Effectiveness of Sprayed Bioactive Fruit Extracts in Counteracting Protein Oxidation in Lamb Cutlets Subjected to a High-Oxygen MAP


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Raw Material
2.2. Extraction and Characterization of Fruit Extracts
2.3. In Vitro Bioactivities of Fruit Extracts
2.4. Preparation of Sprayable Extracts
2.5. Experimental Setting and Meat Processing and Sampling
2.6. Analytical Procedures on Lamb Cutlets
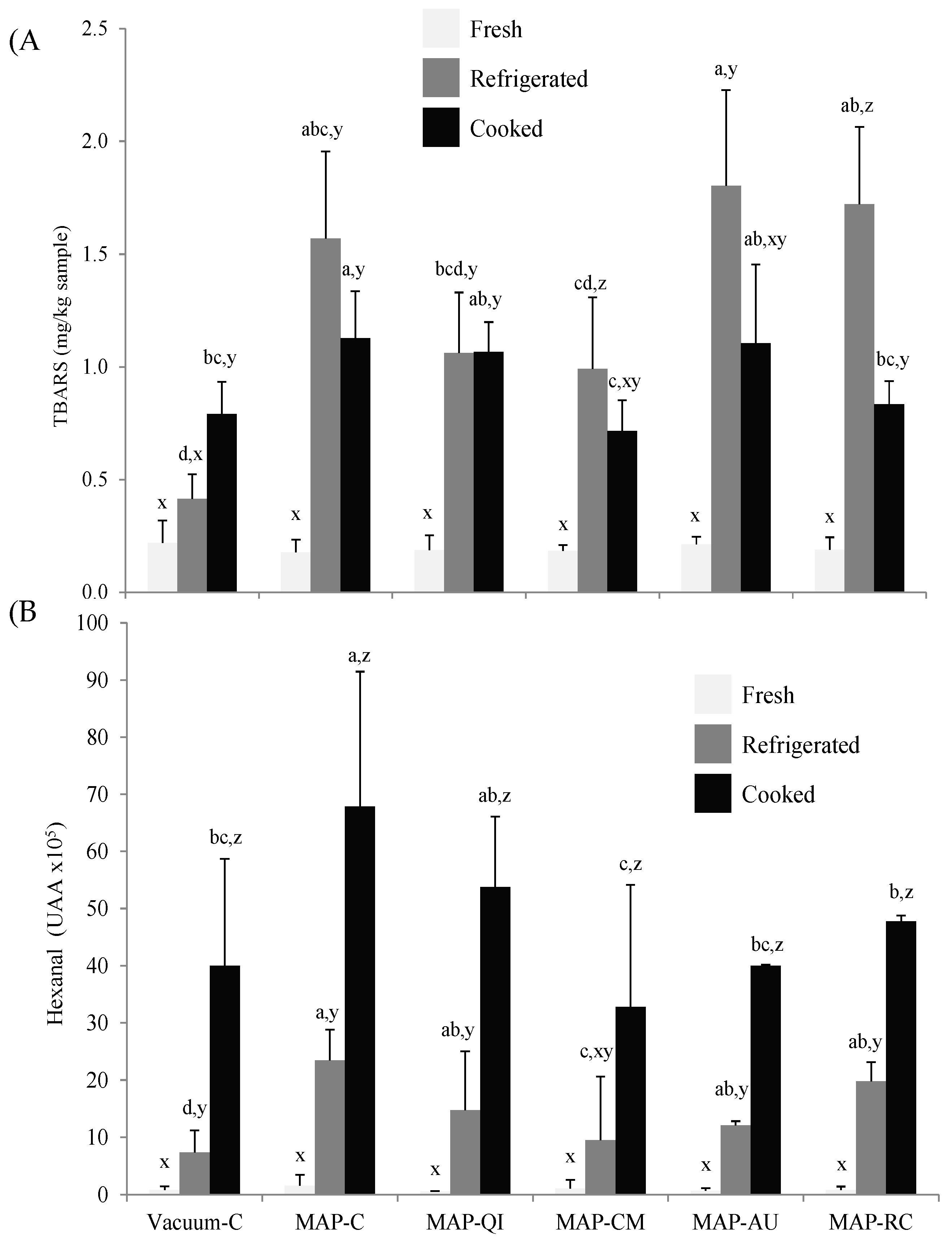
2.6.1. Determination of TBARS Numbers
2.6.2. Analysis of Hexanal
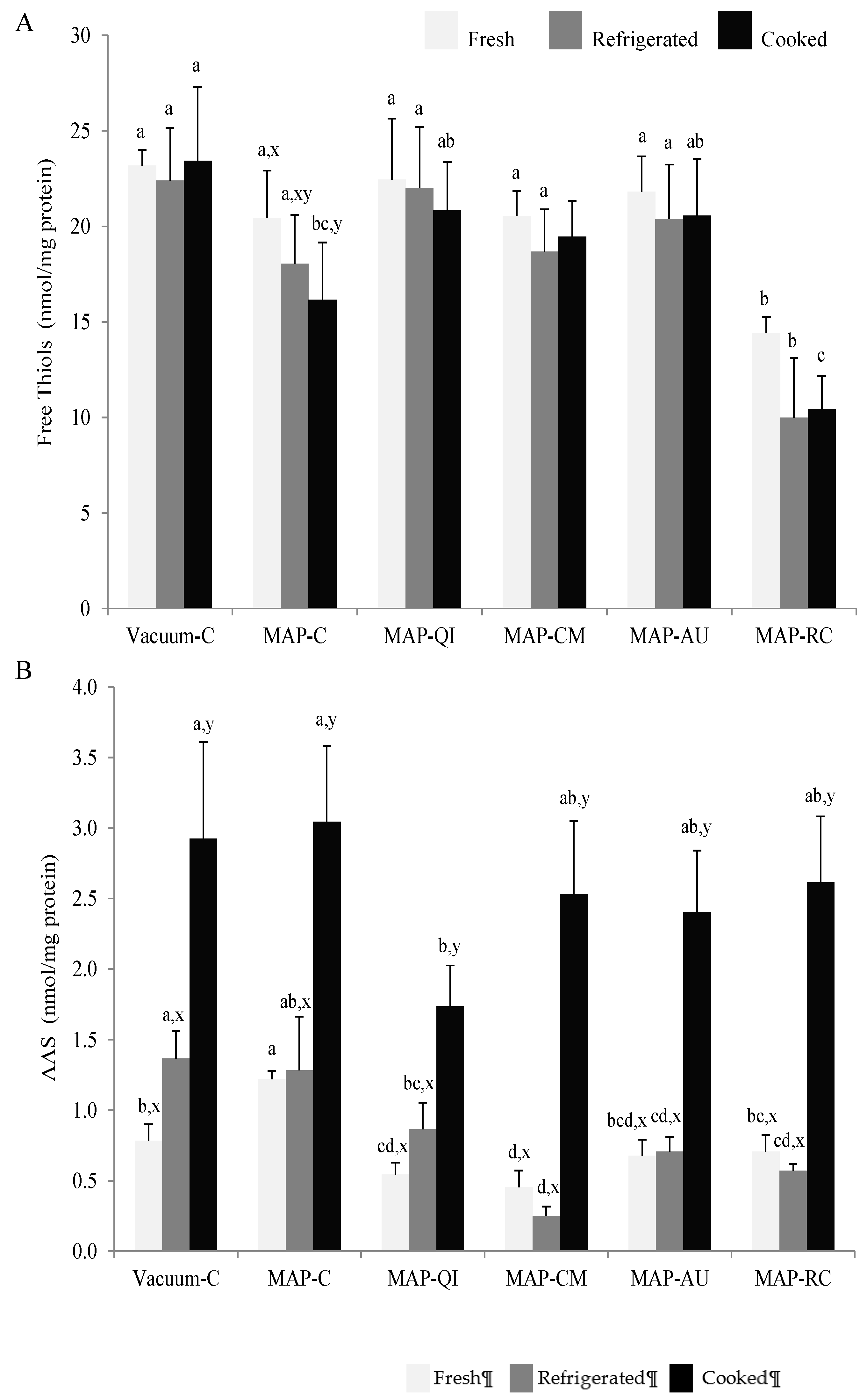
2.6.3. Quantification of α-amino Adipic Semialdehyde (AAS)
2.6.4. Analysis of Protein Thiols
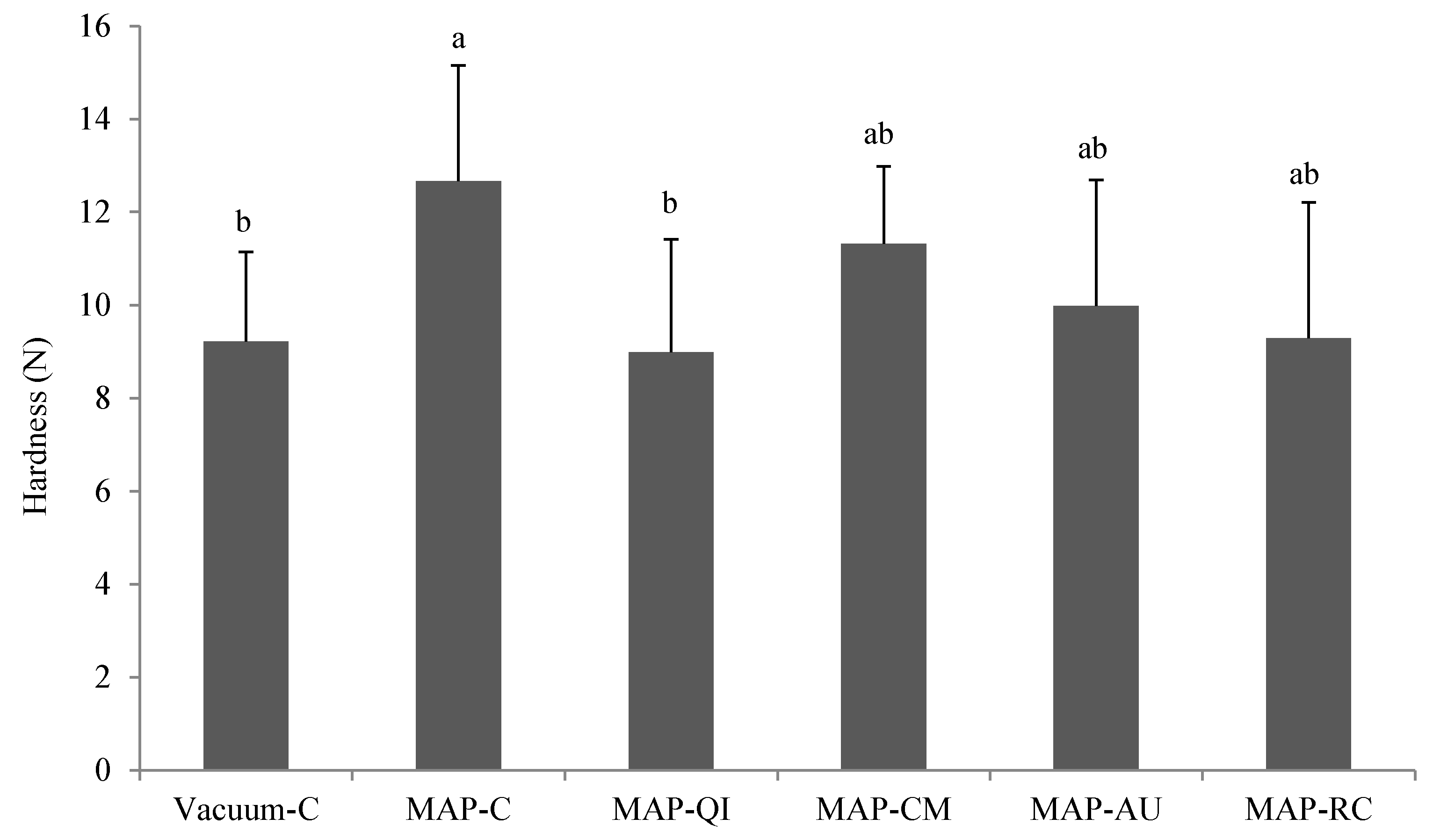
2.6.5. Instrumental Hardness
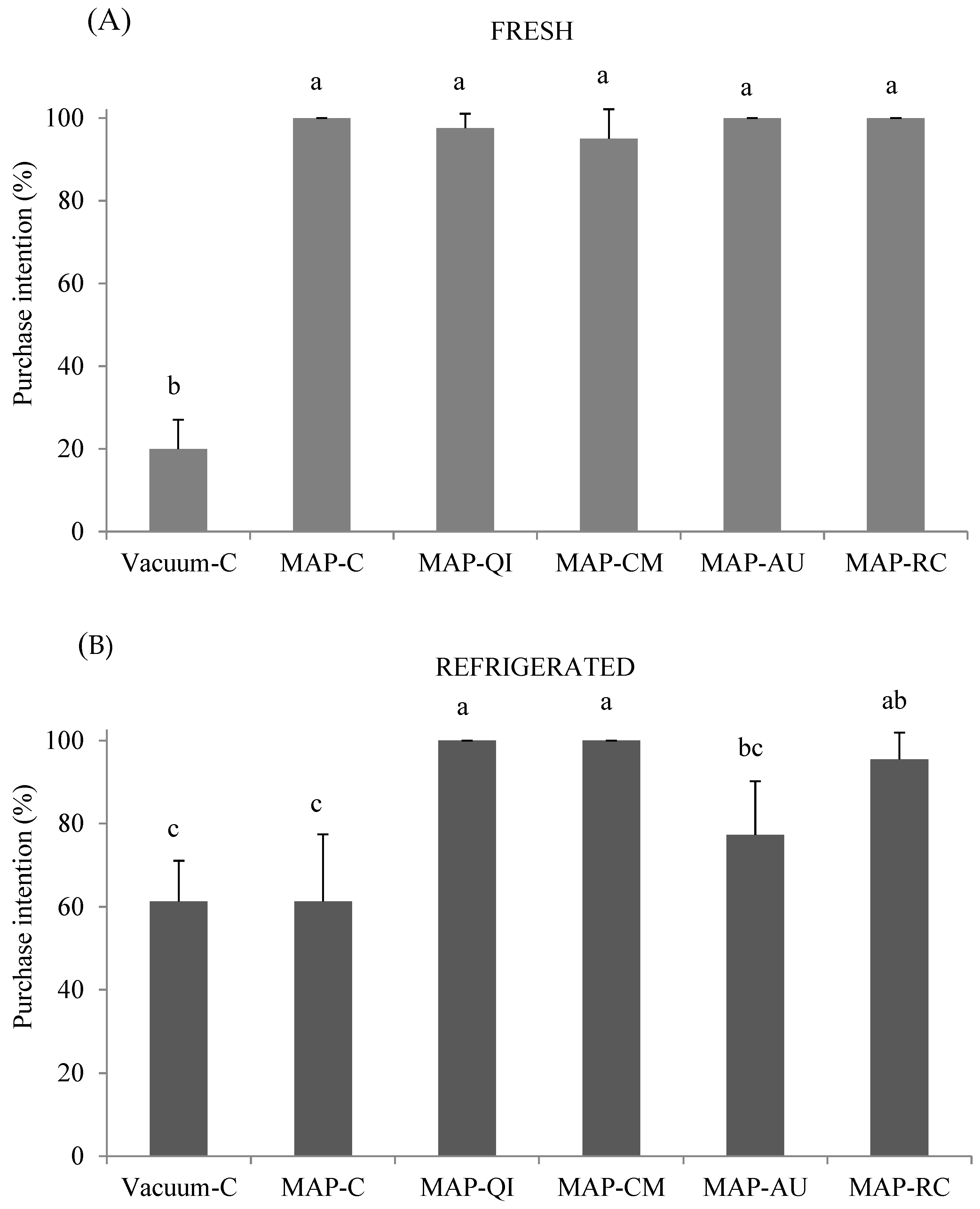
2.6.6. Purchase Intention
2.6.7. Statistical Analysis
3. Results and Discussion
3.1. Characterization of Fruit Extracts
3.2. Fruit Extracts as Inhibitors of Lipid Oxidation in Lamb Cutlets
3.3. Fruit Extracts as Inhibitors of Protein Oxidation in Lamb Cutlets
3.4. Impact of Fruit Extracts and Packaging on Consumers’ Purchase Intention
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Estévez, M.; Geraert, P.-A.; Liu, R.; Delgado, J.; Mercier, Y.; Zhang, W. Sulphur amino acids, muscle redox status and meat quality: More than building blocks—Invited review. Meat Sci. 2020, 163, 108087. [Google Scholar] [CrossRef]
- Soladoye, O.; Juárez, M.; Aalhus, J.; Shand, P.; Estévez, M. Protein Oxidation in Processed Meat: Mechanisms and Potential Implications on Human Health. Compr. Rev. Food Sci. Food Saf. 2015, 14, 106–122. [Google Scholar] [CrossRef]
- Frame, C.A.; Johnson, E.; Kilburn, L.; Huff-Lonergan, E.; Kerr, B.J.; Serao, M.C.R. Impact of dietary oxidized protein on oxidative status and performance in growing pigs. J. Anim. Sci. 2020, 98, 097. [Google Scholar] [CrossRef]
- Estévez, M.; Xiong, Y. Intake of Oxidized Proteins and Amino Acids and Causative Oxidative Stress and Disease: Recent Scientific Evidences and Hypotheses. J. Food Sci. 2019, 84, 387–396. [Google Scholar] [CrossRef]
- Estaras, M.; Ameur, F.Z.; Estévez, M.; Díaz-Velasco, S.; Gonzalez, A. The lysine derivative aminoadipic acid, a biomarker of protein oxidation and diabetes-risk, induces production of reactive oxygen species and impairs trypsin secretion in mouse pancreatic acinar cells. Food Chem. Toxicol. 2020, 145, 111594. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Velasco, S.; González, A.; Peña, F.; Estévez, M. Noxious effects of selected food-occurring oxidized amino acids on differentiated CACO-2 intestinal human cells. Food Chem. Toxicol. 2020, 144, 111650. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.N.; Lametsch, R.; Hviid, M.S.; Jensen, O.N.; Skibsted, L.H. High-oxygen packaging atmosphere influences protein oxidation and tenderness of porcine longissimus dorsi during chill storage. Meat Sci. 2007, 77, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Utrera, M.; Parra, V.; Estévez, M. Protein oxidation during frozen storage and subsequent processing of different beef muscles. Meat Sci. 2014, 96, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Li, S.; Warner, R.D.; Fang, Z. Effect of oregano essential oil and resveratrol nanoemulsion loaded pectin edible coating on the preservation of pork loin in modified atmosphere packaging. Food Control 2020, 114, 107226. [Google Scholar] [CrossRef]
- Moczkowska, M.; Półtorak, A.; Montowska, M.; Pospiech, E.; Wierzbicka, A. The effect of the packaging system and storage time on myofibrillar protein degradation and oxidation process in relation to beef tenderness. Meat Sci. 2017, 130, 7–15. [Google Scholar] [CrossRef]
- Peng, Y.; Adhiputra, K.; Padayachee, A.; Channon, H.A.; Ha, M.; Warner, R.D. High Oxygen Modified Atmosphere Packaging Negatively Influences Consumer Acceptability Traits of Pork. Foods 2019, 8, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, H.; Li, X.; Zhang, C.-H. Effects of oxygen concentration in modified atmosphere packaging on water holding capacity of pork steaks. Meat Sci. 2019, 148, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Polkinghorne, R.; Philpott, J.; Perovic, J.; Lau, J.; Davies, L.; Mudannayake, W.; Watson, R.; Tarr, G.; Thompson, J. The effect of packaging on consumer eating quality of beef. Meat Sci. 2018, 142, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Estévez, M.; Lorenzo, J. Impact of Antioxidants on Oxidized Proteins and Lipids in Processed Meat. Encycl. Food Chem. 2019, 600–608. [Google Scholar] [CrossRef]
- Kalogianni, A.I.; Lazou, T.; Bossis, I.; Gelasakis, A. Natural Phenolic Compounds for the Control of Oxidation, Bacterial Spoilage, and Foodborne Pathogens in Meat. Foods 2020, 9, 794. [Google Scholar] [CrossRef]
- Arcanjo, N.; Morcuende, D.; Andrade, M.; Padilla, P.; Madruga, M.; Estévez, M. Bioactivities of wine components on marinated beef during aging. J. Funct. Foods 2019, 57, 19–30. [Google Scholar] [CrossRef]
- Lahmar, A.; Morcuende, D.; Andrade, M.-J.; Chekir-Ghedira, L.; Estévez, M. Prolonging shelf life of lamb cutlets packed under high-oxygen modified atmosphere by spraying essential oils from North-African plants. Meat Sci. 2018, 139, 56–64. [Google Scholar] [CrossRef]
- Armenteros, M.; Morcuende, D.; Ventanas, J.; Estévez, M. The application of natural antioxidants via brine injection protects Iberian cooked hams against lipid and protein oxidation. Meat Sci. 2016, 116, 253–259. [Google Scholar] [CrossRef]
- Akcan, T.; Estévez, M.; Serdaroğlu, M. Antioxidant protection of cooked meatballs during frozen storage by whey protein edible films with phytochemicals from Laurus nobilis L. and Salvia officinalis. LWT 2017, 77, 323–331. [Google Scholar] [CrossRef]
- Andrade, M.A.; Ribeiro-Santos, R.; Guerra, M.; Silva, A.S. Evaluation of the Oxidative Status of Salami Packaged with an Active Whey Protein Film. Foods 2019, 8, 387. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, A.; Masuda, T. Chemical Interaction between Polyphenols and a Cysteinyl Thiol under Radical Oxidation Conditions. J. Agric. Food Chem. 2012, 60, 5142–5151. [Google Scholar] [CrossRef] [PubMed]
- Soong, Y.-Y.; Barlow, P.J. Antioxidant activity and phenolic content of selected fruit seeds. Food Chem. 2004, 88, 411–417. [Google Scholar] [CrossRef]
- Rodríguez-Carpena, J.; Morcuende, D.; Estévez, M. Avocado, sunflower and olive oils as replacers of pork back-fat in burger patties: Effect on lipid composition, oxidative stability and quality traits. Meat Sci. 2012, 90, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Özyürek, M.; Karademir, S.E. Novel Total Antioxidant Capacity Index for Dietary Polyphenols and Vitamins C and E, Using Their Cupric Ion Reducing Capability in the Presence of Neocuproine: CUPRAC Method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Utrera, M.; Estévez, M. Impact of trolox, quercetin, genistein and gallic acid on the oxidative damage to myofibrillar proteins: The carbonylation pathway. Food Chem. 2013, 141, 4000–4009. [Google Scholar] [CrossRef]
- Ganhão, R.; Estévez, M.; Morcuende, D. Suitability of the TBA method for assessing lipid oxidation in a meat system with added phenolic-rich materials. Food Chem. 2011, 126, 772–778. [Google Scholar] [CrossRef]
- Estévez, M.; Ventanas, S.; Ramírez, R.; Cava, R. Influence of the Addition of Rosemary Essential Oil on the Volatiles Pattern of Porcine Frankfurters. J. Agric. Food Chem. 2005, 53, 8317–8324. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Rodríguez-Carpena, J.-G.; Estévez, M. Fluorescent HPLC for the detection of specific protein oxidation carbonyls—α-aminoadipic and γ-glutamic semialdehydes—In meat systems. Meat Sci. 2011, 89, 500–506. [Google Scholar] [CrossRef]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.-P.; Pihlaja, K.; Kujala, A.T.S.; Heinonen, M. Antioxidant Activity of Plant Extracts Containing Phenolic Compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef]
- Ganhão, R.; Estévez, M.; Kylli, P.; Heinonen, M.; Morcuende, D. Characterization of Selected Wild Mediterranean Fruits and Comparative Efficacy as Inhibitors of Oxidative Reactions in Emulsified Raw Pork Burger Patties. J. Agric. Food Chem. 2010, 58, 8854–8861. [Google Scholar] [CrossRef]
- Cantos, E.; Espín, J.C.; López-Bote, C.; De La Hoz, L.; Ordóñez, J.A.; Tomás-Barberán, F.A. Phenolic Compounds and Fatty Acids from Acorns (Quercus spp.), the Main Dietary Constituent of Free-Ranged Iberian Pigs. J. Agric. Food Chem. 2003, 51, 6248–6255. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.C.S.; Morcuende, D.; Hérnandez-López, S.H.; Madruga, M.; Da Silva, F.A.P.; Estévez, M. Antioxidant Extracts from Acorns (Quercus ilex L.) Effectively Protect Ready-to-Eat (RTE) Chicken Patties Irrespective of Packaging Atmosphere. J. Food Sci. 2017, 82, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Akcan, T.; Gökçe, R.; Asensio, M.; Estévez, M.; Morcuende, D. Acorn (Quercus spp.) as a novel source of oleic acid and tocopherols for livestock and humans: Discrimination of selected species from Mediterranean forest. J. Food Sci. Technol. 2017, 54, 3050–3057. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Yamaguchi, T.; Takamura, H.; Atoba, T.M. Effects of Ascorbic Acid and α-Tocopherol on Antioxidant Activity of Polyphenolic Compounds. J. Food Sci. 2003, 68, 1622–1625. [Google Scholar] [CrossRef]
- Zhang, Q.; Cheng, Z.; Wang, Y.; Fu, L. Dietary protein-phenolic interactions: Characterization, biochemical-physiological consequences, and potential food applications. Crit. Rev. Food Sci. Nutr. 2020, 1–27. [Google Scholar] [CrossRef]
- Ganhão, R.; Morcuende, D.; Estévez, M. Tryptophan Depletion and Formation of α-Aminoadipic and γ-Glutamic Semialdehydes in Porcine Burger Patties with Added Phenolic-Rich Fruit Extracts. J. Agric. Food Chem. 2010, 58, 3541–3548. [Google Scholar] [CrossRef]
- Estévez, M.; Padilla, P.; Carvalho, L.; Martín, L.; Carrapiso, A.; Delgado, J. Malondialdehyde interferes with the formation and detection of primary carbonyls in oxidized proteins. Redox Biol. 2019, 26, 101277. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Ganhão, R.; Estévez, M. Role of Phenolics Extracting from Rosa canina L. on Meat Protein Oxidation during Frozen Storage and Beef Patties Processing. Food Bioprocess Technol. 2015, 8, 854–864. [Google Scholar] [CrossRef]
- Bermúdez, R.; Franco, D.; Carballo, J.; Lorenzo, J.M. Physicochemical changes during manufacture and final sensory characteristics of dry-cured Celta ham. Effect of muscle type. Food Control 2014, 43, 263–269. [Google Scholar] [CrossRef]
- Jongberg, S.; Wen, J.; Tørngren, M.A.; Lund, M.N. Effect of high-oxygen atmosphere packaging on oxidative stability and sensory quality of two chicken muscles during chill storage. Food Packag. Shelf Life 2014, 1, 38–48. [Google Scholar] [CrossRef]
- Lu, X.; Cornforth, D.P.; Carpenter, C.E.; Zhu, L.; Luo, X. Effect of oxygen concentration in modified atmosphere packaging on color changes of the M. longissimus thoraces et lumborum from dark cutting beef carcasses. Meat Sci. 2020, 161, 107999. [Google Scholar] [CrossRef] [PubMed]
- Vossen, E.; Utrera, M.; De Smet, S.; Morcuende, D.; Estévez, M. Dog rose (Rosa canina L.) as a functional ingredient in porcine frankfurters without added sodium ascorbate and sodium nitrite. Meat Sci. 2012, 92, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Akcan, T.; Estévez, M.; Rico, S.; Ventanas, S.; Morcuende, D. Hawberry (Crataegus monogyna Jaqc.) extracts inhibit lipid oxidation and improve consumer liking of ready-to-eat (RTE) pork patties. J. Food Sci. Technol. 2017, 54, 1248–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estévez, M.; Li, Z.; Soladoye, O.P.; Van-Hecke, T.; Soladoye, O. Health Risks of Food Oxidation. Adv. Food Nutr. Res. 2017, 82, 45–81. [Google Scholar] [CrossRef]
- Fu, Q.; Ge, Q.; Wang, H.; Zhang, W.; Liu, R.; Zhou, G. Influence of modified atmosphere packaging on protein oxidation, calpain activation and desmin degradation of beef muscles. J. Sci. Food Agric. 2017, 97, 4508–4514. [Google Scholar] [CrossRef]
- Lara, M.; Gutierrez, J.; Timón, M.; Andrés, A. Evaluation of two natural extracts (Rosmarinus officinalis L. and Melissa officinalis L.) as antioxidants in cooked pork patties packed in MAP. Meat Sci. 2011, 88, 481–488. [Google Scholar] [CrossRef]
- Jongberg, S.; Skov, S.H.; Tørngren, M.A.; Skibsted, L.H.; Lund, M.N. Effect of white grape extract and modified atmosphere packaging on lipid and protein oxidation in chill stored beef patties. Food Chem. 2011, 128, 276–283. [Google Scholar] [CrossRef]
- Nieto, G.; Jongberg, S.; Andersen, M.L.; Skibsted, L.H.; Nieto, G. Thiol oxidation and protein cross-link formation during chill storage of pork patties added essential oil of oregano, rosemary, or garlic. Meat Sci. 2013, 95, 177–184. [Google Scholar] [CrossRef]
- Viljanen, K.; Kylli, P.; Kivikari, R.; Heinonen, M. Inhibition of Protein and Lipid Oxidation in Liposomes by Berry Phenolics. J. Agric. Food Chem. 2004, 52, 7419–7424. [Google Scholar] [CrossRef]
- Luna, C.; Estévez, M. Formation of allysine in β-lactoglobulin and myofibrillar proteins by glyoxal and methylglyoxal: Impact on water-holding capacity and in vitro digestibility. Food Chem. 2019, 271, 87–93. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QI | CM | AU | RC | pA | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TFC B | 2055c | ± | 356 | 4362a | ± | 113 | 3480b | ± | 33 | 3117b | ± | 236 | 0.000 |
| Tocopherol C | 0.58c | ± | 0.09 | 2.09a | ± | 0.36 | 1.51b | ± | 0.05 | 1.25b | ± | 0.11 | 0.000 |
| Ascorbic acid D | 0.05c | ± | 0.05 | 0.12c | ± | 0.07 | 6.47a | ± | 0.48 | 4.79b | ± | 0.06 | 0.000 |
| Phenolic profile C | |||||||||||||
| HBA | 41.8a | ± | 4.9 | 0.8b | ± | 0.4 | 40.9a | ± | 3.8 | N.D | 0.000 | ||
| HCA | N.D | 38.9a | ± | 8.1 | N.D | 19.1b | ± | 1.7 | 0.000 | ||||
| FV | 1.50c | ± | 0.34 | 36.26a | ± | 0.84 | 1.00c | ± | 0.21 | 9.03b | ± | 1.08 | 0.000 |
| AC | N.D | 1.39b | ± | 0.24 | 2.33a | ± | 0.20 | 0.77c | ± | 0.10 | 0.000 | ||
| ET | 317 | ± | 31 | N.D | N.D | N.D | 0.000 | ||||||
| PC | 904c | ± | 45 | 1498b | ± | 126 | 315d | ± | 39 | 2322a | ± | 140 | 0.000 |
| Total | 1264c | ± | 133 | 1575b | ± | 168 | 359d | ± | 38 | 2350a | ± | 36 | 0.000 |
| Antioxidant activity E | |||||||||||||
| DPPH | 145ab | ± | 6 | 232a | ± | 48 | 218a | ± | 64 | 90b | ± | 3 | 0.008 |
| ABTS | 198 | ± | 25 | 153 | ± | 12 | 204 | ± | 3 | 201 | ± | 28 | 0.036 |
| CUPRAC | 354b | ± | 10 | 385ab | ± | 20 | 434a | ± | 23 | 431a | ± | 39 | 0.012 |
| % Inhibition POX F | 67.5a | ± | 0.3 | 31.4b | ± | 3.3 | 68.6a | ± | 0.4 | 73.6a | ± | 7.6 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morcuende, D.; Vallejo-Torres, C.; Ventanas, S.; Martínez, S.L.; Ruiz, S.C.; Estévez, M. Effectiveness of Sprayed Bioactive Fruit Extracts in Counteracting Protein Oxidation in Lamb Cutlets Subjected to a High-Oxygen MAP. Foods 2020, 9, 1715. https://doi.org/10.3390/foods9111715
Morcuende D, Vallejo-Torres C, Ventanas S, Martínez SL, Ruiz SC, Estévez M. Effectiveness of Sprayed Bioactive Fruit Extracts in Counteracting Protein Oxidation in Lamb Cutlets Subjected to a High-Oxygen MAP. Foods. 2020; 9(11):1715. https://doi.org/10.3390/foods9111715
Chicago/Turabian StyleMorcuende, D., C. Vallejo-Torres, S. Ventanas, S. L. Martínez, S. C. Ruiz, and M. Estévez. 2020. "Effectiveness of Sprayed Bioactive Fruit Extracts in Counteracting Protein Oxidation in Lamb Cutlets Subjected to a High-Oxygen MAP" Foods 9, no. 11: 1715. https://doi.org/10.3390/foods9111715