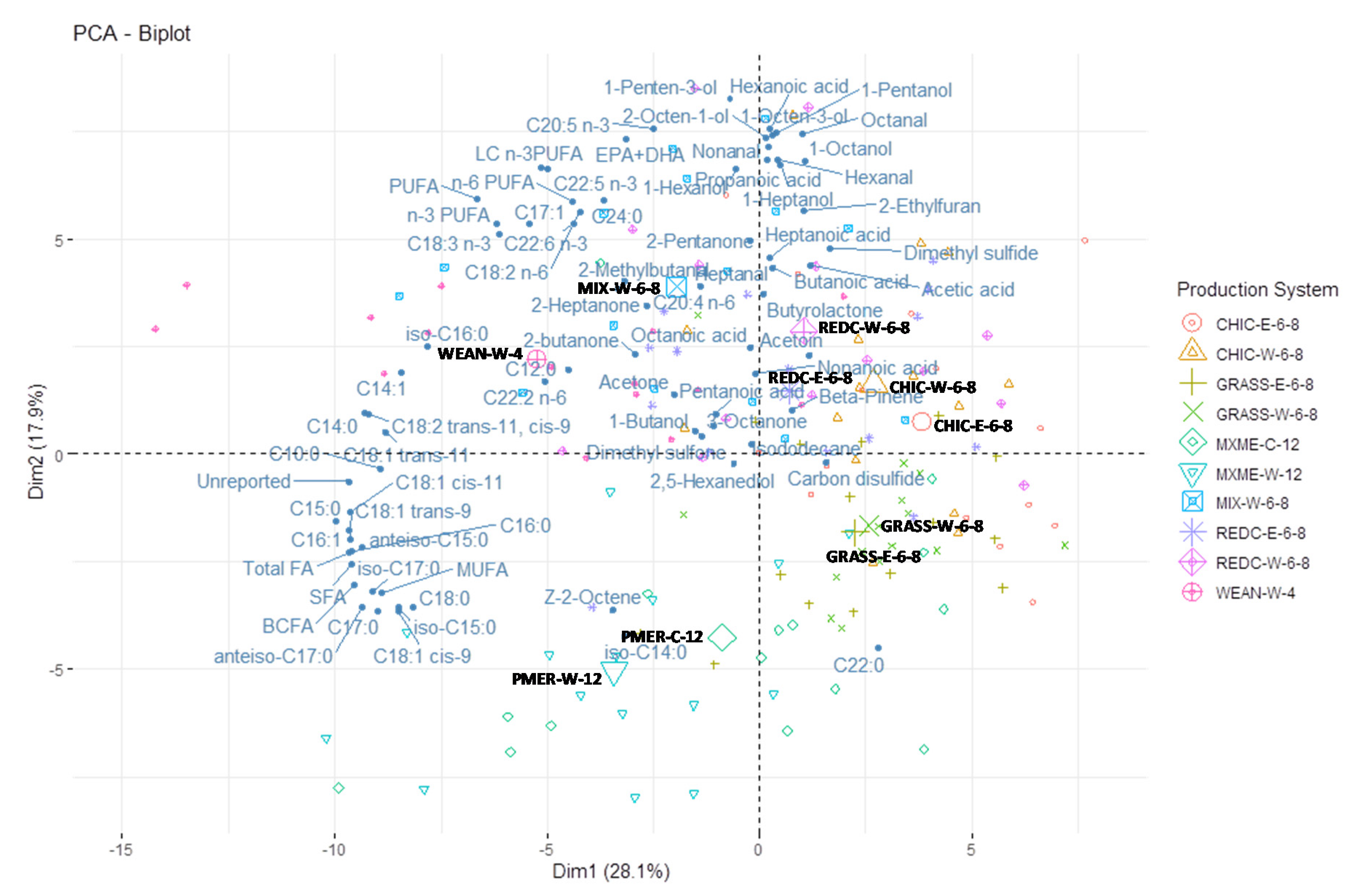
3.5. Principal Component Analysis
4. Discussion
4.1. Relationships between Production Systems, Fatty Acids and Lipid Volatiles
4.2. Non-Lipid Volatiles
4.3. Nutritional Quality: Selected Fatty Acids, Ratios and Indexes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pereira, P.M.; Vicente, A.F. Meat nutritional composition and nutritive role in the human diet. Meat Sci. 2013, 93, 586–592. [Google Scholar] [CrossRef] [Green Version]
- Scollan, N.; Hocquette, J.F.; Nuernberg, K.; Dannenberger, D.; Richardson, I.; Moloney, A. Innovations in beef production systems that enhance the nutritional and health value of beef lipids and their relationship with meat quality. Meat Sci. 2006, 74, 17–33. [Google Scholar] [CrossRef]
- McGuire, M.A.; McGuire, M.K. Conjugated linoleic acid (CLA): A ruminant fatty acid with beneficial effects on human health. J. Anim. Sci. 2000, 77, 1–8. [Google Scholar] [CrossRef]
- Hu, F.B.; Bronner, L.; Willett, W.C.; Stampfer, M.J.; Rexrode, K.M.; Albert, C.M.; Hunter, D.; Manson, J.E. Fish and omega-3 fatty acid intake and risk of coronary heart disease in women. J. Am. Med. Assoc. 2002, 287, 1815–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, A.J.; Ross, M.A. Fatty acid metabolism in neurodevelopmental disorder: A new perspective on associations between attention-deficit/hyperactivity disorder, dyslexia, dyspraxia and the autistic spectrum. Prostaglandins Leukot. Essent. Fat. Acids 2000, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; van Dam, R.M.; Liu, S. Diet and risk of Type II diabetes: The role of types of fat and carbohydrate. Diabetologia 2001, 44, 805–817. [Google Scholar] [CrossRef]
- United States, Dietary Guidelines Advisory Committee. Dietary Guidelines for Americans, 2010; Department of Health and Human Services: Washington, DC, USA; US Department of Agriculture: Washington, DC, USA, 2010.
- Díaz, M.T.; Álvarez, I.; De la Fuente, J.; Sañudo, C.; Campo, M.M.; Oliver, M.A.; Font i Furnols, M.; Montossi, F.; San Julián, R.; Nute, G.R.; et al. Fatty acid composition of meat from typical lamb production systems of Spain, United Kingdom, Germany and Uruguay. Meat Sci. 2005, 71, 256–263. [Google Scholar] [CrossRef]
- Fowler, S.M.; Morris, S.; Hopkins, D.L. Nutritional composition of lamb retail cuts from the carcases of extensively finished lambs. Meat Sci. 2019, 154, 126–132. [Google Scholar] [CrossRef]
- FSANZ. Food Standards Australia & New Zealand—Schedule 4—Nutrition, Health and Related Claims; Food Standards Australia New Zealand: Canberra, Australia, 2016; Volume F2016c0018. [Google Scholar]
- Commission Regulation of European Union. Amending regulation (EC) no 1924/2006 of the European parliament and of the council with regard to the list of nutrition claims. Off. J. Eur. Union 2010, 37, 16–18.
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Díaz, M.T.; Velasco, S.; Pérez, C.; Lauzurica, S.; Huidobro, F.; Cañeque, V. Physico-chemical characteristics of carcass and meat Manchego-breed suckling lambs slaughtered at different weights. Meat Sci. 2003, 65, 1247–1255. [Google Scholar] [CrossRef]
- Horcada, A.; Beriain, M.J.; Purroy, A.; Lizaso, G.; Chasco, J. Effect of sex on meat quality of Spanish lamb breeds (Lacha and Rasa Aragonesa). Anim. Sci. 1998, 67, 541–547. [Google Scholar] [CrossRef]
- Howes, N.L.; Bekhit, A.E.D.A.; Burritt, D.J.; Campbell, A.W. Opportunities and implications of pasture-based lamb fattening to enhance the long-chain fatty acid composition in meat. Compr. Rev. Food Sci. Food Saf. 2015, 14, 22–36. [Google Scholar] [CrossRef]
- Priolo, A.; Micol, D.; Agabriel, J. Effects of grass feeding systems on ruminant meat colour and flavour. A review. Anim. Res. 2001, 50, 185–200. [Google Scholar] [CrossRef]
- Resconi, V.C.; Escudero, A.; Campo, M.M. The development of aromas in ruminant meat. Molecules 2013, 18, 6748–6781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravador, R.S.; Serra, A.; Luciano, G.; Pennisi, P.; Vasta, V.; Mele, M.; Pauselli, M.; Priolo, A. Volatiles in raw and cooked meat from lambs fed olive cake and linseed. Animal 2015, 9, 715–722. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Schreurs, N.M.; Johnson, P.L.; Corner-thomas, R.A.; Agnew, M.P.; Silcock, P.; Eyres, G.T.; Maclennan, G.; Realini, C.E. Carcass characteristics and meat quality of commercial lambs reared in diff erent forage systems. Livest. Sci. 2020, 232, 103908. [Google Scholar] [CrossRef]
- Agnew, M.P.; Craigie, C.R.; Weralupitiya, G.; Reis, M.M.; Johnson, P.L.; Reis, M.G. Comprehensive evaluation of parameters affecting one-step method for quantitative analysis of fatty acids in meat. Metabolites 2019, 9, 189. [Google Scholar] [CrossRef] [Green Version]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Santos-Silva, J.; Bessa, R.J.; Santos-Silva, F. Effect of genotype, feeding system and slaughter weight on the quality of light lambs II. Fatty acid composition of meat. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Estévez, M.; Morcuende, D.; Ramírez, R.; Ventanas, J.; Cava, R. Extensively reared Iberian pigs versus intensively reared white pigs for the manufacture of liver pâté. Meat Sci. 2004, 67, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A.; Torchiano, M. lmPerm: Permutation tests for linear models. R Packag Version 210. PLoS Comput. Biol. 2016, 13, e1005752. [Google Scholar]
- Ihaka, R.; Gentleman, R. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. R Package Version 2017, 1, 337–354. [Google Scholar]
- Kęszycka, M.M.-; Czyżak, G.; Lipińska, P.; Wójtowski, J. Fatty acid profile of milk-a review. Bull. Vet. Inst. Pulawy 2013, 57, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Bas, P.; Morand-Fehr, P. Effect of nutritional factors on fatty acid composition of lamb fat deposits. Livest. Prod. Sci. 2000, 64, 61–79. [Google Scholar] [CrossRef]
- Salvatori, G.; Pantaleo, L.; Di Cesare, C.; Maiorano, G.; Filetti, F.; Oriani, G. Fatty acid composition and cholesterol content of muscles as related to genotype and vitamin E treatment in crossbred lambs. Meat Sci. 2004, 67, 45–55. [Google Scholar] [CrossRef]
- Warren, H.E.; Scollan, N.D.; Enser, M.; Hughes, S.I.; Richardson, R.I.; Wood, J.D. Effects of breed and a concentrate or grass silage diet on beef quality in cattle of 3 ages. I: Animal performance, carcass quality and muscle fatty acid composition. Meat Sci. 2008, 78, 256–269. [Google Scholar] [CrossRef]
- Calkins, C.R.; Hodgen, J.M. A fresh look at meat flavor. Meat Sci. 2007, 77, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Casaburi, A.; Piombino, P.; Nychas, G.J.; Villani, F.; Ercolini, D. Bacterial populations and the volatilome associated to meat spoilage. Food Microbiol. 2015, 45, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Broda, D.M.; Delacy, K.M.; Bell, R.G.; Penney, N. Association of psychrotrophic Clostridium spp. with deep tissue spoilage of chilled vacuum-packed lamb. Int. J. Food Microbiol. 1996, 29, 371–378. [Google Scholar] [CrossRef]
- Resconi, V.C.; Bueno, M.; Escudero, A.; Magalhaes, D.; Ferreira, V.; Campo, M.M. Ageing and retail display time in raw beef odour according to the degree of lipid oxidation. Food Chem. 2018, 242, 288–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almela, E.; Jordán, M.J.; Martínez, C.; Sotomayor, J.A.; Bedia, M.; Bañón, S. Ewe s diet (Pasture vs Grain-Based Feed) affects volatile profile of cooked meat from light lamb. J. Agric. Food Chem. 2010, 58, 9641–9646. [Google Scholar] [CrossRef]
- Elmore, J.S.; Cooper, S.L.; Enser, M.; Mottram, D.S.; Sinclair, L.A.; Wilkinson, R.G.; Wood, J.D. Dietary manipulation of fatty acid composition in lamb meat and its effect on the volatile aroma compounds of grilled lamb. Meat Sci. 2005, 69, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Dainty, R.H.; Edwards, R.A.; Hibbard, C.M. Spoilage of vacuum-packed beef by a clostridium sp. J. Sci. Food Agric. 1989, 49, 473–486. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Mortimer, S.I. Effect of genotype, gender and age on sheep meat quality and a case study illustrating integration of knowledge. Meat Sci. 2014, 98, 544–555. [Google Scholar] [CrossRef]
- Chevance, F.F.V.; Farmer, L.J. Identification of major volatile odor compounds in frankfurters. J. Agric. Food Chem. 1999, 47, 5151–5160. [Google Scholar] [CrossRef]
- Priolo, A.; Cornu, A.; Prache, S.; Krogmann, M.; Kondjoyan, N.; Micol, D.; Berdagué, J.L. Fat volatiles tracers of grass feeding in sheep. Meat Sci. 2004, 66, 475–481. [Google Scholar] [CrossRef]
- Cornu, A.; Kondjoyan, N.; Frencia, J.P.; Berdagué, J.L. Deciphering the message from volatile components of fat tissues. Viandes Prod. Carnés 2001, 22, 35–38. [Google Scholar]
- Mottram, D.S. Flavour formation in meat and meat products: A review. Food Chem. 1998, 62, 415–424. [Google Scholar] [CrossRef]
- Young, O.A.; Lane, G.A.; Priolo, A.; Fraser, K. Pastoral and species flavour in lambs raised on pasture, lucerne or maize. J. Sci. Food Agric. 2003, 83, 93–104. [Google Scholar] [CrossRef]
- Casaburi, A.; De Filippis, F.; Villani, F.; Ercolini, D. Activities of strains of Brochothrix thermosphacta in vitro and in meat. Food Res. Int. 2014, 62, 366–374. [Google Scholar] [CrossRef]
- Resconi, V.C.; Campo, M.M.; Montossi, F.; Ferreira, V.; Sañudo, C.; Escudero, A. Relationship between odour-active compounds and flavour perception in meat from lambs fed different diets. Meat Sci. 2010, 85, 700–706. [Google Scholar] [CrossRef]
- Frank, D.; Hughes, J.; Piyasiri, U.; Zhang, Y.; Kaur, M.; Li, Y.; Mellor, G.; Stark, J. Volatile and non-volatile metabolite changes in 140-day stored vacuum packaged chilled beef and potential shelf life markers. Meat Sci. 2019, 161, 108016. [Google Scholar] [CrossRef]
- Young, O.A.; Braggins, T.J. Flavor of Meat, Meat Products and Seafoods, 2nd ed.; Shahidi, F., Ed.; Blackie Academic: London, UK, 1998. [Google Scholar]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- De Lorgeril, M.; Renaud, S.; Salen, P.; Monjaud, I.; Mamelle, N.; Martin, J.L.; Guidollet, J.; Touboul, P.; Delaye, J. Mediterranean alpha-linolenic acid-rich diet in secondary prevention of coronary heart disease. Lancet 1994, 343, 1454–1459. [Google Scholar] [CrossRef]
- Gabriella, A.; Alessandro, D.; Palazzo, M.; Petrotos, K.; Goulas, P.; Martemucci, G. Fatty acid composition of light lamb meat from Leccese and Comisana dairy breeds as affected by slaughter age. Small Rumin. Res. 2015, 127, 36–43. [Google Scholar]
- Department of Health. Nutritional Aspects of Cardiovascular Disease. Report on Health and Social Subjects 46; HM Stationery Office: London, UK, 1994. [Google Scholar]
- World Health Organization. Interim Summary of Conclusions and Dietary Recommendations on Total Fat & Fatty Acids; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Hunter, J.E.; Zhang, J.; Kris-etherton, P.M. Cardiovascular disease risk of dietary stearic acid compared with trans, other saturated, and unsaturated fatty acids: A systematic review. Am. J. Clin. Nutr. 2010, 91, 46–63. [Google Scholar] [CrossRef] [Green Version]
- Bonanome, A.; Grundy, S.M. Effect of dietary stearic acid on plasma cholesterol and lipoprotein levels. N. Engl. J. Med. 1988, 318, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Russell, F.D.; Bürgin-Maunder, C.S. Distinguishing health benefits of eicosapentaenoic and docosahexaenoic acids. Mar. Drugs 2012, 10, 2535–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnampalam, E.N.; Butler, K.L.; Pearce, K.M.; Mortimer, S.I.; Pethick, D.W.; Ball, A.J.; Hopkins, D.L. Sources of variation of health claimable long chain omega-3 fatty acids in meat from Australian lamb slaughtered at similar weights. Meat Sci. 2014, 96, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, V.B.; Leite, R.F.; Almeida, M.T.C.; Paschoaloto, J.R.; Carvalho, E.B.; Lanna, D.P.D.; Perez, H.L.; Van Cleef, E.H.C.B.; Homem Junior, A.C.; Ezequiel, J.M.B. Carcass characteristics and meat quality of lambs fed high concentrations of crude glycerin in low-starch diets. Meat Sci. 2015, 110, 285–292. [Google Scholar] [CrossRef]
- Realini, C.E.; Bianchi, G.; Bentancur, O.; Garibotto, G. Effect of supplementation with linseed or a blend of aromatic spices and time on feed on fatty acid composition, meat quality and consumer liking of meat from lambs fed dehydrated alfalfa or corn. Meat Sci. 2017, 127, 21–29. [Google Scholar] [CrossRef]
- Geay, Y.; Bauchart, D.; Hocquette, J.-F.; Culioli, J. Effect of nutritional factors on biochemical, structural and metabolic characteristics of muscles in ruminants, consequences on dietetic value and sensorial qualities of meat. Reprod. Nutr. Dev. 2001, 41, 1–26. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Production System | Farm | Approximate Age at Slaughter (Months) | Sex | Diet 1 | Genetics |
|---|---|---|---|---|---|
| REDC-W-6-8 | A | 6–8 | Wethers | Red Clover | Perendale × LambSupreme 2 |
| REDC-E-6-8 | A | 6–8 | Ewes | Red Clover | Perendale × LambSupreme 2 |
| MIX-W-6-8 | A | 6–8 | Wethers | Mixed pasture | Perendale × Romney |
| PMER-W-12 | B | 12 | Wethers | Pasture | Merino |
| PMER-C-12 | B | 12 | Cryptorchids | Pasture | Merino |
| GRASS-W-6-8 | C | 6–8 | Wethers | Pasture | Composite 3 |
| GRASS-E-6-8 | C | 6–8 | Ewes | Pasture | Composite 3 |
| CHIC-W-6-8 | C | 6–8 | Wethers | Chicory | Composite 3 |
| CHIC-E-6-8 | C | 6–8 | Ewes | Chicory | Composite 3 |
| WEAN-W-4 | C | 4 | Wethers | Pre-weaning | Composite 3 |
| Fatty Acids | 4 Months Old | 6–8 Months Old | 12 Months Old | SEM 10 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WEAN-W-4 | GRASS-W-6-8 | GRASS-E-6-8 | CHIC-W-6-8 | CHIC-E-6-8 | REDC-W-6-8 | REDC-E-6-8 | MIX-W-6-8 | PMER-W-12 | PMER-C-12 | |||
| C10:0 | 6.64 c | 3.09 ab | 3.57 ab | 2.97 ab | 2.73 a | 3.17 ab | 3.09 ab | 3.73 ab | 4.31 b | 3.94 ab | 0.448 | <0.001 |
| C12:0 | 12.31 c | 6.23 ab | 6.05 ab | 4.96 ab | 7.00 b | 5.89 ab | 3.37 a | 5.45 ab | 4.03 ab | 3.47 ab | 1.097 | <0.001 |
| C14:0 | 122.15 b | 47.69 a | 52.23 a | 45.51 a | 41.49 a | 56.82 a | 46.82 a | 66.08 a | 69.03 a | 56.69 a | 9.76 | <0.001 |
| C15:0 | 13.08 d | 6.95 ab | 7.11 ab | 6.08 a | 5.31 a | 7.94 ab | 6.53 ab | 8.67 abc | 12.03 cd | 9.95 bcd | 1.107 | <0.001 |
| C16:0 | 645.84 bc | 423.61 a | 499.05 ab | 458.19 ab | 431.67 a | 503.66 ab | 497.20 ab | 579.09 abc | 733.65 c | 634.20 bc | 62.20 | <0.001 |
| C17:0 | 26.55 bc | 20.55 ab | 21.28 ab | 19.17 ab | 17.50 a | 22.99 abc | 21.63 ab | 25.95 abc | 37.37 d | 30.65 cd | 2.660 | <0.001 |
| C18:0 | 349.26 ab | 320.44 a | 333.71 a | 308.54 a | 297.93 a | 362.39 ab | 351.25 ab | 414.19 ab | 557.91 c | 468.19 bc | 40.48 | <0.001 |
| C20:0 1 | 0.66 a | 0.01 a | 0.15 a | bd | bd | 0.59 a | 0.22 a | 0.53 a | 3.48 b | 2.69 b | 0.285 | <0.001 |
| C22:0 | bd | 2.19 c | 0.87 a | 1.18 ab | 2.11 c | bd | bd | bd | 1.22 ab | 1.79 bc | 0.223 | <0.001 |
| C24:0 | 1.88 b | bd | bd | bd | bd | 1.39 a | 1.43 a | 1.35 a | bd | bd | 0.095 | <0.001 |
| iso-C14:0 | bd | 0.20 a | 0.09 ab | bd | bd | bd | bd | bd | 0.63 b | 0.40 ab | 0.145 | <0.001 |
| iso-C15:0 | 2.67 cd | 2.37 ab | 2.07 ab | 1.65 ab | 1.05 a | 2.17 abc | 1.56 | 2.46 | 4.38 | 3.71 | 0.464 | <0.001 |
| anteiso-C15:0 | 5.09 cd | 3.23 ab | 3.15 ab | 2.75 ab | 2.10 a | 3.50 abc | 2.70 a | 3.60 abc | 5.56 d | 4.30 bcd | 0.499 | <0.001 |
| iso-C16:0 | 3.91 | 1.98 | 1.86 | 1.78 | 1.75 | 2.69 | 2.23 | 2.12 | 2.16 | 2.16 | 0.310 | <0.001 |
| iso-C17:0 | 9.47 cd | 7.57 abc | 7.27 abc | 6.00 ab | 5.01 a | 8.31 bc | 7.23 abc | 9.64 cd | 13.19 e | 11.47 de | 0.938 | <0.001 |
| anteiso-C17:0 | 13.01 cd | 8.45 ab | 8.86 abc | 7.90 ab | 9.09 a | 8.11 abc | 10.85 ab | 10.85 bcd | 17.73 e | 13.75 de | 1.326 | <0.001 |
| BCFA 2 | 34.14 cde | 23.82 abc | 23.33 abc | 20.09 ab | 16.24 a | 25.78 abcd | 21.82 ab | 29.42 bcd | 43.59 e | 35.77 de | 3.416 | <0.001 |
| SFA 3 | 1213.86 bcd | 855.57 ab | 947.21 abc | 866.63a b | 821.99 a | 990.07 abc | 953.16 abc | 1133.91 abcd | 1463.15 d | 1244.67 cd | 116.1 | <0.001 |
| C14:1 | 4.07 b | 1.00 a | 1.14 a | 0.81 a | 0.75 a | 1.51 a | 1.17 a | 1.91 a | 1.46 a | 1.22 a | 0.430 | <0.001 |
| C16:1 | 35.82 c | 19.53 a | 23.70 ab | 19.94 a | 18.85 a | 23.85 ab | 22.85 ab | 27.99 abc | 34.85 c | 31.84 bc | 3.271 | <0.001 |
| C17:1 | 13.57 b | 3.73 a | 2.42 a | 3.90 a | 2.85 a | 10.93 b | 11.95 b | 14.15 b | 2.45 a | 3.47 a | 1.112 | <0.001 |
| C18:1 trans-9 | 5.41 c | 3.77 ab | 3.73 ab | 3.87 ab | 3.33 a | 4.77 ab | 4.50 ab | 5.29 bc | 6.73 c | 5.52 bc | 0.606 | <0.001 |
| C18:1 trans-11 | 73.36 c | 42.34 a | 48.71 ab | 54.54 abc | 45.91 ab | 69.80 abc | 62.51 abc | 66.83 abc | 78.06 c | 54.09 abc | 9.09 | <0.001 |
| C18:1 cis-9 | 816.06 abc | 672.16 ab | 742.11 abc | 676.69 ab | 625.41 a | 755.61 abc | 783.97 abc | 964.95 bcd | 1249.19 d | 1041.75 cd | 95.3 | <0.001 |
| C18:1 cis-11 | 27.56 bc | 20.26 a | 21.47 a | 21.09 a | 19.29 a | 24.58 ab | 24.28 ab | 27.98 bc | 31.39 c | 28.35 bc | 1.891 | <0.001 |
| C24:1 | bd | 0.01 c | 0.01 ab | 0.01 bc | 0.01 bc | bd | bd | bd | 0.01 c | 0.01 bc | 0.179 | <0.001 |
| MUFA4 | 975.88 abc | 762.79 ab | 843.23abc | 780.85 ab | 716.39 a | 891.04 abc | 911.27 abc | 1109.14 bcd | 1404.15 d | 1166.15 cd | 108.0 | <0.001 |
| C18:2n-6 | 148.49 e | 92.03 abc | 92.63 abc | 119.99 e | 115.69 de | 105.13 cde | 103.34 cde | 98.32 bcd | 80.28 ab | 74.76 a | 6.15 | <0.001 |
| C18:2 cis-9, trans-11 | 35.78 b | 18.73 a | 20.65 a | 22.45 ab | 18.64 a | 26.67 b | 23.93 ab | 30.44 ab | 31.43 ab | 21.35 a | 4.179 | <0.001 |
| C20:4n-6 | 36.85 ab | 37.05 ab | 31.46 a | 32.29 ab | 30.12 ab | 38.00 b | 36.29 ab | 38.21 b | 33.65 ab | 31.55 ab | 2.311 | <0.001 |
| C22:2n-6 | 0.78 b | bd | bd | bd | bd | 0.45 ab | 0.13 a | bd | bd | 0.17 a | 0.165 | <0.001 |
| C18:3n-3 | 84.05 d | 46.43 ab | 49.35 abc | 61.21 c | 58.16 bc | 61.44 c | 59.16 bc | 58.10 bc | 46.01 ab | 42.48 a | 4.167 | <0.001 |
| C20:5n-3 | 33.03 d | 24.65 ab | 23.92 ab | 26.29 bc | 25.05 ab | 30.87 cd | 30.44 cd | 31.89 d | 21.32 a | 21.54 ab | 1.491 | <0.001 |
| C22:5n-3 | 31.59 c | 23.94 a | 22.41 a | 23.37 a | 22.27 a | 28.29 bc | 27.51 b | 29.35 bc | 22.20 a | 21.83 a | 1.055 | <0.001 |
| C22:6n-3 | 10.49 d | 7.49 abc | 7.21 abc | 6.87 ab | 6.31 a | 8.47 bc | 8.15 abc | 8.93 cd | 6.43 a | 6.47 a | 0.587 | <0.001 |
| PUFA 5 | 381.08 e | 250.32 abc | 247.64 abc | 292.57 cd | 276.23 bcd | 299.33 d | 288.97 bcd | 295.25 cd | 241.32 ab | 220.17 a | 15.04 | <0.001 |
| EPA+DHA 6 | 43.52 c | 32.13 a | 31.13 a | 33.15 ab | 31.35 a | 39.34 bc | 38.59 bc | 40.82 c | 27.77 a | 28.01 a | 1.934 | <0.001 |
| n-6 PUFA sum 7 | 185.35 e | 129.09 abcd | 124.07 abc | 152.38 d | 145.81 cd | 143.13 cd | 139.64 cd | 136.54 bcd | 113.93 ab | 106.33 a | 7.61 | <0.001 |
| n-3 PUFA sum 8 | 159.18 d | 101.54 ab | 103.73 ab | 117.74 bc | 111.77 abc | 129.08 c | 125.27 c | 128.28 c | 95.97 a | 92.31 a | 6.26 | <0.001 |
| LC n-3 PUFA 9 | 75.11 c | 56.60 a | 55.02 a | 57.76 a | 54.66 a | 67.75 bc | 67.75 b | 70.17 bc | 51.31 a | 50.75 a | 2.805 | <0.001 |
| Unreported | 223.00 bc | 187.44 ab | 190.99 ab | 177.62 a | 174.03 a | 214.70 abc | 197.75 ab | 222.63 bc | 243.98 c | 223.42 bc | 12.95 | <0.001 |
| Total FA | 2792.48 bcd | 2084.40 ab | 2241.72 abc | 2118.91 abc | 1989.71 a | 2395.82 abc | 2351.36 abc | 2761.45 abcd | 3357.82 d | 2857.42 cd | 243.5 | <0.001 |
| Fatty Acids | 4 Months Old | 6–8 Months Old | 12 Months Old | SEM 9 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WEAN-W-4 | GRASS-W-6-8 | GRASS-E-6-8 | CHIC-W-6-8 | CHIC-E-6-8 | REDC-W-6-8 | REDC-E-6-8 | MIX-W-6-8 | PMER-W-12 | PMER-C-12 | |||
| C10:0 | 0.23 c | 0.15 ab | 0.16 b | 0.14 ab | 0.14 ab | 0.13 ab | 0.13 a | 0.13 ab | 0.13 a | 0.14 ab | 0.009 | <0.001 |
| C12:0 | 0.43 e | 0.33 cde | 0.27 bcde | 0.24 abcd | 0.38 de | 0.24 abcd | 0.14 ab | 0.19 abc | 0.12 a | 0.12 a | 0.047 | <0.001 |
| C14:0 | 4.26 b | 2.31 a | 2.32 a | 2.14 a | 2.02 a | 2.24 a | 1.95 a | 2.34 a | 2.06 a | 1.91 a | 0.215 | <0.001 |
| C15:0 | 0.46 e | 0.34 cd | 0.32 abcd | 0.29 abc | 0.27a | 0.32 abcd | 0.28 ab | 0.31 abcd | 0.36 d | 0.34 bcd | 0.021 | <0.001 |
| C16:0 | 23.01 b | 20.42 a | 22.05 ab | 21.39 ab | 21.43 ab | 20.59 a | 20.89 a | 20.82 a | 21.74 ab | 21.91 ab | 0.548 | <0.001 |
| C17:0 | 0.95 ab | 0.99 bc | 0.96 ab | 0.91 a | 0.88 a | 0.95 ab | 0.92 ab | 0.94 ab | 1.11 d | 1.07 cd | 0.027 | <0.001 |
| C18:0 | 12.61 a | 15.55 bcd | 15.00 bc | 14.56 b | 14.99 bc | 14.99 bc | 14.89 b | 15.01 bc | 16.61 d | 16.38 cd | 0.441 | <0.001 |
| C20:0 | 0.0229 | 0.0003 | 0.0051 | - | - | 0.0214 | 0.0080 | 0.0181 | 0.1118 | 0.0971 | 0.008 | <0.001 |
| C22:0 | - | 0.1119 cd | 0.0624 b | 0.0634 b | 0.1237 d | - | - | - | 0.0385 ab | 0.0753 bc | 0.012 | <0.001 |
| C24:0 | 0.0757 c | - | - | - | - | 0.0717 bc | 0.0710 bc | 0.0555 b | - | - | 0.006 | <0.001 |
| iso-C14:0 | - | 0.0119 ab | 0.0039 a | - | - | - | - | - | 0.0180 b | 0.0116 ab | 0.005 | <0.001 |
| iso-C15:0 | 0.10 bcd | 0.13 cd | 0.11 bcd | 0.09 abc | 0.05 a | 0.09 abc | 0.07 ab | 0.09 abcd | 0.14 d | 0.14 d | 0.014 | <0.001 |
| anteiso-C15:0 | 0.18 d | 0.16 cd | 0.15 bcd | 0.13 abc | 0.10 a | 0.14 abcd | 0.11 ab | 0.13 abc | 0.17 cd | 0.15 bcd | 0.013 | <0.001 |
| iso-C16:0 | 0.14 c | 0.10 b | 0.09 ab | 0.09 ab | 0.09 ab | 0.11 b | 0.09 ab | 0.10 b | 0.07 a | 0.08 ab | 0.009 | <0.001 |
| iso-C17:0 | 0.33 bcd | 0.37 cde | 0.33 bc | 0.29 ab | 0.25 a | 0.35 bcde | 0.31 ab | 0.35 bcde | 0.39 de | 0.40 e | 0.019 | <0.001 |
| anteiso-C17:0 | 0.46 def | 0.41 cde | 0.40 bcd | 0.37 abc | 0.31a | 0.37 abc | 0.34 ab | 0.39 bc | 0.53 f | 0.48 ef | 0.021 | <0.001 |
| BCFA 1 | 1.20 cd | 1.17 cd | 1.07 bc | 0.95 ab | 0.81 a | 1.04 bc | 0.92 ab | 1.06 bc | 1.30 d | 1.25 cd | 0.067 | <0.001 |
| SFA 2 | 43.24 bc | 41.38 abc | 42.20 abc | 40.68 ab | 41.01 ab | 40.58 ab | 40.19 a | 40.86 ab | 43.56 c | 43.28 c | 0.697 | <0.001 |
| C14:1 | 0.14 b | 0.05 a | 0.05 a | 0.04 a | 0.03 a | 0.06 a | 0.05 a | 0.07 a | 0.04 a | 0.04 a | 0.014 | <0.001 |
| C16:1 | 1.26 c | 0.94 ab | 1.04 ab | 0.92 a | 0.94 a | 0.97 ab | 0.96 ab | 1.00 ab | 1.04 ab | 1.12 bc | 0.055 | <0.001 |
| C17:1 | 0.48 c | 0.19 b | 0.11 ab | 0.19 b | 0.15 ab | 0.47 c | 0.51 c | 0.52 c | 0.08 a | 0.12 ab | 0.031 | <0.001 |
| C18:1 trans-9 | 0.19 ab | 0.18 ab | 0.16 a | 0.18 ab | 0.16 a | 0.19 ab | 0.19 ab | 0.19 ab | 0.20 b | 0.19 ab | 0.010 | <0.001 |
| C18:1 trans-11 | 2.62 b | 2.08 ab | 2.19 ab | 2.53 ab | 2.24 ab | 2.78 b | 2.65 b | 2.40 ab | 2.33 ab | 1.89 a | 0.214 | <0.001 |
| C18:1 cis-9 | 29.21 a | 32.26 abc | 32.70 bc | 31.55 ab | 31.06 ab | 31.15 ab | 33.02 bc | 34.74 cd | 37.04 d | 36.17 d | 0.953 | <0.001 |
| C18:1 cis-11 | 1.00 ab | 1.00 ab | 0.98 ab | 1.01 ab | 0.99 ab | 1.06 ab | 1.04 ab | 1.02 ab | 0.94 a | 1.01 ab | 0.033 | <0.001 |
| C24:1 | - | 0.0801 e | 0.0339 bc | 0.0655 de | 0.0575 cde | - | - | - | 0.0440 cd | 0.0383 cd | 0.009 | <0.001 |
| MUFA 3 | 34.89 a | 36.76 abc | 37.26 bcd | 36.48 abc | 35.62 ab | 36.67 abc | 38.42 cde | 39.94 def | 41.72 f | 40.57 ef | 0.935 | <0.001 |
| C18:2n-6 | 5.41 def | 4.66 cd | 4.42 cd | 5.85 ef | 6.03 f | 4.72 cde | 4.57 cd | 3.63 bc | 2.46 a | 2.71 ab | 0.353 | <0.001 |
| C18:2 cis-9, trans-11 | 1.25 c | 0.91 ab | 0.92 ab | 1.03 abc | 0.90 ab | 1.05 abc | 1.00 abc | 1.08 bc | 0.94 abc | 0.74 a | 0.100 | <0.001 |
| C20:4n-6 | 1.36 abc | 1.90 d | 1.52 abcd | 1.60 bcd | 1.58 abcd | 1.77 cd | 1.62 bcd | 1.45 abcd | 1.04 a | 1.18 ab | 0.172 | <0.001 |
| C22:2n-6 | 0.0289 b | - | - | - | - | 0.0142 ab | 0.0053 ab | - | - | 0.0068 ab | 0.005 | <0.001 |
| C18:3n-3 | 3.03 d | 2.30 bc | 2.32 bc | 2.99 d | 3.03 d | 2.71 cd | 2.59 bcd | 2.13 b | 1.39 a | 1.52 a | 0.462 | <0.001 |
| C20:5n-3 | 1.21 d | 1.23 bc | 1.13 bc | 1.30 d | 1.33 d | 1.43 cd | 1.37 bcd | 1.21 b | 0.66 a | 0.82 a | 0.169 | <0.001 |
| C22:5n-3 | 1.16 c | 1.21 bc | 1.06 bc | 1.14 c | 1.18 c | 1.30 c | 1.23 c | 1.11 c | 0.68 a | 0.82 ab | 0.119 | <0.001 |
| C22:6n-3 | 0.38 c | 0.38 c | 0.34 bc | 0.34 bc | 0.33 bc | 0.39 c | 0.37 c | 0.34 bc | 0.20 a | 0.24 ab | 0.036 | <0.001 |
| PUFA 4 | 13.83 c | 12.59 bc | 11.70 bc | 14.24 c | 14.37 c | 13.38 bc | 12.76 bc | 10.95 b | 7.36 a | 8.04 a | 0.821 | <0.001 |
| EPA+DHA 5 | 1.59 c | 1.61 c | 1.47 bc | 1.64 c | 1.66 c | 1.82 c | 1.74 c | 1.55 c | 0.86 a | 1.06 ab | 0.151 | <0.001 |
| n-6 PUFA 6 | 6.80 cd | 6.56 bcd | 5.94 bc | 7.44 cd | 7.62 d | 6.50 bcd | 6.20 bcd | 5.08 ab | 3.50 a | 3.90 a | 0.501 | <0.001 |
| n-3 PUFA 7 | 5.78 b | 5.12 b | 4.85 b | 5.77 b | 5.86 b | 5.83 b | 5.56 b | 4.79 b | 2.93 a | 3.41 a | 0.390 | <0.001 |
| LC n-3PUFA 8 | 2.75 c | 2.82 c | 2.53 bc | 2.78 c | 2.83 c | 3.12 c | 2.97 c | 2.66 bc | 1.54 a | 1.88 ab | 0.146 | <0.001 |
| Unreported | 8.04 ab | 9.28 a | 8.84 a | 8.60 ab | 9.00 a | 9.67 a | 8.63 ab | 8.25 ab | 7.36 a | 8.12 ab | 0.425 | <0.001 |
| Atty Acids | 4 Months Old | 6–8 Months Old | 12 Months Old | SEM 5 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WEAN-W-4 | GRASS-W-6-8 | GRASS-E-6-8 | CHIC-W-6-8 | CHIC-E-6-8 | REDC-W-6-8 | REDC-E-6-8 | MIX-W-6-8 | PMER -W-12 | PMER -C-12 | |||
| PUFA:SFA | 0.32 bc | 0.31 bc | 0.28 bc | 0.35 c | 0.35 c | 0.33 bc | 0.32 bc | 0.27 b | 0.17 a | 0.19 a | 0.02 | <0.001 |
| n-6:n-3 | 1.17 abc | 1.30 bc | 1.23 abc | 1.30 bc | 1.32 c | 1.11 ab | 1.12 abc | 1.06 a | 1.19 abc | 1.15 abc | 0.06 | <0.001 |
| Thrombogenic index 1 | 0.97 a | 0.95 a | 0.98 a | 0.89 a | 0.90 a | 0.89 a | 0.89 a | 0.95 a | 1.19 b | 1.14 b | 0.05 | <0.001 |
| Atherogenic index 2 | 0.86 b | 0.62 a | 0.66 a | 0.61 a | 0.61 a | 0.61 a | 0.58 a | 0.61 a | 0.63 a | 0.62 a | 0.03 | <0.001 |
| Nutritional ratio 3 | 0.80 b | 0.63 a | 0.66 a | 0.64 a | 0.64 a | 0.64 a | 0.61 a | 0.61 a | 0.61 a | 0.62 a | 0.01 | <0.001 |
| hypocholesterolaemic/Hypercholesterolaemic ratio 4 | 1.54 a | 1.94 b | 1.80 b | 1.91 b | 1.92 b | 1.92 b | 1.98 b | 1.93 b | 1.83 b | 1.84 b | 0.02 | <0.001 |
| Compounds | Calculated RI 1 | Ion Used | 4 Months Old | 6–8 Months Old | 12 Months Old | SEM 2 | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WEAN-W-4 | GRASS-W-6-8 | GRASS-E-6-8 | CHIC-W-6-8 | CHIC-E-6-8 | REDC-W-6-8 | REDC-E-6-8 | MIX-W-6-8 | PMER-W-12 | PMER-C-12 | |||||
| Acids | ||||||||||||||
| Acetic acid | 1483 | 43 | 6.29 b | 5.55 ab | 5.93 b | 6.19 b | 6.62 b | 6.06 b | 6.27 b | 5.48 ab | 4.46 ab | 3.60 a | 0.48 | 0.010 |
| Propanoic acid | 1566 | 74 | 0.41 b | 0.31 ab | 0.35 ab | 0.41 b | 0.39 b | 0.40 b | 0.35 ab | 0.45 b | 0.23 a | 0.20 a | 0.03 | <0.001 |
| Butanoic acid | 1654 | 60 | 3.43 c | 3.41 c | 2.28 abc | 2.82 bc | 3.02 c | 3.32 c | 3.16 c | 2.98 c | 1.54 ab | 1.33 a | 0.30 | <0.001 |
| Pentanoic acid | 1765 | 86 | 0.91 | 0.18 | 0.22 | 0.09 | 0.32 | 0.12 | 0.24 | 0.16 | 0.20 | 0.09 | 0.19 | 0.740 |
| Hexanoic acid | 1872 | 60 | 3.43 bcd | 2.72 abc | 2.77 abc | 4.03 d | 3.59 cd | 3.35 abcd | 3.12 abcd | 4.13 d | 2.43 ab | 2.28 a | 0.24 | <0.001 |
| Heptanoic acid | 1979 | 60 | 0.43 | 0.32 | 0.40 | 0.41 | 0.39 | 0.38 | 0.41 | 0.44 | 0.33 | 0.33 | 0.03 | 0.336 |
| Octanoic acid | 2086 | 60 | 0.80 | 0.47 | 0.64 | 0.64 | 0.55 | 0.55 | 0.55 | 0.61 | 0.56 | 0.58 | 0.05 | 0.059 |
| Nonanoic acid | 2193 | 73 | 0.34 | 0.25 | 0.35 | 0.37 | 0.40 | 0.34 | 0.50 | 0.44 | 0.30 | 0.33 | 0.05 | 0.180 |
| Alcohols | ||||||||||||||
| 1-Butanol | 1140 | 56 | 0.66 abcd | 0.36 a | 0.39 a | 0.52 abc | 0.47 ab | 0.52 abc | 0.42 a | 0.97 d | 0.85 bcd | 0.93 cd | 0.09 | <0.001 |
| 1-Penten-3-ol | 1155 | 57 | 6.47 ab | 5.92 ab | 5.37 a | 11.56 cd | 9.87 bc | 11.56 cd | 8.70 abc | 14.49 d | 4.70 a | 6.20 ab | 0.89 | <0.001 |
| 1-Pentanol | 1244 | 42 | 2.69 a | 4.06 ab | 3.47 a | 6.61 c | 5.91 bc | 5.54 bc | 4.46 ab | 7.12 c | 2.70 a | 3.38 a | 0.43 | <0.001 |
| 1-Hexanol | 1347 | 56 | 1.42 a | 1.41 a | 1.41 a | 2.66 cd | 2.36 abcd | 2.63 bcd | 1.97 abc | 3.31 d | 1.54 ab | 2.02 abc | 0.25 | <0.001 |
| 2,5-Hexanediol | 1418 | 43 | 0.13 | 0.13 | 0.14 | 0.15 | 0.16 | 0.14 | 0.14 | 0.14 | 0.13 | 0.14 | 0.01 | 0.945 |
| 1-Octen-3-ol | 1442 | 57 | 4.07 ab | 4.54 ab | 3.57 a | 8.01 c | 6.64 bc | 6.39 bc | 4.72 ab | 8.27 c | 3.64 a | 4.50 ab | 0.60 | <0.001 |
| 1-Heptanol | 1449 | 70 | 0.27 a | 0.46 abc | 0.48 abc | 0.68 cd | 0.67 cd | 0.75 de | 0.59 bcd | 1.00 e | 0.38 ab | 0.50 abcd | 0.06 | <0.001 |
| 1-Octanol | 1552 | 56 | 0.26 a | 0.42 abc | 0.46 abcd | 0.54 bcd | 0.58 cd | 0.65 de | 0.52 bcd | 0.80 e | 0.34 ab | 0.42 abc | 0.04 | <0.001 |
| 2-Octen-1-ol | 1608 | 57 | 0.17 ab | 0.20 ab | 0.15 a | 0.31 c | 0.25 bc | 0.25 abc | 0.19 ab | 0.32 c | 0.15 a | 0.18 ab | 0.02 | <0.001 |
| Aldehydes | ||||||||||||||
| 2-Methylbutanal | 910 | 43 | 2.14 ef | 1.74 abcd | 1.60 abc | 1.43 a | 1.56 ab | 2.26 f | 2.04 def | 2.70 g | 1.95 cdef | 1.81 bcde | 0.08 | <0.001 |
| Hexanal | 1089 | 43 | 1.47 abc | 0.68 a | 0.54 a | 4.35 d | 2.47 bc | 2.64 bc | 1.12 ab | 2.98 cd | 0.46 a | 0.73 a | 0.34 | <0.001 |
| Heptanal | 1193 | 57 | 0.08 | 0.07 | 0.07 | 0.09 | 0.09 | 0.11 | 0.09 | 0.11 | 0.09 | 0.09 | 0.01 | 0.050 |
| Octanal | 1295 | 57 | 0.12 a | 0.12 a | 0.11 a | 0.32 c | 0.25 bc | 0.29 c | 0.17 ab | 0.32 c | 0.08 a | 0.13 a | 0.02 | <0.001 |
| Nonanal | 1404 | 57 | 0.92 a | 0.82 a | 0.86 a | 2.15 d | 1.76 bcd | 1.95 cd | 1.25 abc | 2.41 d | 0.89 a | 1.04 ab | 0.18 | <0.001 |
| Ketones | ||||||||||||||
| Acetone | 826 | 70 | 33.93 cd | 31.82 cd | 29.65 bcd | 21.78 ab | 17.96 a | 45.88 e | 37.68 cde | 39.32 de | 28.31 bc | 32.92 cd | 2.16 | <0.001 |
| 2-Butanone | 911 | 72 | 0.56 de | 0.46 abcd | 0.39 abc | 0.34 a | 0.37 ab | 0.61 ef | 0.55 de | 0.71 f | 0.54 cde | 0.50 bcde | 0.03 | <0.001 |
| 2-Pentanone | 987 | 43 | 2.31 d | 1.22 ab | 1.18 a | 2.11 cd | 2.20 cd | 2.30 d | 2.53 d | 1.92 bcd | 1.24 ab | 1.56 abc | 0.16 | <0.001 |
| 2-Heptanone | 1187 | 43 | 2.10 ef | 1.73 abcd | 1.61 abc | 1.43 a | 1.56 ab | 2.26 f | 2.04 def | 2.70 g | 1.95 cdef | 1.84 bcde | 0.08 | <0.001 |
| 3-Octanone | 1260 | 42 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | <0.01 | 0.653 |
| Acetoin | 1304 | 45 | 2.10 ab | 2.29 ab | 2.54 abc | 7.55 bcd | 8.15 cd | 6.26 abcd | 8.91 d | 1.25 a | 1.99 ab | 2.46 abc | 1.29 | 0.008 |
| Butyrolactone | 1680 | 42 | 2.45 c | 2.99 c | 2.22 bc | 2.25 bc | 2.49 c | 2.97 c | 2.82 c | 4.32 d | 1.16 a | 1.41 ab | 0.23 | <0.001 |
| Hydrocarbons | ||||||||||||||
| Z-2-Octene | 873 | 43 | 0.06 bc | 0.02 ab | 0.02 ab | 0.02 ab | 0.02 ab | 0.01 a | 0.02 abc | 0.02 ab | 0.07 c | 0.13 d | 0.01 | <0.001 |
| Isododecane | 953 | 57 | 0.70 | 1.24 | 2.02 | 1.04 | 1.27 | 1.34 | 1.91 | 2.01 | 1.51 | 1.28 | 0.30 | 0.088 |
| Beta-Pinene | 1110 | 93 | 0.30 a | 0.78 ab | 0.94 b | 0.57 ab | 0.75 ab | 0.99 b | 1.06 b | 1.01 b | 0.67 ab | 0.60 ab | 0.12 | <0.001 |
| Furan | ||||||||||||||
| 2-Ethylfuran | 959 | 81 | 0.32 ab | 0.26 ab | 1.31 e | 1.06 cde | 0.66 abcd | 0.78 bcde | 0.53 abc | 1.23 de | 0.14 a | 0.35 ab | 0.13 | <0.001 |
| Sulphur compounds | ||||||||||||||
| Carbon disulphide | 745 | 76 | 17.04 | 16.73 | 14.54 | 15.93 | 21.03 | 12.98 | 13.10 | 14.72 | 15.33 | 20.52 | 4.09 | 0.987 |
| Dimethyl sulphide | 959 | 62 | 0.77 ab | 0.93 ab | 1.14 bc | 1.44 bcd | 2.02 d | 1.79 cd | 1.92 cd | 2.20 d | 0.18 a | 0.14 a | 0.19 | <0.001 |
| Dimethyl sulfone | 1939 | 79 | 7.33 bcde | 4.48 ab | 6.30 abcd | 4.84 abc | 3.68 a | 12.44 f | 10.11 ef | 6.11 abcd | 8.00 cde | 8.14 de | 0.72 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, Y.; Eyres, G.T.; Reis, M.G.; Schreurs, N.M.; Silcock, P.; Agnew, M.P.; Johnson, P.L.; Maclean, P.; Realini, C.E. Fatty Acid Composition and Volatile Profile of M. longissimus thoracis from Commercial Lambs Reared in Different Forage Systems. Foods 2020, 9, 1885. https://doi.org/10.3390/foods9121885
Ye Y, Eyres GT, Reis MG, Schreurs NM, Silcock P, Agnew MP, Johnson PL, Maclean P, Realini CE. Fatty Acid Composition and Volatile Profile of M. longissimus thoracis from Commercial Lambs Reared in Different Forage Systems. Foods. 2020; 9(12):1885. https://doi.org/10.3390/foods9121885
Chicago/Turabian StyleYe, Yangfan, Graham T. Eyres, Mariza G. Reis, Nicola M. Schreurs, Patrick Silcock, Michael P. Agnew, Patricia L. Johnson, Paul Maclean, and Carolina E. Realini. 2020. "Fatty Acid Composition and Volatile Profile of M. longissimus thoracis from Commercial Lambs Reared in Different Forage Systems" Foods 9, no. 12: 1885. https://doi.org/10.3390/foods9121885