Quality Changes during Frozen Storage of Mechanical-Separated Flesh Obtained from an Underutilized Crustacean


 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Raw Materials
2.3. Storage
2.4. Analytical Determinations
2.4.1. Physico-Chemical Parameters
2.4.2. Lipid Oxidation
2.4.3. Lipid Extraction
2.4.4. Total Lipid Profile
2.4.5. Total Fatty Acid Profile
2.4.6. Nuclear Magnetic Resonance (1HR-NMR) Metabolomics for Quality Indexes
2.5. Statistical Analysis
3. Results and Discussion
3.1. Comparison between Manually and Mechanically Separated Fresh Flesh
3.2. Variation of Quality Indices during Frozen Storage
3.3. Data Correlation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsagarakis, K.; Palialexis, A.; Vassilopoulou, V. Mediterranean fishery discards: Review of the existing knowledge. ICES J. Marine Sci. 2014, 71, 1219–1234. [Google Scholar] [CrossRef] [Green Version]
- FAO. The State of Mediterranean and Black Sea Fisheries; General Fisheries Commission for the Mediterranean: Rome, Italy, 2016. [Google Scholar]
- Secci, G.; Borgogno, M.; Lupi, P.; Rossi, S.; Paci, G.; Mancini, S.; Bonelli, A.; Parisi, G. Effect of mechanical separation process on lipid oxidation in European aquacultured sea bass, gilthead sea bream, and rainbow trout products. Food Cont. 2016, 67, 75–81. [Google Scholar] [CrossRef]
- Hamed, I.; Özogul, F.; Regenstein, J.M. Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): A review. Trends Food Sci. Technol. 2016, 48, 40–50. [Google Scholar] [CrossRef]
- Ghaly, A.E.; Dave, D.; Budge, S.; Brooks, M.S. Fish Spoilage Mechanisms and Preservation Techniques: Review. Am. J. Appl. Sci. 2010, 7, 859–877. [Google Scholar] [CrossRef] [Green Version]
- IREPA Strutture Produttive. Andamento Della Pesca.Rapporto Annuale; Edizioni Scientifiche Italiane s.p.a.: Napoli, Italy, 2012. [Google Scholar]
- Mili, S.; Bouriga, N.; Missaoui, H.; Jarboui, O. Morphometric, Reproductive Parameters and Seasonal Variations in Fatty Acid Composition of the Mantis Shrimp Squilla mantis (Crustacea: Stomatopoda) in the Gulf of Gabes (Tunisia). J. Life Sci. 2011, 5, 1058–1071. [Google Scholar]
- Abelló, P.; Martín, P. Fishery dynamics of the mantis shrimp Squilla mantis (Crustacea: Stomatopoda) population off the Ebro delta (northwestern Mediterranean). Fish. Res. 1993, 16, 131–145. [Google Scholar] [CrossRef]
- Bao, Y.; Ertbjerg, P. Relationship between oxygen concentration, shear force and protein oxidation in modified atmosphere packaged pork. Meat Sci. 2015, 110, 174–179. [Google Scholar] [CrossRef]
- Bligh, G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Gallina Toschi, T.; Cardenia, V.; Bonaga, G.; Mandrioli, M.; Rodriguez-Estrada, M.T. Coffee silverskin: Characterization, possible uses, and safety aspects. J. Agri. Food Chem. 2014, 62, 10836–10844. [Google Scholar] [CrossRef]
- Luise, D.; Cardenia, V.; Zappaterra, M.; Motta, V.; Bosi, P.; Rodriguez-Estrada, M.T.; Trevisi, P. Evaluation of breed and parity order effects on the lipid composition of porcine colostrum. J. Agric. Food Chem. 2018, 66, 12911–12920. [Google Scholar] [CrossRef]
- European Commission; Allegate, X.B. regulation n. 796/02. Off. J. Eur. Communities 2002, L128, 14–18. [Google Scholar]
- Picone, G.; Engelsen, S.B.; Savorani, F.; Testi, S.; Badiani, A.; Capozzi, F. Metabolomics as a powerful tool for molecular quality assessment of the fish Sparus aurata. Nutrients 2011, 212–227. [Google Scholar] [CrossRef]
- Tappi, S.; Rocculi, P.; Ciampa, A.; Romani, S.; Balestra, F.; Capozzi, F.; Dalla Rosa, M. Computer vision system (CVS): A powerful non-destructive technique for the assessment of red mullet (Mullus barbatus) freshness. Eur. Food Res. Technol. 2017, 243, 2225–2233. [Google Scholar] [CrossRef]
- Rocculi, P.; Cevoli, C.; Tappi, S.; Genovese, J.; Urbinati, E.; Picone, G.; Fabbri, A.; Capozzi, F.; Dalla Rosa, M. Freshness assessment of European hake (Merluccius merluccius) through the evaluation of eye chromatic and morphological characteristics. Food Res. Int. 2019, 115, 234–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secci, G.; Borgogno, M.; Mancini, S.; Paci, G.; Parisi, G. Mechanical separation process for the value enhancement of Atlantic horse mackerel (Trachurus trachurus), a discard fish. Innov. Food Sci. Emerg. Technol. 2017, 39, 13–18. [Google Scholar] [CrossRef]
- Sundararajan, S.; Prudente, A.D.; Bankston, J.; King, J.M.; Wilson, P.; Sathivel, S. Evaluation of Green Tea Extract as a Glazing Material for Shrimp Frozen by Cryogenic Freezing. J. Food Sci. 2011, 76, E511–E518. [Google Scholar] [CrossRef]
- Passi, S.; Cataudella, S.; Di Marco, P.; De Simone, F.; Rastrelli, L. Fatty Acid Composition and Antioxidant Levels in Muscle Tissue of Different Mediterranean Marine Species of Fish and Shellfish. J. Agric. Food Chem. 2002, 50, 7314–7322. [Google Scholar] [CrossRef]
- Pacetti, D.; Mozzon, M.; Lucci, P.; Frega, N. Bioactive fish fatty acids: Health effects and their use as functional food ingredients. Curr. Nutr. Food Sci. 2013, 9, 283–297. [Google Scholar] [CrossRef]
- Olsson, G.B.; Olsen, R.L.; Carleho, M. Seasonal variations in chemical and sensory characteristics of farmed and wild Atlantic halibut ( Hippoglossus hippoglossus). Aquaculture 2003, 217, 191–205. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6 / omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef]
- Chéret, R.; Chapleau, N.; Delbarre-Ladrat, C.; Verrez-Bagnis, V.; Lamballerie, M.D. Effects of high pressure on texture and microstructure of sea bass (Dicentrarchus labrax L.) fillets. J. Food Sci. 2005, 70, e477–e483. [Google Scholar] [CrossRef] [Green Version]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Food Chemistry Changes of pigments and color in sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) muscle during iced storage. Food Chem. 2005, 93, 607–617. [Google Scholar] [CrossRef]
- Tsironi, T.; Dermesonlouoglou, E.; Giannakourou, M.; Taoukis, P. Shelf life modelling of frozen shrimp at variable temperature conditions. LWT Food Sci. Technol. 2009, 42, 664–671. [Google Scholar] [CrossRef]
- Torres, J.A.; Saraiva, J.A.; Guerra-rodríguez, E.P.; Aubourg, S.; Vázquez, M. Effect of combining high-pressure processing and frozen storage on the functional and sensory properties of horse mackerel (Trachurus trachurus). Innov. Food Sci. Emerg. Technol. 2014, 21, 2–11. [Google Scholar] [CrossRef]
- García-Soto, B.; Miranda, J.M.; Barros-Velázquez, J.; Aubourg, S.P. Quality changes during the frozen storage of the crustacean lobster krill (Munida spp.). Eur. J. Lipid Sci. Technol. 2015, 117, 431–439. [Google Scholar] [CrossRef]
- Vareltzis, K.; Koufidis, D.; Gavriilidou, E.; Papavergou, E.; Vasiliadou, S. Effectiveness of a natural Rosemary (Rosmarinus officinalis) extract on the stability of filleted and minced fish during frozen storage. Z. Für Lebensm. Und-Forsch. A 1997, 205, 93–96. [Google Scholar] [CrossRef]
- Zhang, B.; Ma, L.; Deng, S.; Xie, C.; Qiu, X. Shelf-life of pacific white shrimp (Litopenaeus vannamei) as affected by weakly acidic electrolyzed water ice-glazing and modified atmosphere packaging. Food Control. 2015, 51, 114–121. [Google Scholar] [CrossRef]
- Eymard, S.; Baron, C.P.; Jacobsen, C. Oxidation of lipid and protein in horse mackerel (Trachurus trachurus) mince and washed minces during processing and storage. Food Chem. 2009, 114, 57–65. [Google Scholar] [CrossRef]
- Rudy, M.D.; Kainz, M.J.; Graeve, M.; Colombo, S.M.; Arts, M.T. Handling and storage procedures have variable effects on fatty acid content in fishes with different lipid quantities. PLoS ONE 2016, 11, e0160497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeonidou, S.; Govaris, A.; Vareltzis, K. Effect of frozen storage on the quality of whole fish and fillets of horse mackerel (Trachurus trachurus) and mediterranean hake (Merluccius mediterraneus). Z. Für Lebensm. Und-Forsch. A 1997, 204, 405–410. [Google Scholar] [CrossRef]
- Badii, F.; Howell, N.K. A comparison of biochemical changes in cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) fillets during frozen storage. J. Sci. Food Agri. 2002, 82, 87–97. [Google Scholar] [CrossRef]
- Ólafsdóttir, G.; Martinsdóttir, E.; Oehlenschläger, J.; Dalgaard, P.; Jensen, B.; Undeland, I.; Mackie, I.M.; Henehan, G.; Nielsen, J.; Nilsen, H. Methods to evaluate fish freshness in research and industry. Trends Food Sci. Technol. 1997, 8, 258–265. [Google Scholar] [CrossRef]
- Sotelo, C.G.; Gallardo, J.M.; Pineiro, C.; Pérez-Martin, R. Trimethylamine oxide and derived compounds’ changes during frozen storage of hake (Merluccius merluccius). Food Chem. 1995, 53, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Özden, Ö. Changes in amino acid and fatty acid composition during shelf-life of marinated fish. J. Sci. Food Agric. 2005, 85, 2015–2020. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Moral, A. Changes in free amino acids during chilled storage of hake (Merluccius merluccius L.) in controlled atmospheres and their use as a quality control index. Eur. Food Res. Technol. 2001, 212, 302–307. [Google Scholar] [CrossRef]





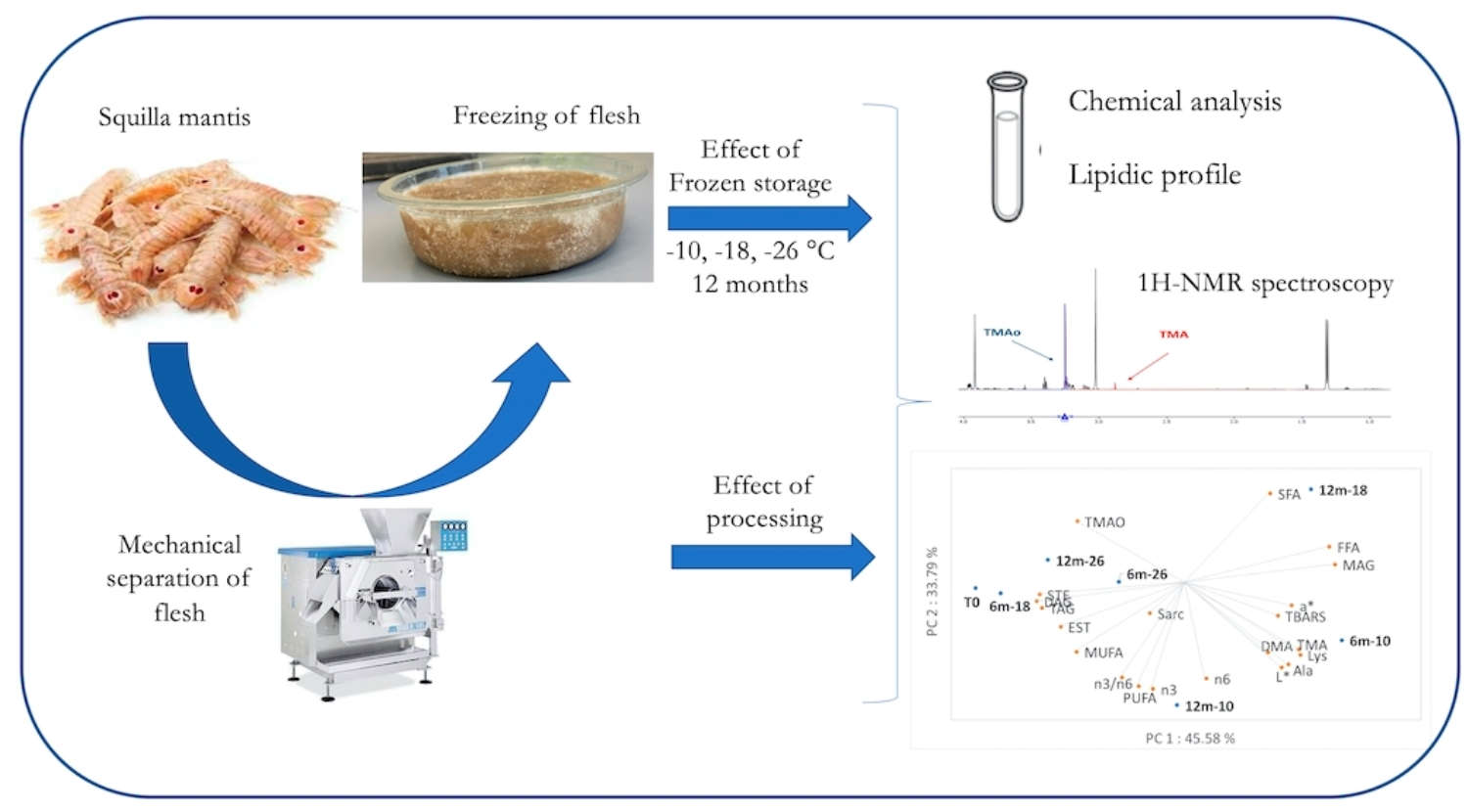{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| FF | MSF | |
|---|---|---|
| Water content (%) | 85.19 ± 0.71 a | 86.19 ± 0.14 a |
| pH | 6.71 ± 0.03 a | 6.74 ± 0.01 a |
| L* | 41.33 ± 0.05 a | 40.93 ± 0.09 b |
| a* | 4.77 ± 0.14 a | 5.56 ± 0.08 a |
| TBARs (mg MDA/kg) | 0.67± 0.03 b | 1.72 ± 0.31 a |
| Total lipid content (%) | 3.01 ± 0.42 a | 2.85 ± 0.35 a |
| FFA (% of total lipid) | 12.74 ± 2.28 a | 9.46 ± 0.18 a |
| MAG (% of total lipids) | 5.10 ± 0.78 a | 6.75 ± 0.13 a |
| DAG (% of total lipids) | 29.94 ± 0.91 a | 31.24 ± 0.19 a |
| TAG (% of total lipids) | 13.78 ± 8.48 a | 17.56 ± 0.03 a |
| EST (% of total lipids) | 28.50 ± 1.32 a | 28.47 ± 0.09 a |
| STE (% of total lipids) | 5.42 ± 1.38 a | 4.97 ± 0.26 a |
| SFA (% of total FA) | 31.27 ± 3.09 a | 30.17 ± 0.77 a |
| MUFA (% of total FA) | 55.72 ± 3.27 a | 57.87 ± 0.48 a |
| PUFA (% of total FA) | 13.00 ± 0.63 a | 11.95 ± 0.29 a |
| PUFA n-3 (% of total FA) | 10.18 ± 0.04 a | 9.35 ± 0.39 a |
| PUFA n-6 (% of total FA) | 2.82 ± 0.27 a | 2.78 ± 0.11 a |
| PUFA n-6/PUFA n-3 | 3.64 ± 0.56 a | 3.69 ± 0.30 a |
| Parameter | |||
|---|---|---|---|
| L* | a* | TBARs | |
| St | 267.82 *** | 48.77 *** | 5.91 *** |
| T | 2933.21 *** | 0.65 ns | 1.64 ns |
| St T | 160.82 *** | 13.41 *** | 2.01 * |
| Storage Temperature | Fat Content | FFA | MAG | DAG | TAG | STE | EST |
|---|---|---|---|---|---|---|---|
| (%) | % of Total Lipids | ||||||
| T0 | |||||||
| - | 2.9 ± 0.4 | 9.5 ± 0.2 d | 6.8 ± 0.1 de | 33.5 ±0.2 a | 17.6 ± 0.0 a | 4.9 ± 0.3 a | 27.9 ± 0.9 ab |
| 6 months | |||||||
| −10 °C | 1.7 ± 0.1 | 41.3 ± 0.5 a | 42.3 ± 0.5 a | 1.9 ± 0.1 e | 1.4 ± 0.2 d | 1.3 ± 0.2 cd | 11.9 ± 0.4 de |
| −18 °C | 1.9 ± 0.1 | 22.2 ± 1.7 c | 18.0 ± 1.8 c | 20.8 ± 1.4 c | 12.1 ± 0.3 b | 2.7 ± 0.1 b | 24.2 ± 1.8 b |
| −26 °C | 1.8 ± 0.8 | 12.8 ± 0.8 d | 9.1 ± 0.3 de | 26.7 ± 0.2 b | 18.6 ± 0.5 a | 4.4 ± 0.0 a | 28.4 ± 0.5 a |
| 12 months | |||||||
| −10 °C | 2.3 ± 1.0 | 47.6 ± 5.1 a | 39.0 ± 3.8 a | 0.4 ± 0.1 e | 1.1 ± 0.7 d | 0.9 ± 0.2 d | 11.0 ± 0.6 e |
| −18 °C | 2.1 ± 0.2 | 31.7 ± 2.4 b | 26.1 ± 1.6 b | 16.2 ±0.3 d | 6.3 ± 1.0 c | 2.0 ± 0.0 bc | 15.2 ± 0.2 cd |
| −26 °C | 2.6 ± 0.3 | 21.5 ± 0.9 c | 12.0 ± 1.4 d | 31.4 ± 2.2 a | 13.6 ± 1.4 b | 4.4 ± 0.5 a | 17.1 ± 2.1 c |
| Factor | F value | ||||||
| St | 7.04 * | 223.79 *** | 254.23 *** | 777.70 *** | 407.13 *** | 167.80 *** | 287.78 *** |
| T | 0.16 ns | 223.38 *** | 246.77 *** | 692.27 *** | 50.82 *** | 118.70 *** | 96.44 *** |
| St T | 0.26 ns | 31.96 *** | 68.18 *** | 187.70 *** | 98.24 *** | 31.11 *** | 40.01 *** |
| SFA | MUFA | PUFA | n-3 | n-6 | n-6/n3 | ||
| % of total fatty acids | |||||||
| T0 | |||||||
| - | 30.2 ± 0.8 b | 57.9 ± 0.5 a | 12.0 ± 0.3 a | 9.4 ± 0.4 ab | 2.8 ± 0.1 | 3.5 ± 0.3 | |
| 6 months | |||||||
| −10 °C | 31.8 ± 0.6 ab | 55.9 ± 0.2 ab | 12.3 ± 0.4 a | 9.6 ± 0. 3 ab | 2.7 ± 0.1 | 3.5 ± 0.1 | |
| −18 °C | 30.5 ± 1.5 b | 57.6 ± 0.4 a | 12.0 ± 1.1 a | 9.4 ± 1.1 ab | 2.6 ± 0.0 | 3.5 ± 0.4 | |
| −26 °C | 29.8 ± 1.0 b | 57.4 ± 0.9 a | 12.7 ± 0.1 a | 10.0 ± 0.0 ab | 2.7 ± 0.1 | 3.5 ± 0.1 | |
| 12 months | |||||||
| −10 °C | 27.5 ± 5.1 b | 58.2 ± 2.9 a | 14.4 ± 2.1 a | 11.2 ± 1.1 a | 3.2 ± 1.1 | 3.6 ±0.9 | |
| −18 °C | 40.2 ± 1.2 a | 51.3 ± 1.9 b | 7.5 ± 0.7 b | 5.0 ±1.5 b | 2.5 ± 0.6 | 2.9 ± 0.1 | |
| −26 °C | 30.2 ± 1.5 b | 58.6 ± 0.6 a | 11.2 ± 0.8 a | 8.6 ± 0.7 ab | 2.6 ± 0.1 | 3.4 ± 0.1 | |
| Factor | F value | ||||||
| St | 2.57 ns | 3.25 ns | 3.29 ns | 1.29 ns | 0.19 ns | 1.02 ns | |
| T | 6.95 * | 5.71 * | 10.59 ** | 5.39 * | 0.57 ns | 0.60 ns | |
| St T | 8.34 ** | 8.26 ** | 9.07 ** | 4.78 * | 0.52 ns | 0.72 ns | |
| Factor | Parameter | |||||
|---|---|---|---|---|---|---|
| TMA-O | TMA | DMA | Lys | Sarc | Ala | |
| St | 32.58 *** | 21.24 *** | 16.61 *** | 8.02 *** | 81.18 *** | 7.84 *** |
| T | 58.10 *** | 41.77 *** | 68.93 *** | 35.67 *** | 0.35 ns | 30.04 *** |
| St T | 45.17 *** | 6.30 *** | 7.66 ** | 4.15 *** | 2.17 * | 3.27 ** |
| L* | a* | TBARs | TMA-O | TMA | DMA | Lys | Sarc | Ala | FFA | MAG | TAG | SFA | MUFA | PUFA | n-6 | n-3 | n-6/n-3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L* | - | |||||||||||||||||
| a* | 0.570 | - | ||||||||||||||||
| TBARs | 0.530 | 0.429 | - | |||||||||||||||
| TMAO | −0.858 * | −0.478 | −0.803 * | - | ||||||||||||||
| TMA | 0.921 * | 0.377 | 0.561 | −0.873 * | - | |||||||||||||
| DMA | 0.846 * | 0.237 | 0.158 | −0.642 | 0.903 * | - | ||||||||||||
| Lys | 0.939 * | 0.462 | 0.647 | −0.890 * | 0.986 * | 0.847 * | - | |||||||||||
| Sarc | −0.099 | −0.096 | 0.384 | 0.083 | −0.121 | −0.323 | 0.009 | - | ||||||||||
| Ala | 0.971 * | 0.456 | 0.610 | −0.888 * | 0.976 * | 0.863 * | 0.990 * | −0.004 | - | |||||||||
| FFA | 0.882 * | 0.367 | 0.458 | −0.874 * | 0.958 * | 0.909 * | 0.914 * | −0.367 | 0.911 * | - | ||||||||
| MAG | 0.894 * | 0.412 | 0.638 | −0.928 * | 0.984 * | 0.845 * | 0.972 * | −0.160 | 0.953 * | 0.969 * | - | |||||||
| TAG | −0.858 * | −0.335 | −0.545 | 0.894 * | −0.966 * | −0.864 * | −0.924 * | 0.312 | −0.909 * | −0.984 * | −0.981 * | - | ||||||
| SFA | −0.228 | 0.020 | 0.067 | 0.037 | 0.038 | −0.074 | −0.023 | −0.331 | −0.136 | 0.061 | 0.091 | −0.167 | - | |||||
| MUFA | 0.021 | −0.139 | −0.167 | 0.110 | −0.255 | −0.136 | −0.206 | 0.266 | −0.087 | −0.241 | −0.290 | 0.339 | −0.963 * | - | ||||
| PUFA | 0.443 | 0.128 | 0.096 | −0.221 | 0.211 | 0.282 | 0.286 | 0.408 | 0.381 | 0.143 | 0.147 | −0.041 | −0.951 * | 0.832 * | - | |||
| n-6 | 0.513 | 0.155 | 0.088 | −0.250 | 0.310 | 0.399 | 0.380 | 0.392 | 0.465 | 0.229 | 0.233 | −0.125 | −0.896 * | 0.742 | 0.986 * | - | ||
| n-3 | 0.715 | 0.322 | −0.067 | −0.344 | 0.574 | 0.767 * | 0.583 | −0.047 | 0.649 | 0.551 | 0.480 | −0.430 | −0.592 | 0.387 | 0.747 | 0.829 * | - | |
| n-6/n-3 | 0.286 | −0.043 | 0.167 | −0.136 | 0.103 | 0.111 | 0.193 | 0.630 | 0.273 | −0.015 | 0.041 | 0.081 | −0.917 * | 0.820 * | 0.954 * | 0.927 * | 0.565 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tappi, S.; De Aguiar Saldanha Pinheiro, A.C.; Mercatante, D.; Picone, G.; Soglia, F.; Rodriguez-Estrada, M.T.; Petracci, M.; Capozzi, F.; Rocculi, P. Quality Changes during Frozen Storage of Mechanical-Separated Flesh Obtained from an Underutilized Crustacean. Foods 2020, 9, 1485. https://doi.org/10.3390/foods9101485
Tappi S, De Aguiar Saldanha Pinheiro AC, Mercatante D, Picone G, Soglia F, Rodriguez-Estrada MT, Petracci M, Capozzi F, Rocculi P. Quality Changes during Frozen Storage of Mechanical-Separated Flesh Obtained from an Underutilized Crustacean. Foods. 2020; 9(10):1485. https://doi.org/10.3390/foods9101485
Chicago/Turabian StyleTappi, Silvia, Ana Cristina De Aguiar Saldanha Pinheiro, Dario Mercatante, Gianfranco Picone, Francesca Soglia, Maria Teresa Rodriguez-Estrada, Massimiliano Petracci, Francesco Capozzi, and Pietro Rocculi. 2020. "Quality Changes during Frozen Storage of Mechanical-Separated Flesh Obtained from an Underutilized Crustacean" Foods 9, no. 10: 1485. https://doi.org/10.3390/foods9101485