Nutritional and Functional Properties of Fermented Mixed Grains by Solid-State Fermentation with Bacillus amyloliquefaciens 245

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Solid-State Fermentation (SSF)
2.3. Metabolic Profiling Using Capillary Electrophoresis Time of Flight Mass Spectrometry (CE-TOF-MS) Analysis
2.4. Free Amino Acid Analysis
2.5. Quantitative Estimation of Targeted Arginine Metabolites Using HPLC
2.6. Determination of Enzymatic Activity
2.6.1. Sample Extraction
2.6.2. Amylase Activity Assay
2.6.3. Protease Activity Assay
2.6.4. Fibrinolytic Activity Assay
2.7. Antioxidant Activity Assay
2.7.1. Sample Extraction
2.7.2. Total Phenolic Content (TPC)
2.7.3. 2,2′-Azino-Bis (2-Ethyl Benzothiazoline-6-Sulphonate) (ABTS) Assay
3. Results and Discussion
3.1. Comparison of MG (Mixed Grains) Metabolites during Fermentation
3.2. The Contents of Free Amino Acids during the Fermentation of MG
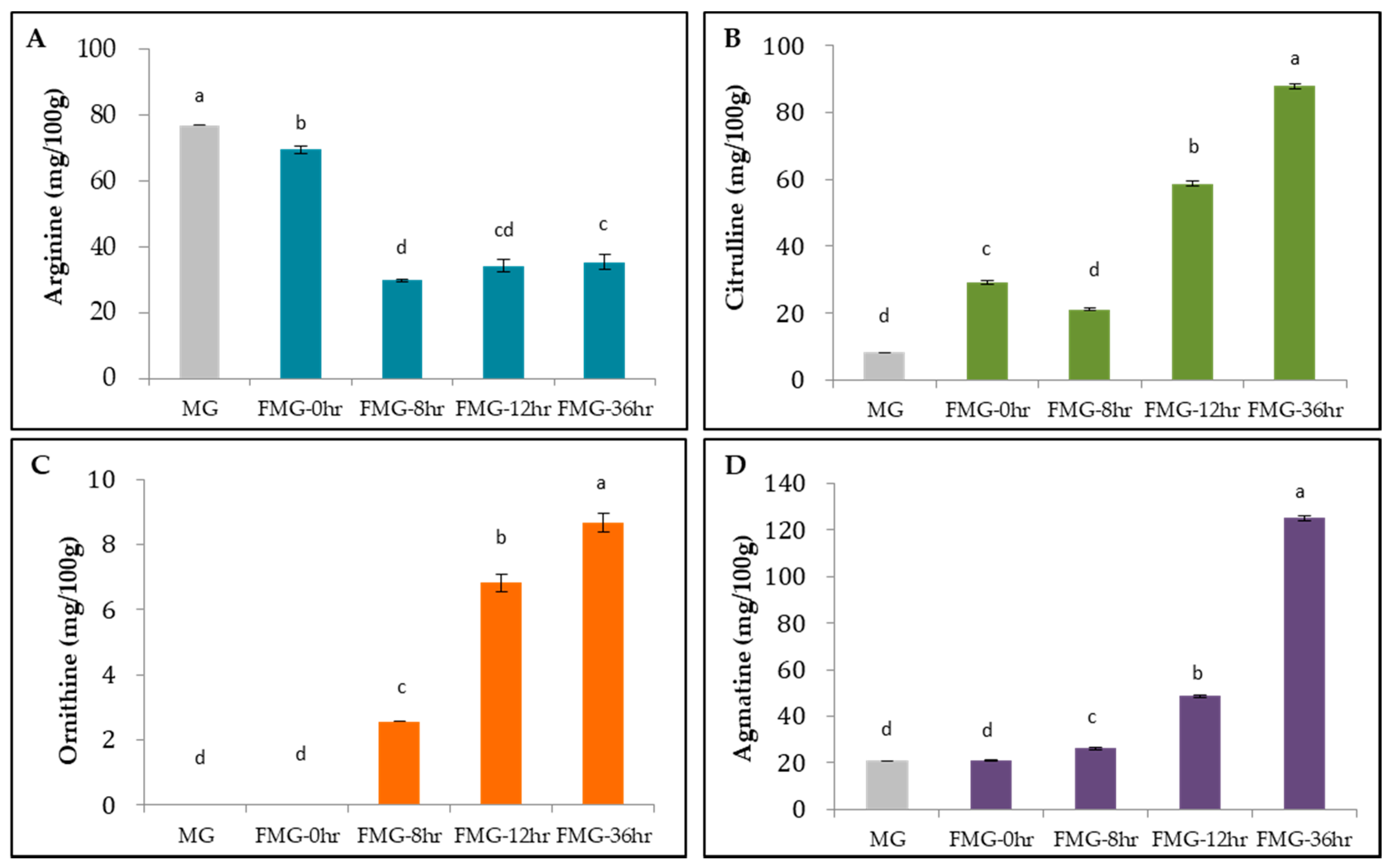
3.3. Content of Arginine Metabolites during Fermentation
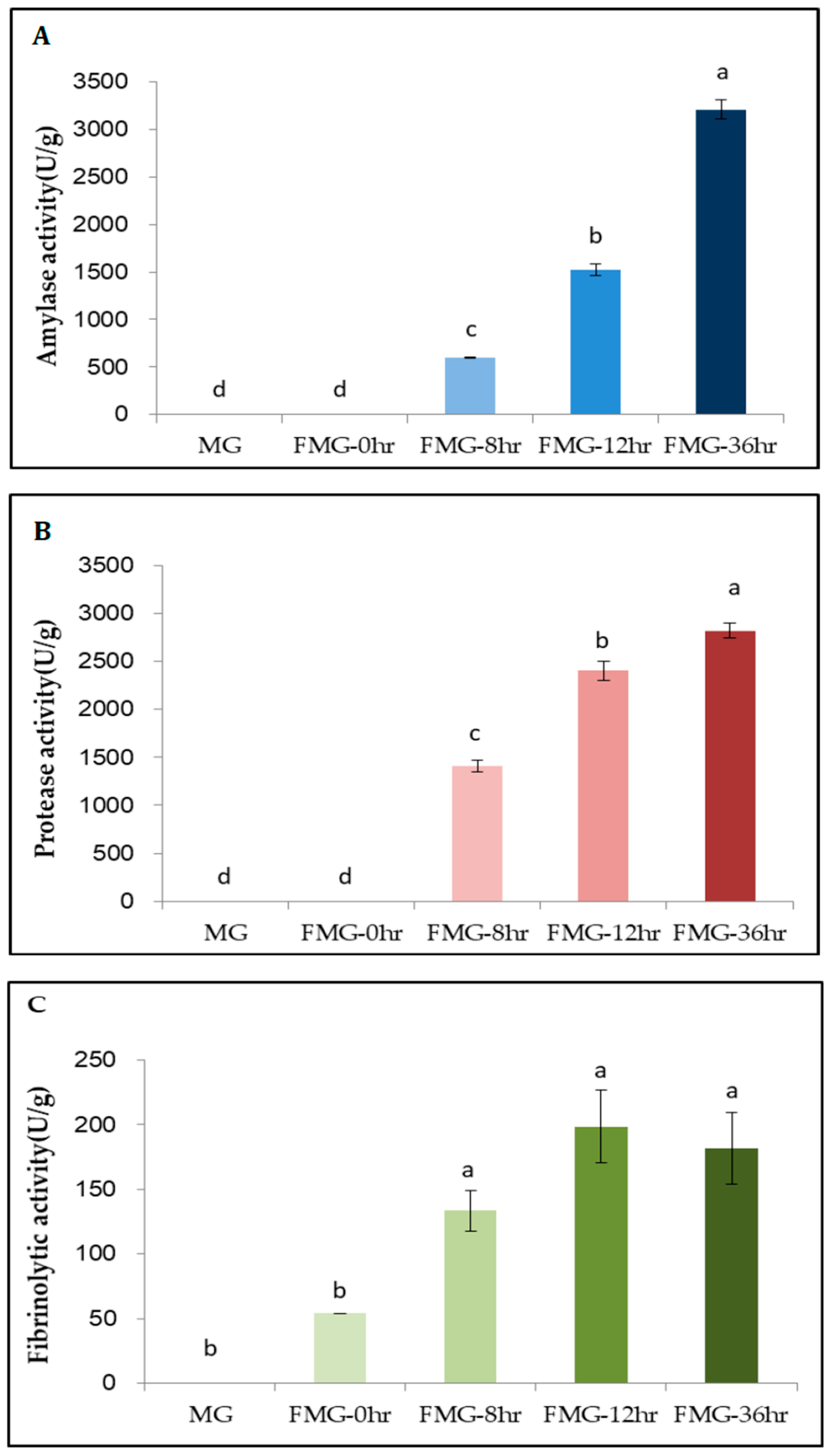
3.4. Enzymatic Activities in FMG
3.5. Total Phenolic Content (TPC) and ABTS+ Radical Scavenging Capacity in MG during Fermentation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aguilar Gorodecki, A. USDA: World Agricultural Supply and Demand Estimates; USDAGOV: Washington, DC, USA, 2012.
- Sefatie, R.S.; Fatoumata, T.; Eric, K.; Shi, Y.H.; Guo-Wei, L. In Vitro Antioxidant Activities of Protein Hydrolysate from Germinated Black Soybean (Glycine Max L.). Adv. J. Food Sci. Technol. 2013, 5, 453–459. [Google Scholar] [CrossRef]
- Brandolini, A.; Hidalgo, A. Wheat Germ: Not Only a by-Product. Int. J. Food Sci. Nutr. 2012, 63, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Coda, R.; Kärki, I.; Nordlund, E.; Heiniö, R.-L.; Poutanen, K.; Katina, K. Influence of Particle Size on Bioprocess Induced Changes on Technological Functionality of Wheat Bran. Food Microbiol. 2014, 37, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; Di Cagno, R.; Gobbetti, M. Use of Sourdough Fermented Wheat Germ for Enhancing the Nutritional, Texture and Sensory Characteristics of the White Bread. Eur. Food Res. Technol. 2010, 230, 645–654. [Google Scholar] [CrossRef]
- Castro-Alba, V.; Lazarte, C.E.; Perez-Rea, D.; Sandberg, A.-S.; Carlsson, N.-G.; Almgren, A.; Bergenståhl, B.; Granfeldt, Y. Effect of Fermentation and Dry Roasting on the Nutritional Quality and Sensory Attributes of Quinoa. Food Sci. Nutr. 2019, 7, 3902–3911. [Google Scholar] [CrossRef] [PubMed]
- Pepper, T.; Olinger, P.M. Xylitol in Sugar-Free Confections. Food Technol. Chic. 1988, 42, 98–106. [Google Scholar]
- Webber, D.M.; Hettiarachchy, N.S.; Li, R.; Horax, R.; Theivendran, S. Phenolic Profile and Antioxidant Activity of Extracts Prepared from Fermented Heat-Stabilized Defatted Rice Bran. J. Food Sci. 2014, 79, H2383–H2391. [Google Scholar] [CrossRef]
- Manan, M.A.; Webb, C. Design Aspects of Solid State Fermentation as Applied to Microbial Bioprocessing. J. Appl. Biotechnol. Bioeng. 2017, 4, 91. [Google Scholar]
- Chi, C.-H.; Cho, S.-J. Improvement of Bioactivity of Soybean Meal by Solid-State Fermentation with Bacillus Amyloliquefaciens Versus Lactobacillus Spp. and Saccharomyces Cerevisiae. LWT-Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Shahzad, R.; Shehzad, A.; Bilal, S.; Lee, I.-J. Bacillus Amyloliquefaciens RWL-1 as a New Potential Strain for Augmenting Biochemical and Nutritional Composition of Fermented Soybean. Molecules 2020, 25, 2346. [Google Scholar] [CrossRef]
- Bacillus Amyloliquefaciens Strain BA245 16S Ribosomal RNA Gene, Partial Sequence. Available online: https://www.ncbi.nlm.nih.gov/nuccore/kr535604 (accessed on 18 July 2015).
- Park, M.-J.; Kim, A.-J.; Han, S.W.; Heo, S.J.; Yang, T.J.; Park, S.W.; Lee, S.B.; Jang, J.H.; Cho, S.J.; Hong, Y.H.; et al. Novel Strain Derived from Traditional Fermented Food and Having Excellent Enzyme Productivity, and Method for Preparing Fermented Cereal Enzyme Food by Using Same. U.S. Patent WO2017074037A1, 4 May 2017. [Google Scholar]
- Soga, T.; Ueno, Y.; Naraoka, H.; Ohashi, Y.; Tomita, M.; Nishioka, T. Simultaneous Determination of Anionic Intermediates for Bacillus subtilis Metabolic Pathways by Capillary Electrophoresis Electrospray Ionization Mass Spectrometry. Anal. Chem. 2002, 74, 2233–2239. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Ohashi, Y.; Ueno, Y.; Naraoka, H.; Tomita, M.; Nishioka, T. Quantitative Metabolome Analysis Using Capillary Electrophoresis Mass Spectrometry. J. Proteome Res. 2003, 2, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Henderson, J.W.; Brooks, A. Improved Amino Acid Methods Using Agilent ZORBAX Eclipse Plus C18 Columns for a Variety of Agilent LC Instrumentation and Separation Goals; Agilent Technologies: Santa Clara, CA, USA, 2010. [Google Scholar]
- Caro, G.M.; Meierhenrich, U.J.; Schutte, W.A.; Barbier, B.; Segovia, A.A.; Rosenbauer, H.; Thiemann, W.-P.; Brack, A.; Greenberg, J.M. Amino Acids from Ultraviolet Irradiation of Interstellar Ice Analogues. Nature 2002, 416, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Pfueller, S.L.; Elliott, W.H. The Extracellular α-Amylase of Bacillus Stearothermophilus. J. Biol. Chem. 1969, 244, 48–54. [Google Scholar]
- Kum, S.-J.; Yang, S.-O.; Lee, S.M.; Chang, P.-S.; Choi, Y.H.; Lee, J.J.; Hurh, B.S.; Kim, Y.-S. Effects of Aspergillus Species Inoculation and Their Enzymatic Activities on the Formation of Volatile Components in Fermented Soybean Paste (Doenjang). J. Agric. Food Chem. 2015, 63, 1401–1418. [Google Scholar] [CrossRef]
- Chang, C.-T.; Fan, M.-H.; Kuo, F.-C.; Sung, H.-Y. Potent Fibrinolytic Enzyme from a Mutant of Bacillus s Ubtilis IMR-NK1. J. Agric. Food Chem. 2000, 48, 3210–3216. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Puchalska, P.; Marina, M.L.; García, M.C. Isolation and Identification of Antioxidant Peptides from Commercial Soybean-Based Infant Formulas. Food Chem. 2014, 148, 147–154. [Google Scholar] [CrossRef]
- Hamad, A.M.; Fields, M.L. Evaluation of the Protein Quality and Available Lysine of Germinated and Fermented Cereals. J. Food Sci. 1979, 44, 456–459. [Google Scholar] [CrossRef]
- Kang, H.J.; Yang, H.J.; Kim, M.J.; Han, E.-S.; Kim, H.-J.; Kwon, D.Y. Metabolomic Analysis of Meju during Fermentation by Ultra Performance Liquid Chromatography-Quadrupole-Time of Flight Mass Spectrometry (UPLC-Q-TOF MS). Food Chem. 2011, 127, 1056–1064. [Google Scholar] [CrossRef]
- Akram, M.; Daniyal, M.; Ali, A.; Zainab, R.; Shah, S.M.A.; Munir, N.; Tahir, I.M. Role of Phenylalanine and Its Metabolites in Health and Neurological Disorders. In Synucleins-Biochemistry and Role in Diseases; IntechOpen: London, UK, 2020. [Google Scholar]
- Jansen, G.R. Lysine in Human Nutrition. J. Nutr. 1962, 76, 1–35. [Google Scholar] [CrossRef]
- Chen, Y.-M.; Lin, C.-L.; Wei, L.; Hsu, Y.-J.; Chen, K.-N.; Huang, C.-C.; Kao, C.-H. Sake Protein Supplementation Affects Exercise Performance and Biochemical Profiles in Power-Exercise-Trained Mice. Nutrients 2016, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Fang, F.; Zhou, X.; Chen, X.; Zhang, L.; Du, G.; Chen, J. Characterization of Arginine Utilization Strains from Fermented Grains and Evaluation of Their Contribution to Citrulline Accumulation in Chinese Luzhou-Flavor Spirits. Acta Microbiol. Sin. 2016, 56, 1638–1646. [Google Scholar]
- Hwang, H.; Lee, J.-H. Characterization of Arginine Catabolism by Lactic Acid Bacteria Isolated from Kimchi. Molecules 2018, 23, 3049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akasaka, N.; Fujiwara, S. The Therapeutic and Nutraceutical Potential of Agmatine, and Its Enhanced Production Using Aspergillus Oryzae. Amino Acids 2020, 52, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.M.; Gills, J.; Gray, M. Acute Effects of Nitrosigine® and Citrulline Maleate on Vasodilation. J. Int. Soc. Sports Nutr. 2020, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, T.; Maeda, M.; Hayashi, T. Administration of L-Arginine plus L-Citrulline or L-Citrulline Alone Successfully Retarded Endothelial Senescence. PLoS ONE 2018, 13, e0192252. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, U.; Didelija, I.C.; Yuan, Y.; Wang, X.; Marini, J.C. Supplemental Citrulline Is More Efficient than Arginine in Increasing Systemic Arginine Availability in Mice. J. Nutr. 2017, 147, 596–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugino, T.; Shirai, T.; Kajimoto, Y.; Kajimoto, O. L-Ornithine Supplementation Attenuates Physical Fatigue in Healthy Volunteers by Modulating Lipid and Amino Acid Metabolism. Nutr. Res. 2008, 28, 738–743. [Google Scholar] [CrossRef]
- Raghavan, S.A.; Dikshit, M. Vascular Regulation by the L-Arginine Metabolites, Nitric Oxide and Agmatine. Pharmacol. Res. 2004, 49, 397–414. [Google Scholar] [CrossRef]
- Li, P.; Liang, H.; Lin, W.-T.; Feng, F.; Luo, L. Microbiota Dynamics Associated with Environmental Conditions and Potential Roles of Cellulolytic Communities in Traditional Chinese Cereal Starter Solid-State Fermentation. Appl. Environ. Microbiol. 2015, 81, 5144–5156. [Google Scholar] [CrossRef] [Green Version]
- Kunamneni, A.; Permaul, K.; Singh, S. Amylase Production in Solid State Fermentation by the Thermophilic Fungus Thermomyces Lanuginosus. J. Biosci. Bioeng. 2005, 100, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Son, S.-J.; Lee, S.-P. Evaluation of Rheological and Functional Properties of Roasted Soybean Flour and Mixed Cereals Fermented by Bacillus Sp. J. Korean Soc. Food Sci. Nutr. 2011, 40, 450–457. [Google Scholar] [CrossRef]
- Vijayaraghavan, P.; Prakash Vincent, S.G. Statistical Optimization of Fibrinolytic Enzyme Production Using Agroresidues by Bacillus Cereus IND1 and Its Thrombolytic Activity in Vitro. BioMed Res. Int. 2014, 2014, 725064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, C.-H.; Shen, M.-C.; Lin, J.-S.; Wen, Y.-K.; Hwang, K.-L.; Cham, T.-M.; Yang, N.-C. Nattokinase Decreases Plasma Levels of Fibrinogen, Factor VII, and Factor VIII in Human Subjects. Nutr. Res. 2009, 29, 190–196. [Google Scholar] [CrossRef]
- Choi, H.-K.; Yoon, J.-H.; Kim, Y.-S.; Kwon, D.Y. Metabolomic Profiling of Cheonggukjang during Fermentation by 1H NMR Spectrometry and Principal Components Analysis. Process Biochem. 2007, 42, 263–266. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant Activity and Total Phenolics in Selected Fruits, Vegetables, and Grain Products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Cao, X.; Yang, L.; Xue, Q.; Yao, F.; Sun, J.; Yang, F.; Liu, Y. Antioxidant Evaluation-Guided Chemical Profiling and Structure-Activity Analysis of Leaf Extracts from Five Trees in Broussonetia and Morus (Moraceae). Sci. Rep. 2020, 10, 4808. [Google Scholar] [CrossRef]
- Dash, P.; Ghosh, G. Amino Acid Composition, Antioxidant and Functional Properties of Protein Hydrolysates from Cucurbitaceae Seeds. J. Food Sci. Technol. 2017, 54, 4162–4172. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids | MG | FMG-0hr | FMG-8hr | FMG-12hr | FMG-36hr |
|---|---|---|---|---|---|
| (mg/100 g) | |||||
| Essential Amino Acids | |||||
| Histidine | 13.27 ± 0.31 c | ND | 13.88 ± 0.46 c | 26.47 ± 1.03 b | 60.37 ± 1.83 a |
| Isoleucine | 7.43 ± 0.21 d | 6.40 ± 0.72 d | 9.86 ± 1.51 c | 31.30 ± 1.01 b | 66.52 ± 1.8 a |
| Leucine | 19.74 ± 0.65 d | 14.17 ± 0.23 e | 33.44 ± 0.88 c | 70.39 ± 2.12 b | 139.21 ± 1.25 a |
| Lysine | 19.46 ± 1.05 d | 19.46 ± 0.11 d | 23.7 ± 0.89 c | 61.64 ± 1.74 b | 184.25 ± 1.12 a |
| Methionine | 16.19 ± 0.17 b | ND | ND | 13.45 ± 0.52 c | 24.21 ± 2.33 a |
| Phenylalanine | 12.65 ± 0.67 d | 9.34 ± 0.20 e | 39.86 ± 1.41 c | 81.46 ± 2.61 b | 239.46 ± 1.5 a |
| Threonine | 10.71 ± 0.73 b | 8.51 ± 0.15 b | 9.35 ± 1.65 b | 22.50 ± 1.18 a | 23.38 ± 2.25 a |
| Tryptophan | 13.92 ± 0.56 d | 11.40 ± 0.20 e | 34.61 ± 1.04 c | 78.95 ± 1.89 b | 178.23 ± 1.68 a |
| Valine | 29.00 ± 0.50 d | 32.00 ± 2.65 cd | 33.83 ± 0.76 c | 43.00 ± 2.00 b | 91.67 ± 1.53 a |
| Non-Essential Amino Acids | |||||
| Alanine | 36.49 ± 0.38 c | 26.47 ± 0.42 d | 23.36 ± 0.75 e | 91.32 ± 1.63 b | 113.88 ± 1.84 a |
| Arginine | 76.90 ± 1.30 a | 69.46 ± 0.29 b | 29.78 ± 1.88 d | 34.16 ± 2.32 c | 35.29 ± 1.69 c |
| Aspartic acid | 13.48 ± 0.80 d | 11.14 ± 0.09 e | 20.48 ± 1.04 c | 22.81 ± 1.30 b | 41.28 ± 2.13 a |
| Glutamic acid | 37.07 ± 1.37 d | 32.38 ± 0.77 e | 149.96 ± 1.89 c | 227.08 ± 2.52 a | 175.07 ± 0.9 b |
| Glycine | 23.61 ± 0.29 a | 15.29 ± 0.68 b | 6.78 ± 0.2 d | 15.26 ± 0.20 b | 14.30 ± 0.33 c |
| Cysteine | ND | ND | ND | ND | ND |
| Proline | 8.23 ± 0.21 e | 12.33 ± 1.53 d | 18.33 ± 1.53 c | 46.33 ± 3.21 b | 79.00 ± 1.00 a |
| Serine | 11.22 ± 0.86 b | 7.96 ± 0.55 c | ND | 10.73 ± 0.45 b | 20.21 ± 2.56 a |
| Tyrosine | 12.02 ± 0.53 d | ND | 18.56 ± 0.76 c | 44.69 ± 1.55 b | 121.38 ± 1.10 a |
| Total | 368.76 | 287.35 | 457.59 | 925.26 | 1615.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heo, S.J.; Kim, A.-J.; Park, M.-J.; Kang, K.; Soung, D.Y. Nutritional and Functional Properties of Fermented Mixed Grains by Solid-State Fermentation with Bacillus amyloliquefaciens 245. Foods 2020, 9, 1693. https://doi.org/10.3390/foods9111693
Heo SJ, Kim A-J, Park M-J, Kang K, Soung DY. Nutritional and Functional Properties of Fermented Mixed Grains by Solid-State Fermentation with Bacillus amyloliquefaciens 245. Foods. 2020; 9(11):1693. https://doi.org/10.3390/foods9111693
Chicago/Turabian StyleHeo, Su Jin, Ah-Jin Kim, Min-Ju Park, Kimoon Kang, and Do Yu Soung. 2020. "Nutritional and Functional Properties of Fermented Mixed Grains by Solid-State Fermentation with Bacillus amyloliquefaciens 245" Foods 9, no. 11: 1693. https://doi.org/10.3390/foods9111693