Digestomics of Cow’s Milk: Short Digestion-Resistant Peptides of Casein Form Functional Complexes by Aggregation

 ,
,  , , , and
, , , and
Abstract
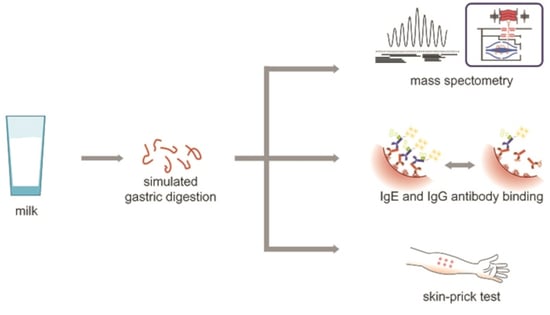
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Digestion of Raw Milk in Simulated Conditions of the Stomach
2.2. Detection of α-Lactalbumin (ALA) and β-Lactoglobulin (BLG) by Immunoblotting
2.3. Preparation of Milk SDRPs
2.4. Mass Spectrometry Analysis
2.5. Patients
2.6. Hydrophobicity Calculations
2.7. Size-Exclusion Chromatography (SEC)
2.8. Electrophoretic Analysis of SDRPs
2.9. IgE-Binding Properties of SDRPs
2.10. IgG4-Binding Properties of SDRPs
2.11. Skin-Prick Tests (SPTs)
2.12. Statistical Analysis
3. Results
3.1. Raw Milk Pepsin Digestion Reveals Intact Proteins
3.2. Peptide Analysis by High-Resolution Mass Spectrometry
3.3. SDRPs Originating from Milk Allergens Aggregate into Higher Molecular Weight Complexes
3.4. SDRPs Associate into Functional Complexes, Bind IgE, and Do Not Contribute to the Tolerance to Milk Allergens
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sampson, H.A. Food allergy. Part 1: Immunopathogenesis and clinical disorders. J. Allergy Clin. Immunol. 1999, 103, 717–728. [Google Scholar] [CrossRef]
- Kattan, J.D.; Cocco, R.R.; Järvinen, K.M. Milk and Soy Allergy. Pediatr. Clin. N. Am. 2011, 58, 407–426. [Google Scholar] [CrossRef]
- Fiocchi, A.; Brozek, J.; Schuenemann, H.; Bahna, S.L.; Von Berg, A.; Beyer, K.; Bozzola, M.; Bradsher, J.; Compalati, E.; Ebisawa, M.; et al. World Allergy Organization (WAO) Diagnosis and Rationale for Action against Cow’s Milk Allergy (DRACMA) Guidelines. Pediatr. Allergy Immunol. 2010, 21, 1–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, R.; Kuslys, M.; Muraro, A.; Høst, A. Controversies on Special Products for Managing Cow’s Milk Protein Allergy in Infants: Safety and Suitability. EMJ Allergy Immunol. 2017, 2, 46–51. [Google Scholar]
- American Academy of Pediatrics. Committee on Nutrition. Hypoallergenic infant formulas. Pediatrics 2000, 106, 346–349. [Google Scholar] [CrossRef] [Green Version]
- Astwood, J.D.; Leach, J.N.; Fuchs, R.L. Stability of food allergens to digestion in vitro. Nat. Biotechnol. 1996, 14, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.-J.; Abbott, U.R.; Hatzos, C. Digestibility of Food Allergens and Nonallergenic Proteins in Simulated Gastric Fluid and Simulated Intestinal FluidA Comparative Study. J. Agric. Food Chem. 2002, 50, 7154–7160. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations (FAO). Evaluation of Allergenicity of Genetically Modified Foods: Report of a Joint FAO/WHO Expert Consultation on Allergenicity of Foods Derived from Biotechnology; FAO: Rome, Italy, 2001. [Google Scholar]
- Thomas, K.; Aalbers, M.; Bannon, G.A.; Bartels, M.; Dearman, R.J.; Esdaile, D.J.; Fu, T.J.; Glatt, C.M.; Hadfield, N.; Hatzos, C.; et al. A multi-laboratory evaluation of a common in vitro pepsin digestion assay protocol used in assessing the safety of novel proteins. Regul. Toxicol. Pharmacol. 2004, 39, 87–98. [Google Scholar] [CrossRef]
- Eiwegger, T.; Rigby, N.; Mondoulet, L.; Bernard, H.; Krauth, M.-T.; Boehm, A.; Dehlink, E.; Valent, P.; Wal, J.M.; Mills, E.N.C.; et al. Gastro-duodenal digestion products of the major peanut allergen Ara h 1 retain an allergenic potential. Clin. Exp. Allergy 2006, 36, 1281–1288. [Google Scholar] [CrossRef]
- Moreno, F.J.; Mackie, A.A.R.; Mills, E.N.C. Phospholipid Interactions Protect the Milk Allergen α-Lactalbumin from Proteolysis during in Vitro Digestion. J. Agric. Food Chem. 2005, 53, 9810–9816. [Google Scholar] [CrossRef]
- Bøgh, K.L.; Kroghsbo, S.; Dahl, L.; Rigby, N.M.; Barkholt, V.; Mills, E.N.C.; Madsen, C.B. Digested Ara h 1 has sensitizing capacity in Brown Norway rats. Clin. Exp. Allergy 2009, 39, 1611–1621. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Genetically Modified Organisms (GMO). Guidance for risk assessment of food and feed from genetically modified plants. EFSA J. 2011, 9, 2150–2187. [Google Scholar] [CrossRef]
- Benedé, S.; López-Expósito, I.; Gimenez, G.; Grishina, G.; Bardina, L.; Sampson, H.A.; Molina, E.; López-Fandiño, R. In vitro digestibility of bovine β-casein with simulated and human oral and gastrointestinal fluids. Identification and IgE-reactivity of the resultant peptides. Food Chem. 2014, 143, 514–521. [Google Scholar] [CrossRef]
- Toomer, O.T.; Do, A.; Pereira, M.; Williams, K. Effect of Simulated Gastric and Intestinal Digestion on Temporal Stability and Immunoreactivity of Peanut, Almond, and Pine Nut Protein Allergens. J. Agric. Food Chem. 2013, 61, 5903–5913. [Google Scholar] [CrossRef]
- Bouzerzour, K.; Morgan, F.; Cuinet, I.; Bonhomme, C.; Jardin, J.; Le Huërou-Luron, I.; Dupont, D. In vivo digestion of infant formula in piglets: Protein digestion kinetics and release of bioactive peptides. Br. J. Nutr. 2012, 108, 2105–2114. [Google Scholar] [CrossRef] [Green Version]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised staticin vitrodigestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Polovic, N.; Blanŭsa, M.; Gavrović-Jankulović, M.; Atanaskovic-Markovic, M.; Burazer, L.; Jankov, R.; Veličković, T. Ćirković A matrix effect in pectin-rich fruits hampers digestion of allergen by pepsin in vivo and in vitro. Clin. Exp. Allergy 2007, 37, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Prodic, I.; Stanić, D.; Apostolović, D.; Mihailović, J.M.; Radibratovic, M.; Radosavljevic, J.; Burazer, L.; Milčić, M.; Smiljanic, K.; Van Hage, M.; et al. Influence of peanut matrix on stability of allergens in gastric-simulated digesta: 2S albumins are main contributors to the IgE reactivity of short digestion-resistant peptides. Clin. Exp. Allergy 2018, 48, 731–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picariello, G.; Mamone, G.; Nitride, C.; Addeo, F.; Ferranti, P. Protein digestomics: Integrated platforms to study food-protein digestion and derived functional and active peptides. TrAC Trends Anal. Chem. 2013, 52, 120–134. [Google Scholar] [CrossRef]
- Van Beresteijn, E.C.H.; Peeters, R.A.; Kaper, J.; Meijer, R.J.G.M.; Robben, A.J.P.M.; Schmidt, D.G. Molecular Mass Distribution Immunological Properties Nutritive Value of Whey Protein Hydrolysates. J. Food Prot. 1994, 57, 619–625. [Google Scholar] [CrossRef]
- Siemensma, A.D.; Weijer, W.J.; Bak, H.J. The importance of peptide lengths in hypoallergenic infant formulae. Trends Food Sci. Technol. 1993, 4, 16–21. [Google Scholar] [CrossRef]
- Apostolovic, D.; Stanic-Vucinic, D.; De Jongh, H.H.J.; De Jong, G.A.H.; Mihailovic, J.; Radosavljevic, J.; Radibratovic, M.; Nordlee, J.A.; Baumert, J.L.; Milcic, M.; et al. Conformational stability of digestion-resistant peptides of peanut conglutins reveals the molecular basis of their allergenicity. Sci. Rep. 2016, 6, 29249. [Google Scholar] [CrossRef] [PubMed]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Busse, P.J.; Järvinen, K.-M.; Vila, L.; Beyer, K.; Sampson, H.A. Identification of Sequential IgE-Binding Epitopes on Bovine αs2-Casein in Cow’s Milk Allergic Patients. Int. Arch. Allergy Immunol. 2002, 129, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Chatchatee, P.; Järvinen, K.-M.; Bardina, L.; Beyer, K.; Sampson, H.A. Identification of IgE- and IgG-binding epitopes on αs1-casein: Differences in patients with persistent and transient cow’s milk allergy. J. Allergy Clin. Immunol. 2001, 107, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Chatchatee, P.; Jarvinen, K.M.; Bardina, L.; Vila, L.; Beyer, K.; A Sampson, H. Identification of IgE and IgG binding epitopes on β- and κ-casein in cow’s milk allergic patients. Clin. Exp. Allergy 2001, 31, 1256–1262. [Google Scholar] [CrossRef]
- Järvinen, K.-M.; Chatchatee, P.; Bardina, L.; Beyer, K.; Sampson, H.A. IgE and IgG Binding Epitopes on α-Lactalbumin and β-Lactoglobulin in Cow’s Milk Allergy. Int. Arch. Allergy Immunol. 2001, 126, 111–118. [Google Scholar] [CrossRef]
- Picariello, G.; Iacomino, G.; Mamone, G.; Ferranti, P.; Fierro, O.; Gianfrani, C.; Di Luccia, A.; Addeo, F. Transport across Caco-2 monolayers of peptides arising from in vitro digestion of bovine milk proteins. Food Chem. 2013, 139, 203–212. [Google Scholar] [CrossRef]
- Barbana, C.; Pérez, M.; Sánchez, L.; Dalgalarrondo, M.; Chobert, J.; Haertlé, T.; Calvo, M. Interaction of bovine -lactalbumin with fatty acids as determined by partition equilibrium and fluorescence spectroscopy. Int. Dairy J. 2006, 16, 18–25. [Google Scholar] [CrossRef]
- Al-Hanish, A.; Stanic-Vucinic, D.; Mihailovic, J.; Prodic, I.; Minić, S.; Stojadinovic, M.; Radibratovic, M.; Milcic, M.; Velickovic, T.C. Noncovalent interactions of bovine α-lactalbumin with green tea polyphenol, epigalocatechin-3-gallate. Food Hydrocoll. 2016, 61, 241–250. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Grishchenko, V.M.; Kalinichenko, L.P.; Orlov, N.Y.; Kuwajima, K.; Sugai, S. Calcium-regulated interactions of human α-lactalbumin with bee venom melittin. Biophys. Chem. 1991, 39, 111–117. [Google Scholar] [CrossRef]
- Wal, J.-M. Cow’s milk allergens. Allergy 1998, 53, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Egger, L.; Ménard, O.; Baumann, C.; Duerr, D.; Schlegel, P.; Stoll, P.; Vergères, G.; Dupont, D.; Portmann, R. Digestion of milk proteins: Comparing static and dynamic in vitro digestion systems with in vivo data. Food Res. Int. 2019, 118, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, M.; Kühne, Y.; Ballmer-Weber, B.K.; Becker, W.-M.; Holzhauser, T.; Lauer, I.; Reuter, A.; Randow, S.; Falk, S.; Wangorsch, A.; et al. Relevance of IgE binding to short peptides for the allergenic activity of food allergens. J. Allergy Clin. Immunol. 2009, 124, 328–336.e6. [Google Scholar] [CrossRef]
- Bøgh, K.L.; Barkholt, V.; Rigby, N.M.; Mills, E.N.C.; Madsen, C.B. Digested Ara h 1 Loses Sensitizing Capacity When Separated into Fractions. J. Agric. Food Chem. 2012, 60, 2934–2942. [Google Scholar] [CrossRef] [Green Version]
- Huby, R.D.J.; Dearman, R.J.; Kimber, I. Why Are Some Proteins Allergens? Toxicol. Sci. 2000, 55, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Luyt, D.; Ball, H.; Makwana, N.; Green, M.R.; Bravin, K.; Nasser, S.M.; Clark, A.T. BSACI guideline for the diagnosis and management of cow’s milk allergy. Clin. Exp. Allergy 2014, 44, 642–672. [Google Scholar] [CrossRef]
- Caubet, J.C.; Lin, J.; Ahrens, B.; Gimenez, G.; Bardina, L.; Niggemann, B.; Sampson, H.A.; Beyer, K. Natural tolerance development in cow’s milk allergic children: IgE and IgG4 epitope binding. Allergy 2017, 72, 1677–1685. [Google Scholar] [CrossRef]
- Savilahti, E.M.; Rantanen, V.; Lin, J.S.; Karinen, S.; Saarinen, K.M.; Goldis, M.; Mäkelä, M.J.; Hautaniemi, S.; Savilahti, E.; Sampson, H.A. Early recovery from cow’s milk allergy is associated with decreasing IgE and increasing IgG4 binding to cow’s milk epitopes. J. Allergy Clin. Immunol. 2010, 125, 1315–1321.e9. [Google Scholar] [CrossRef] [Green Version]
- Savilahti, E.M.; Kuitunen, M.; Valori, M.; Rantanen, V.; Bardina, L.; Gimenez, G.; Mäkelä, M.J.; Hautaniemi, S.; Savilahti, E.; Sampson, H.A. Use of IgE and IgG4 epitope binding to predict the outcome of oral immunotherapy in cow’s milk allergy. Pediatr. Allergy Immunol. 2014, 25, 227–235. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radosavljević, J.; Apostolović, D.; Mihailović, J.; Atanasković-Marković, M.; Burazer, L.; van Hage, M.; Ćirković Veličković, T. Digestomics of Cow’s Milk: Short Digestion-Resistant Peptides of Casein Form Functional Complexes by Aggregation. Foods 2020, 9, 1576. https://doi.org/10.3390/foods9111576
Radosavljević J, Apostolović D, Mihailović J, Atanasković-Marković M, Burazer L, van Hage M, Ćirković Veličković T. Digestomics of Cow’s Milk: Short Digestion-Resistant Peptides of Casein Form Functional Complexes by Aggregation. Foods. 2020; 9(11):1576. https://doi.org/10.3390/foods9111576
Chicago/Turabian StyleRadosavljević, Jelena, Danijela Apostolović, Jelena Mihailović, Marina Atanasković-Marković, Lidija Burazer, Marianne van Hage, and Tanja Ćirković Veličković. 2020. "Digestomics of Cow’s Milk: Short Digestion-Resistant Peptides of Casein Form Functional Complexes by Aggregation" Foods 9, no. 11: 1576. https://doi.org/10.3390/foods9111576