Understanding Antimicrobial Resistance (AMR) Profiles of Salmonella Biofilm and Planktonic Bacteria Challenged with Disinfectants Commonly Used During Poultry Processing


Abstract
:1. Introduction
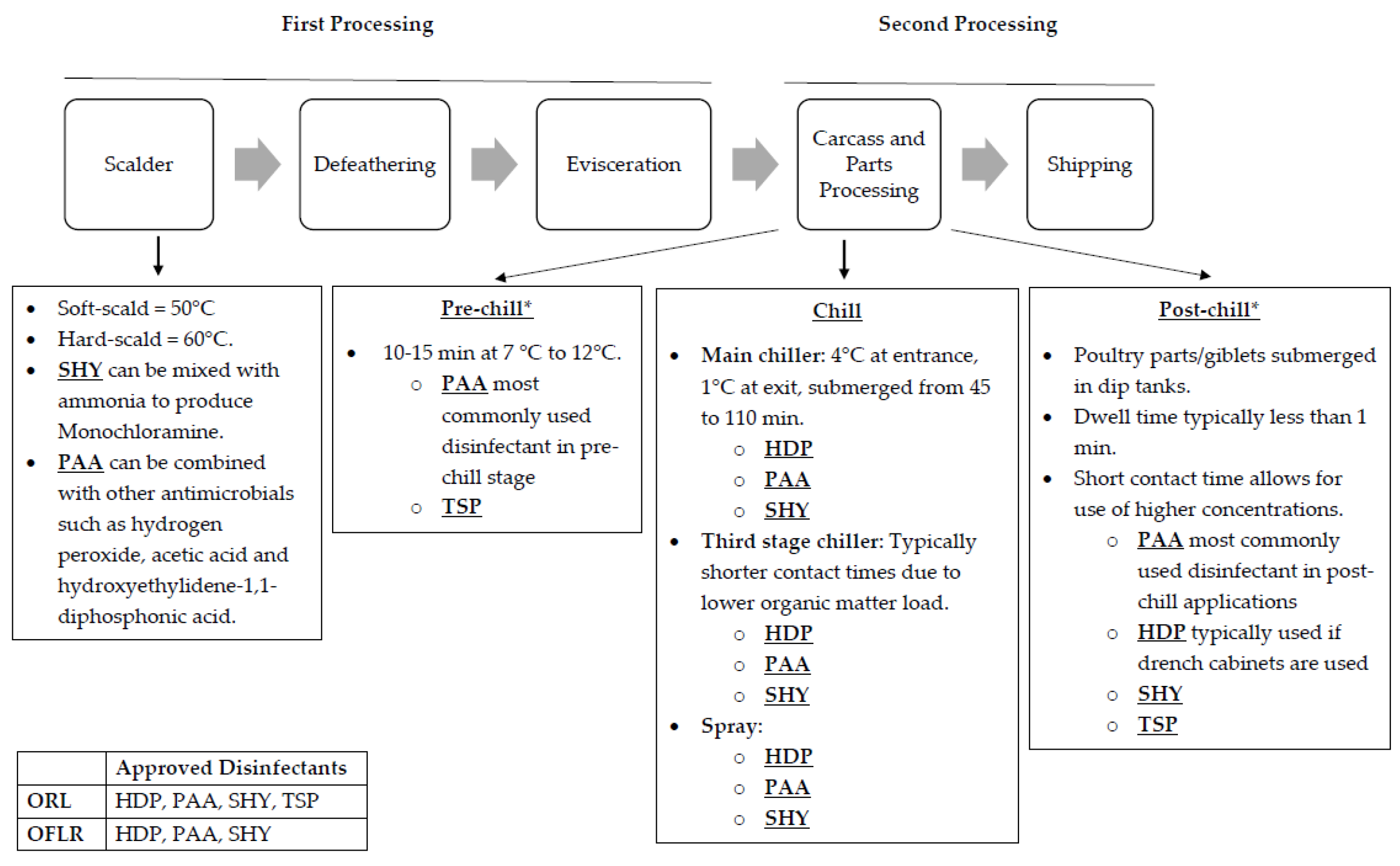
2. Disinfectants Commonly Used during Poultry Processing
3. Poultry Processing Methods Conferring Biocide Tolerance
4. Proposed Mechanisms of Bacterial Resistance to Antimicrobials Induced by Disinfectants Used During Poultry Processing
5. Antimicrobial Resistance Profiles of Foodborne Pathogens Challenged with Disinfectants
6. Biofilm Detection
7. Biofilm Characterization
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- CDC. Salmonella. Available online: https://www.cdc.gov/salmonella/index.html (accessed on 20 July 2019).
- Youn, S.Y.; Jeong, O.M.; Choi, B.K.; Jung, S.C.; Kang, M.S. Comparison of the antimicrobial and sanitizer resistance of Salmonella isolates from chicken slaughter processes in Korea. J. Food Sci. 2017, 82, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Mion, L.; Parizotto, L.; Calasans, M.; Dickel, E.L.; Pilotto, F.; Rodrigues, L.B.; Nascimento, V.P.; Santos, L.R. Effect of antimicrobials on Salmonella spp. strains isolated from poultry processing plants. Braz. J. Poult. Sci. 2016, 18, 337–341. [Google Scholar] [CrossRef]
- Marquez, M.L.F.; Burgos, M.J.G.; Pulido, R.P.; Galvez, A.; Lopez, R.L. Biocide tolerance and antibiotic resistance in Salmonella isolates from hen eggshells. Foodborne Pathog. Dis. 2017, 14, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Marquez, M.L.F.; Burgos, M.J.G.; Pulido, R.P.; Galvez, A.; Lopez, R.L. Correlations among Resistances to Different Antimicrobial Compounds in Salmonella Strains from Hen Eggshells. J. Food Prot. 2018, 81, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Wales, A.D.; Davies, R.H. Co-selection of resistance to antibiotics, biocides and heavy metals, and its relevance to foodborne pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef] [PubMed]
- Steenackers, H.; Hermans, K.; Vanderleyden, J.; De Keersmaecker, S.C.J. Salmonella biofilms: An overview on occurrence, structure, regulation and eradication. Food Res. Int. 2012, 45, 502–531. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Merino, L.; Procura, F.; Trejo, F.M.; Bueno, D.J.; Golowczyc, M.A. Biofilm formation by Salmonella sp. in the poultry industry: Detection, control and eradication strategies. Food Res. Int. 2017, 119, 530–540. [Google Scholar] [CrossRef]
- Moraes, J.O.; Cruz, E.A.; Souza, E.G.F.; Oliveira, T.C.M.; Alvarenga, V.O.; Peña, W.E.L.; Sant’Ana, A.S.; Magnani, M. Predicting adhesion and biofilm formation boundaries on stainless steel surfaces by five Salmonella enterica strains belonging to different serovars as a function of pH, temperature and NaCl concentration. Int. J. Food Microbiol. 2018, 281, 90–100. [Google Scholar] [CrossRef]
- Li, J.; Feng, J.; Ma, L.; de la Fuente Núñez, C.; Gölz, G.; Lu, X. Effects of meat juice on biofilm formation of Campylobacter and Salmonella. Int. J. Food Microbiol. 2017, 253, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giaouris, E.E.; Simões, M.V. Pathogenic Biofilm Formation in the Food Industry and Alternative Control Strategies. In Foodborne Diseases; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 309–377. [Google Scholar]
- Sereno, M.; Ziech, R.; Druziani, J.; Pereira, J.; Bersot, L. Antimicrobial Susceptibility and Biofilm Production by Salmonella sp. Strains Isolated from Frozen Poultry Carcasses. Braz. J. Poult. Sci. 2017, 19, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Stoodley, P. Evolving concepts in biofilm infections. Cell. Microbiol. 2009, 11, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- Ziech, R.E.; Perin, A.P.; Lampugnani, C.; Sereno, M.J.; Viana, C.; Soares, V.M.; Pereira, J.G.; Pinto, J.P.d.A.N.; Bersot, L.d.S. Biofilm-producing ability and tolerance to industrial sanitizers in Salmonella spp. isolated from Brazilian poultry processing plants. LWT Food Sci. Technol. 2016, 68, 85–90. [Google Scholar] [CrossRef] [Green Version]
- Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Salmonella and Campylobacter biofilm formation: A comparative assessment from farm to fork. J. Sci. Food Agric. 2018, 98, 4014–4032. [Google Scholar] [CrossRef] [PubMed]
- Rossi, D.A.; Melo, R.T.; Mendonça, E.P.; Monteiro, G.P. Biofilms of Salmonella and Campylobacter in the poultry industry. In Poultry Science; IntechOpen: London, UK, 2017. [Google Scholar]
- Turki, Y.; Ouzari, H.; Mehri, I.; Ben Aissa, R.; Hassen, A. Biofilm formation, virulence gene and multi-drug resistance in Salmonella Kentucky isolated in Tunisia. Food Res. Int. 2012, 45, 940–946. [Google Scholar] [CrossRef]
- Bell, D.D.; Weaver, W.D. Commercial Chicken Meat and Egg Production, 5th ed.; Springer: New York, NY, USA, 2002; Volume 11, pp. 224–225. [Google Scholar]
- Russell, S.M. Controlling Salmonella in Poultry Production and Processing; Crc Press-Taylor & Francis Group: Boca Raton, FL, USA, 2012. [Google Scholar]
- FSIS. Modernization of Poultry Slaughter Inspection. 9 CFR Parts 381 and 500; 2014. Available online: https://www.fsis.usda.gov/wps/wcm/connect/00ffa106-f373-437a-9cf3-6417f289bfc2/2011-0012.pdf?MOD=AJPERES (accessed on 20 July 2019).
- Rasekh, J.; Thaler, A.M.; Engeljohn, D.L.; Pihkala, N.H. Food Safety and Inspection Service policy for control of poultry contaminated by digestive tract contents: A review. J. Appl. Poult. Res. 2005, 14, 603–611. [Google Scholar] [CrossRef]
- USDA. Related Documents for FSIS Directive 7120.1—Safe and Suitable Ingredients Used in the Production of Meat, Poultry, and Egg Products. 2019. Available online: https://www.fsis.usda.gov/wps/portal/fsis/topics/regulations/directives/7000-series/safe-suitable-ingredients-related-document (accessed on 20 July 2019).
- Smith, J.; Corkran, S.; McKee, S.R.; Bilgili, S.F.; Singh, M. Evaluation of post-chill applications of antimicrobials against Campylobacter jejuni on poultry carcasses. J. Appl. Poult. Res. 2015, 24, 451–456. [Google Scholar] [CrossRef]
- Duan, D.B.; Wang, H.H.; Xue, S.W.; Li, M.; Xu, X.L. Application of disinfectant sprays after chilling to reduce the initial microbial load and extend the shelf-life of chilled chicken carcasses. Food Control 2017, 75, 70–77. [Google Scholar] [CrossRef]
- Chen, X.; Bauermeister, L.J.; Hill, G.N.; Singh, M.; Bilgili, S.F.; McKee, S.R. Efficacy of various antimicrobials on reduction of Salmonella and Campylobacter and quality attributes of ground chicken obtained from poultry parts treated in a postchill decontamination tank. J. Food Prot. 2014, 77, 1882–1888. [Google Scholar] [CrossRef]
- Morente, E.O.; Fernandez-Fuentes, M.A.; Burgos, M.J.G.; Abriouel, H.; Pulido, R.P.; Galvez, A. Biocide tolerance in bacteria. Int. J. Food Microbiol. 2013, 162, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Gadea, R.; Fuentes, M.A.F.; Pulido, R.P.; Galvez, A.; Ortega, E. Effects of exposure to quaternary-ammonium-based biocides on antimicrobial susceptibility and tolerance to physical stresses in bacteria from organic foods. Food Microbiol. 2017, 63, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Mavri, A.; Mozina, S.S. Development of antimicrobial resistance in Campylobacter jejuni and Campylobacter coli adapted to biocides. Int. J. Food Microbiol. 2013, 160, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Capita, R.; Alvarez-Fernandez, E.; Fernandez-Buelta, E.; Manteca, J.; Alonso-Calleja, C. Decontamination treatments can increase the prevalence of resistance to antibiotics of Escherichia coli naturally present on poultry. Food Microbiol. 2013, 34, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Finstad, S.; O’Bryan, C.A.; Marcy, J.A.; Crandall, P.G.; Ricke, S.C. Salmonella and broiler processing in the United States: Relationship to foodborne salmonellosis. Food Res. Int. 2012, 45, 789–794. [Google Scholar] [CrossRef]
- Chylkova, T.; Cadena, M.; Ferreiro, A.; Pitesky, M. Susceptibility of Salmonella biofilm and planktonic bacteria to common disinfectant agents used in poultry processing. J. Food Prot. 2017, 80, 1072–1079. [Google Scholar] [CrossRef]
- Shah, J.; Desai, P.T.; Chen, D.; Stevens, J.R.; Weimer, B.C. Preadaptation to cold stress in Salmonella enterica serovar Typhimurium increases survival during subsequent acid stress exposure. Appl. Environ. Microbiol. 2013, 79, 7281–7289. [Google Scholar] [CrossRef]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Molina-Gonzalez, D.; Alonso-Calleja, C.; Alonso-Hernando, A.; Capita, R. Effect of sub-lethal concentrations of biocides on the susceptibility to antibiotics of multi-drug resistant Salmonella enterica strains. Food Control 2014, 40, 329–334. [Google Scholar] [CrossRef]
- Alav, I.; Rahman, K.M.; Sutton, J.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [Green Version]
- Su, C.-C.; Yin, L.; Kumar, N.; Dai, L.; Radhakrishnan, A.; Bolla, J.R.; Lei, H.-T.; Chou, T.-H.; Delmar, J.A.; Rajashankar, K.R.; et al. Structures and transport dynamics of a Campylobacter jejuni multidrug efflux pump. Nat. Commun. 2017, 8, 171. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Xiong, Y.L.L. Technologies and mechanisms for safety control of ready-to-eat muscle foods: An updated review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1886–1901. [Google Scholar] [CrossRef] [PubMed]
- Wessels, S.; Ingmer, H. Modes of action of three disinfectant active substances: A review. Regul. Toxicol. Pharmacol. 2013, 67, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Su, X.W.; D’Souza, D.H. Reduction of Salmonella Typhimurium and Listeria monocytogenes on produce by trisodium phosphate. LWT Food Sci. Technol. 2012, 45, 221–225. [Google Scholar] [CrossRef]
- Strempel, N.; Nusser, M.; Neidig, A.; Brenner-Weiss, G.; Overhage, J. The oxidative stress agent hypochlorite stimulates c-di-GMP synthesis and biofilm formation in Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.Z.; Chen, P.X.; Yang, L.; Li, W.; Chang, M.X.; Jiang, H.X. Coordinated expression of acrAB-tolC and eight other functional efflux pumps through activating ramA and marA in Salmonella enterica serovar Typhimurium. Microb. Drug Resist. 2018, 24, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Urdaneta, V.; Casadesus, J. Adaptation of Salmonella enterica to bile: Essential role of AcrAB-mediated efflux. Environ. Microbiol. 2018, 20, 1405–1418. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Subirats, J.; Borrego, C.M. The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 2015, 6, 1216. [Google Scholar] [CrossRef]
- Doroshenko, N.; Tseng, B.S.; Howlin, R.P.; Deacon, J.; Wharton, J.A.; Thurner, P.J.; Gilmore, B.F.; Parsek, M.R.; Stoodley, P. Extracellular DNA Impedes the Transport of Vancomycin in Staphylococcus epidermidis Biofilms Preexposed to Subinhibitory Concentrations of Vancomycin. Antimicrob. Agents Chemother. 2014, 58, 7273–7282. [Google Scholar] [CrossRef] [Green Version]
- Özdemir, C.; Akçelik, N.; Akçelik, M. The Role of Extracellular DNA in Salmonella Biofilms. Mol. Genet. Microbiol. Virol. 2018, 33, 60–71. [Google Scholar] [CrossRef]
- Jia, K.; Wang, G.Y.; Liang, L.J.; Wang, M.; Wang, H.H.; Xu, X.L. Preliminary transcriptome analysis of mature biofilm and planktonic cells of Salmonella Enteritidis exposure to acid stress. Front. Microbiol. 2017, 8, 1861. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Ordóñez, A.; Prieto, M.; Bernardo, A.; Hill, C.; López, M. The Acid Tolerance Response of Salmonella spp.: An adaptive strategy to survive in stressful environments prevailing in foods and the host. Food Res. Int. 2012, 45, 482–492. [Google Scholar] [CrossRef]
- MacKenzie, K.D.; Wang, Y.; Shivak, D.J.; Wong, C.S.; Hoffman, L.J.L.; Lam, S.; Kröger, C.; Cameron, A.D.S.; Townsend, H.G.G.; Köster, W.; et al. Bistable expression of CsgD in Salmonella enterica serovar Typhimurium connects virulence to persistence. Infect. Immun. 2015, 83, 2312–2326. [Google Scholar] [CrossRef] [PubMed]
- Wideman, N.; Bailey, M.; Bilgili, S.F.; Thippareddi, H.; Wang, L.; Bratcher, C.; Sanchez-Plata, M.; Singh, M. Evaluating best practices for Campylobacter and Salmonella reduction in poultry processing plants. Poult. Sci. 2016, 95, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Biswal, B.K.; Khairallah, R.; Bibi, K.; Mazza, A.; Gehr, R.; Masson, L.; Frigon, D. Impact of UV and peracetic acid disinfection on the prevalence of virulence and antimicrobial resistance genes in uropathogenic Escherichia coli in wastewater effluents. Appl. Environ. Microbiol. 2014, 80, 3656–3666. [Google Scholar] [CrossRef] [PubMed]
- Karaca, B.; Akcelik, N.; Akcelik, M. Biofilm-producing abilities of Salmonella strains isolated from Turkey. Biologia 2013, 68, 1–10. [Google Scholar] [CrossRef]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef]
- Kırmusaoğlu, S. Antimicrobials, Antibiotic Resistance, Antibiofilm Strategies and Activity Methods; IntechOpen: London, UK, 2019. [Google Scholar]
- Wang, H.H.; Ye, K.P.; Wei, X.R.; Cao, J.X.; Xu, X.L.; Zhou, G.H. Occurrence, antimicrobial resistance and biofilm formation of Salmonella isolates from a chicken slaughter plant in China. Food Control 2013, 33, 378–384. [Google Scholar] [CrossRef]
- González-Rivas, F.; Ripolles-Avila, C.; Fontecha-Umaña, F.; Ríos-Castillo, A.G.; Rodríguez-Jerez, J.J. Biofilms in the Spotlight: Detection, Quantification, and Removal Methods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1261–1276. [Google Scholar] [CrossRef] [Green Version]
- Romanova, N.A.; Gawande, P.V.; Brovko, L.Y.; Griffiths, M.W. Rapid methods to assess sanitizing efficacy of benzalkonium chloride to Listeria monocytogenes biofilms. J. Microbiol. Methods 2007, 71, 231–237. [Google Scholar] [CrossRef]
- Lavilla Lerma, L.; Benomar, N.; Gálvez, A.; Abriouel, H. Prevalence of bacteria resistant to antibiotics and/or biocides on meat processing plant surfaces throughout meat chain production. Int. J. Food Microbiol. 2013, 161, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.W.; Silvers, S. Comparison of poultry processing equipment surfaces for susceptibility to bacterial attachment and biofilm formation. Poult. Sci. 2000, 79, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Okabe, S.; Hirata, K.; Watanabe, Y. Dynamic changes in spatial microbial distribution in mixed-population biofilms: Experimental results and model simulation. Water Sci. Technol. 1995, 32, 67–74. [Google Scholar] [CrossRef]
- Itram Higiene. BioFinder. 2019. Available online: https://biofilmremove.com/en/detection/ (accessed on 22 June 2019).
- REALCO. Biofilm Detection Kit. 2019. Available online: https://www.realco.be/en/our-markets/food-beverage/biofilm-audit/biofilm-detection-kit/ (accessed on 26 June 2019).
- Betelgeux-Christeyns. Detection. 2019. Available online: https://biofilmtest.com/detection/?lang=en (accessed on 26 June 2019).
- bioMérieux. TEMPO. 2019. Available online: https://www.biomerieux.com/en/biomerieux-launches-tempo-eb-first-automated-test-enterobacteriaceae-enumeration-food-products (accessed on 26 June 2019).
- Van Houdt, R.; Michiels, C. Biofilm formation and the food industry, a focus on the bacterial outer surface. J. Appl. Microbiol. 2010, 109, 1117–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelieveld, H.L.M.; Mostert, M.A.; Holah, J. Handbook of Hygiene Control in the Food Industry; Woodhead Publishing Ltd.: Cambridge, UK, 2005. [Google Scholar]
- Maukonen, J.; Mättö, J.; Wirtanen, G.; Raaska, L.; Mattila-Sandholm, T.; Saarela, M. Methodologies for the characterization of microbes in industrial environments: A review. J. Ind. Microbiol. Biotechnol. 2003, 30, 327–356. [Google Scholar] [CrossRef] [PubMed]
- Sy-Lab. BacTrac 4300 Microbiological Impedance Analyser. 2019. Available online: https://microbiology.sylab.com/products/p/show/Product/product/bactrac-4300.html (accessed on 26 June 2019).
- Wirtanen, G.; Salo, S.; Helander, I.M.; Mattila-Sandholm, T. Microbiological methods for testing disinfectant efficiency on Pseudomonas biofilm. Colloids Surf. B Biointerfaces 2001, 20, 37–50. [Google Scholar] [CrossRef]
- Abcam. XTT Sodium Salt, Tetrazolium Salt (ab146310). 2019. Available online: https://www.abcam.com/xtt-sodium-salt-tetrazolium-salt-ab146310.html (accessed on 26 June 2019).
- Abcam. Resazurin Assay Kit (Cell Viability) (ab129732). 2019. Available online: https://www.abcam.com/resazurin-assay-kit-cell-viability-ab129732.html (accessed on 26 June 2019).
- Olson, M.E.; Ceri, H.; Morck, D.W.; Buret, A.G.; Read, R.R. Biofilm bacteria: Formation and comparative susceptibility to antibiotics. Can. J. Vet. Res. 2002, 66, 86–92. [Google Scholar] [PubMed]
- Sarjit, A.; Dykes, G.A. Antimicrobial activity of trisodium phosphate and sodium hypochlorite against Salmonella biofilms on abiotic surfaces with and without soiling with chicken juice. Food Control 2017, 73, 1016–1022. [Google Scholar] [CrossRef]
- Korber, D.R.; Choi, A.; Wolfaardt, G.M.; Ingham, S.C.; Caldwell, D.E. Substratum topography influences susceptibility of Salmonella enteritidis biofilms to trisodium phosphate. Appl. Environ. Microbiol. 1997, 63, 3352–3358. [Google Scholar] [PubMed]
- Wang, H.H.; Zhang, X.X.; Dong, Y.; Xu, X.L.; Zhou, G.H. Insights into the transcriptome profile of mature biofilm of Salmonella Typhimurium on stainless steels surface. Food Res. Int. 2015, 77, 378–384. [Google Scholar] [CrossRef]
- Borges, K.A.; Furian, T.Q.; de Souza, S.N.; Menezes, R.; de Lima, D.A.; Fortes, F.B.B.; Salle, C.T.P.; Moraes, H.L.S.; Nascimento, V.P. Biofilm formation by Salmonella Enteritidis and Salmonella Typhimurium isolated from avian sources is partially related with their in vivo pathogenicity. Microb. Pathog. 2018, 118, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Influence of milk, chicken residues and oxygen levels on biofilm formation on stainless steel, gene expression and small RNAs in Salmonella enterica. Food Control 2018, 90, 1–9. [Google Scholar] [CrossRef]
- Wang, H.; Dong, Y.; Wang, G.; Xu, X.; Zhou, G. Effect of growth media on gene expression levels in Salmonella Typhimurium biofilm formed on stainless steel surface. Food Control 2016, 59, 546–552. [Google Scholar] [CrossRef]
- Lianou, A.; Koutsoumanis, K.P. Strain variability of the biofilm-forming ability of Salmonella enterica under various environmental conditions. Int. J. Food Microbiol. 2012, 160, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Vestby, L.K.; Møretrø, T.; Langsrud, S.; Heir, E.; Nesse, L.L. Biofilm forming abilities of Salmonellaare correlated with persistence in fish meal- and feed factories. Bmc Vet. Res. 2009, 5, 20. [Google Scholar] [CrossRef]
- Aksoy, D. DETERMINATION OF in vitro BIOFILM FORMATION ABILITIES OF FOOD BORNE Salmonella enterica ISOLATES. Trak. Univ. J. Nat. Sci. 2019, 20, 57–62. [Google Scholar] [CrossRef]
- Anonymous. The increasing challenge of biofilms. Int. Food Hyg. 2008, 18, 11–12. [Google Scholar]
- Ju, X.; Li, J.; Zhu, M.; Lu, Z.; Lv, F.; Zhu, X.; Bie, X. Effect of the luxS gene on biofilm formation and antibiotic resistance by Salmonella serovar Dublin. Food Res. Int. 2018, 107, 385–393. [Google Scholar] [CrossRef]
- Ripolles-Avila, C.; Hascoet, A.S.; Guerrero-Navarro, A.E.; Rodriguez-Jerez, J.J. Establishment of incubation conditions to optimize the in vitro formation of mature Listeria monocytogenes biofilms on food-contact surfaces. Food Control 2018, 92, 240–248. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.-J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Ramaswamy, R. Application of emerging technologies to control Salmonella in foods: A review. Food Res. Int. 2012, 45, 666–677. [Google Scholar] [CrossRef]


{kind=link}
| Disinfectant | Disinfectant Type | Proposed Modes of Action | References | Proposed Mechanism Conferring Antibiotic Resistance | References |
|---|---|---|---|---|---|
| HDP | Quaternary ammonium |
| [30,39] |
| [39] |
| PAA | Organic acid and an oxidant |
| [40] |
| [40] |
| SHY | Chlorine |
| [41] |
| [42] |
| TSP | Alkaline detergent |
| [30,41] |
| [30] |
| Reference | Disinfectant | Application Parameters | Isolate | AMR Profile | |||
|---|---|---|---|---|---|---|---|
| [29] | Showed increased resistance (i.e., twice the MIC) to the following antimicrobials compared to the wildtype: | Showed decreased resistance to the following antimicrobials compared to the wildtype: | |||||
| HDP | Exposed to increasing concentrations of CPC (0.01, 0.1, 1, 5, 10, 50, 100, 200, 500 mg/mL, 1, 2, 5 and 10 mg/mL). | Salmonella UJA59l | Ampicillin, Sulfamethoxazole, Nalidixic acid | Ceftazidime | |||
| Salmonella UJA82k | Ceftazidime | ||||||
| Salmonella UJA82l | Nalidixic acid | Ampicillin, Cefotaxime, Ceftazidime, Sulfametoxazol | |||||
| [4] | HDP tolerance level: | Showed resistance to: | |||||
| HDP | HDP tolerance and antibiotic resistance were determined by using MIC assays. | Salmonella spp. UJAS6 | Tolerant | Ampicillin, Chloramphenicol, Tetracycline, Nalidixic acid, Trimethoprim-sulfamethoxazole | |||
| S. enterica UJAS10 | Tolerant | Ampicillin, Tetracycline, Nalidixic acid, Trimethoprim-sulfamethoxazole | |||||
| Salmonella spp. UJAS18 | Tolerant | Ampicillin, Cefotaxime; Ceftazidime, Ciprofloxacin, Chloramphenicol, Tetracycline, Netilmicin, Nalidixic acid, Trimethoprim-sulfamethoxazole | |||||
| Salmonella spp. UJAS19 | Tolerant | Cefotaxime; Ceftazidime, Ciprofloxacin, Chloramphenicol, Streptomycin, Tetracycline, Netilmicin, Nalidixic acid, Trimethoprim-sulfamethoxazole | |||||
| [30] | HDP | Step-wise exposure to gradually increasing concentrations (2, 2.5, 3, 4 to 5 mg/mL, depending upon the growth of the adapted microorganism) of HDP over 15 days. | MIC fold change of Campylobacter strains relative to the pre-adapted strains. | ||||
| Campylobacter jejuni K49/4 | Days after repeated exposure to HDP | 5 | 10 | 15 | |||
| MIC fold change | 1 | 1 | 1 | ||||
| Campylobacter jejuni NCTC11168 | Days after repeated exposure to HDP | 5 | 10 | 15 | |||
| MIC fold change | 2 | 1 | 4 | ||||
| Campylobacter jejuni ATCC33560 | Days after repeated exposure to HDP | 5 | 10 | 15 | |||
| MIC fold change | 0.5 | 1 | 1 | ||||
| Campylobacter coli 137 | Days after repeated exposure to HDP: | 5 | 10 | 15 | |||
| MIC fold change | 1 | 1 | 0.5 | ||||
| Campylobacter coli ATCC33559 | Days after repeated exposure to HDP | 5 | 10 | 15 | |||
| MIC fold change | 1 | 2 | 2 | ||||
| [52] | Number of antimicrobial resistance gene classes in PAA treated strains: | ||||||
| PAA | Exposed to 0.9 to 2.0 mg/L of PAA to reach target disinfection level of 200 CFU/100mL | Escherichia coli | Mean number of classes decreased by an average of 47% with significant reductions in the following classes: Macrolides (−62.3%), Beta-lactams (−41.3), Phenicols (−64) and Trimethoprim (−49.9). | ||||
| [36] | Showed increased resistance (i.e., susceptible to resistant via disk diffusion assay) after exposure to disinfectant: | ||||||
| SHY | Exposed to increasing sub-inhibitory concentrations (starting at MIC/2). | Salmonella Enteritidis | Ceftazidime | ||||
| Salmonella Kentucky | Amikacin, Ampicillin/ sulbactam | ||||||
| Salmonella Typhimurium | Amikacin, Tobramycin, Cefazolin, Cefotaxime | ||||||
| TSP | Salmonella Enteritidis | Amikacin, Cefazolin, Cefoxitin, Ceftazidime, Aztreonam, Nalidixic acid, Phosphomycin | |||||
| Salmonella Kentucky | Amikacin, Ceftazidime, Aztreonam, Phosphomycin | ||||||
| Salmonella Typhimurium | Amikacin, Cephalothin, Cefazolin, Cefoxitin, Cefepime, Aztreonam, Phosphomycin | ||||||
| [31] | Mean number of antibiotics the strains were resistant to at 0 days of storage: | Mean number of antibiotics the strains were resistant to after 5 days of storage: | |||||
| TSP | Chicken legs containing E. coli were dipped in 12% TSP at 20 ± 1 °C for 15 min and subsequently refrigerated at 7 ± 1 °C and stored. Chicken legs dipped in tap water were used as a control. | Escherichia coli | Control: 3.76 ± 2.01 aa TSP: 3.80 ±2.48 aa | Control: 3.44 ± 1.42 aa TSP: 4.64 ± 2.64 bb | |||
| The mean numbers from the same day (different treatments) with no letters in common (superscript) are significantly different (P < 0.05). The mean numbers within the same treatment (day 0 versus day 5) with no letters in common (subscript) are significantly different (P < 0.05). | |||||||
| [30] | MIC fold change of Campylobacter strains relative to the pre-adapted strains. | ||||||
| TSP | Step-wise exposure to gradually increasing concentrations (2, 2.5, 3, 4 to 5 mg/mL, depending upon thegrowth of the adapted microorganism) of TSP over 15 days. | Campylobacter jejuni K49/4 | Days after repeated exposure to TSP | 5 | 10 | 15 | |
| MIC fold change | 2 | 2 | 2 | ||||
| Campylobacter jejuni NCTC11168 | Days after repeated exposure to TSP | 5 | 10 | 15 | |||
| MIC fold change | 2 | 0.5 | 2 | ||||
| Campylobacter jejuni ATCC33560 | Days after repeated exposure to TSP | 5 | 10 | 15 | |||
| MIC fold change | 1 | 1 | 0.125 | ||||
| Campylobacter coli 137 | Days after repeated exposure to TSP | 5 | 10 | 15 | |||
| MIC fold change | 1 | 0.008 | 0.004 | ||||
| Campylobacter coli ATCC33559 | Days after repeated exposure to TSP | 5 | 10 | 15 | |||
| MIC fold change | 2 | 2 | 1 | ||||
| Test | Type | Method | References |
|---|---|---|---|
| Direct | |||
| BioFinder | Qualitative | Direct observation of color change due to dying of biofilm components. | [62] |
| Contact plates | Quantitative | Sterile agar plate is placed on surface of interest and biofilm is detected via conventional culture methods. | [67] |
| Direct epifluorescence microscopy | Quantitative | Automatic cell quantification using computer software on digital images. | [68] |
| REALCO Biofilm Detection Kit | Qualitative | Direct observation of color change due to dying of biofilm components. | [63] |
| TBF® 300/ TBF® 300S | Qualitative | Direct observation of color change due to dying of biofilm components. | [64] |
| Indirect | |||
| BacTrac 4300 | Quantitative | Total viable counts calculated via impedance. | [69,70] |
| Plate count | Quantitative | Culture plating to determine the number of colony forming units (CFU). | [57] |
| TEMPO® | Quantitative | Cell counts from biofilms are calculated using most probable number (MPN) system based on fluorescence. | [65] |
| Abcam XTT tetrazolium salt and resazurin assay kit | Quantitative | Metabolic assays combined with spectrophotometry can be used to quantify biofilm. | [57,71,72] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cadena, M.; Kelman, T.; Marco, M.L.; Pitesky, M. Understanding Antimicrobial Resistance (AMR) Profiles of Salmonella Biofilm and Planktonic Bacteria Challenged with Disinfectants Commonly Used During Poultry Processing. Foods 2019, 8, 275. https://doi.org/10.3390/foods8070275
Cadena M, Kelman T, Marco ML, Pitesky M. Understanding Antimicrobial Resistance (AMR) Profiles of Salmonella Biofilm and Planktonic Bacteria Challenged with Disinfectants Commonly Used During Poultry Processing. Foods. 2019; 8(7):275. https://doi.org/10.3390/foods8070275
Chicago/Turabian StyleCadena, Myrna, Todd Kelman, Maria L. Marco, and Maurice Pitesky. 2019. "Understanding Antimicrobial Resistance (AMR) Profiles of Salmonella Biofilm and Planktonic Bacteria Challenged with Disinfectants Commonly Used During Poultry Processing" Foods 8, no. 7: 275. https://doi.org/10.3390/foods8070275