Maternal Consumption of a Diet Rich in Maillard Reaction Products Accelerates Neurodevelopment in F1 and Sex-Dependently Affects Behavioral Phenotype in F2 Rat Offspring

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diets
Quantification of Maillard Reaction Products in Feeds
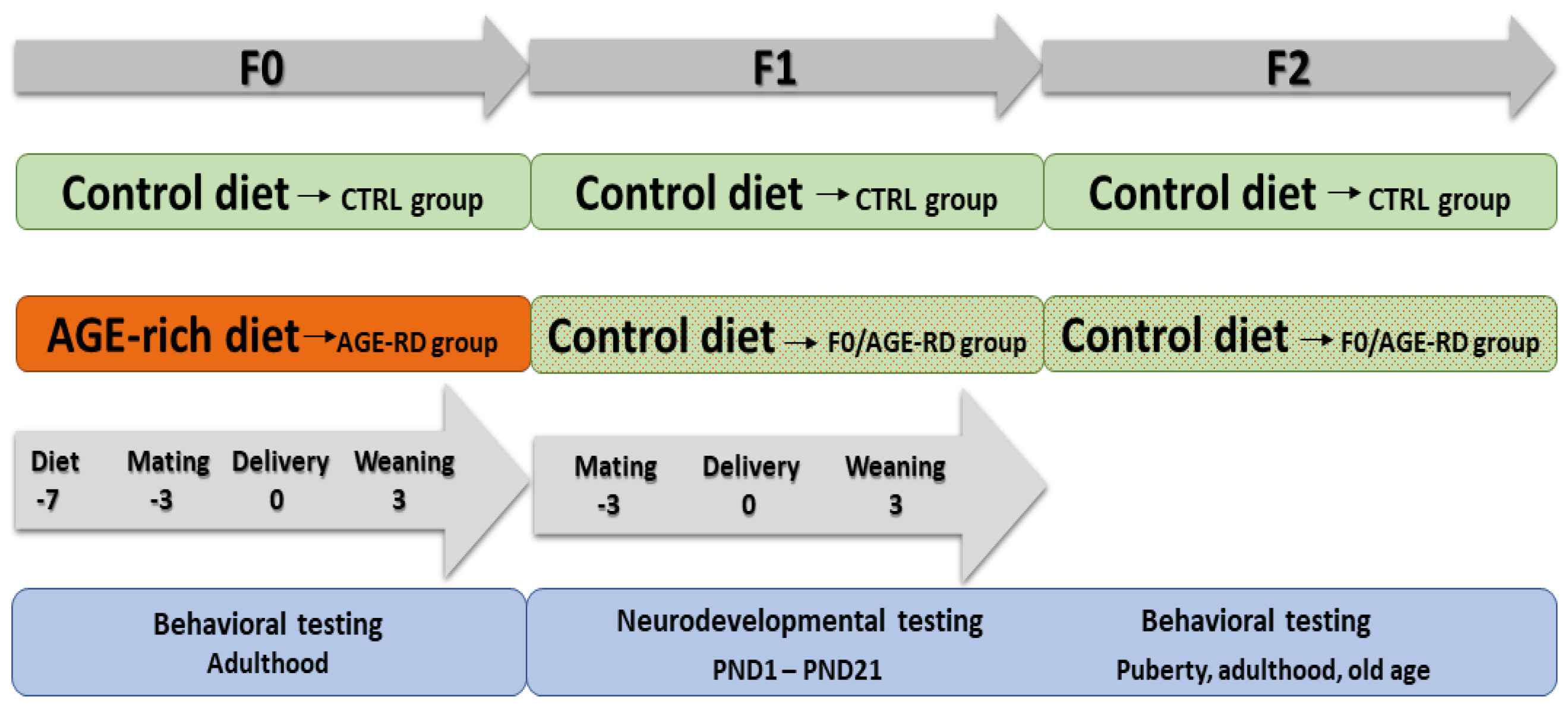
2.3. Experimental Design
2.4. Maternal Studies (F0)
2.4.1. Body Weight and Chow Consumption
2.4.2. Behavioral Tests
2.4.3. Sacrifice
2.5. Offspring Studies
2.5.1. Body Weight and Chow Consumption
2.5.2. Maturation of Physical Features and Development of Neurological Reflexes
2.5.3. Behavioral Tests
2.5.4. Sacrifice
2.6. Statistical analysis
3. Results
3.1. AGEs Content in Feed
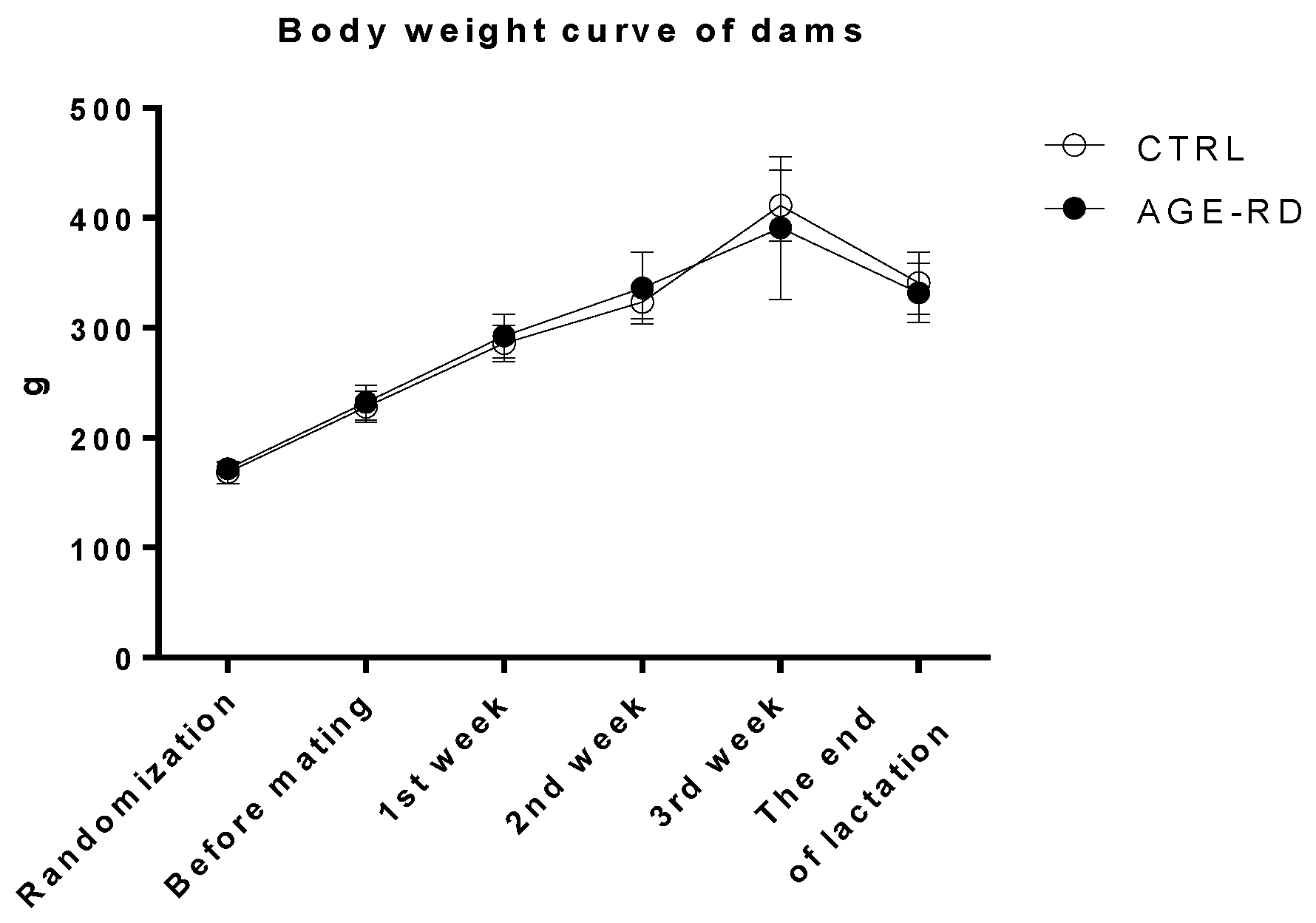
3.2. Maternal Data
3.3. Offspring
3.3.1. Body Weight and Chow Consumption
3.3.2. Effects of F0 Diet on Maturation of Physical Features and Development of Neurological Reflexes in F1 and F2 Offspring
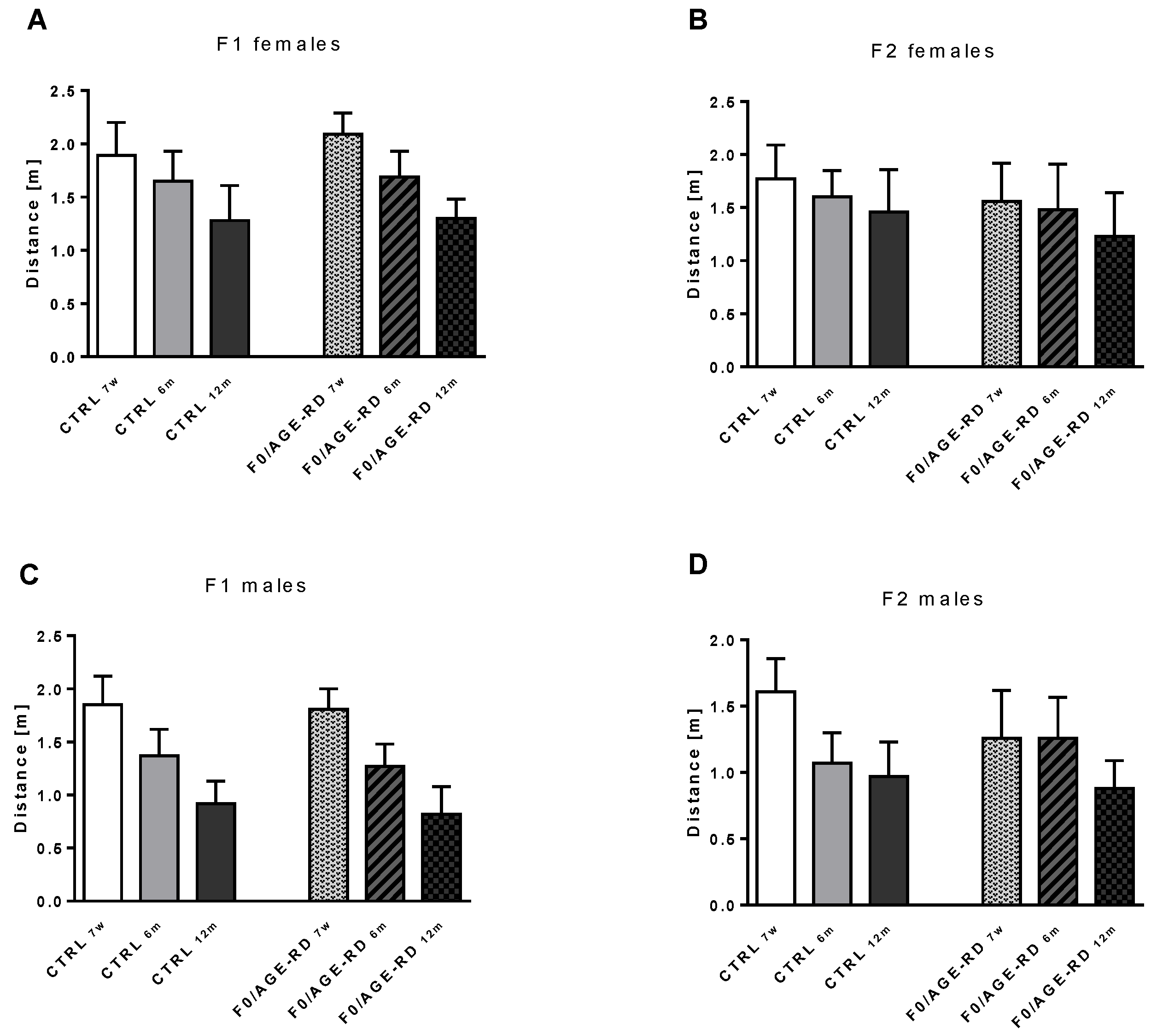
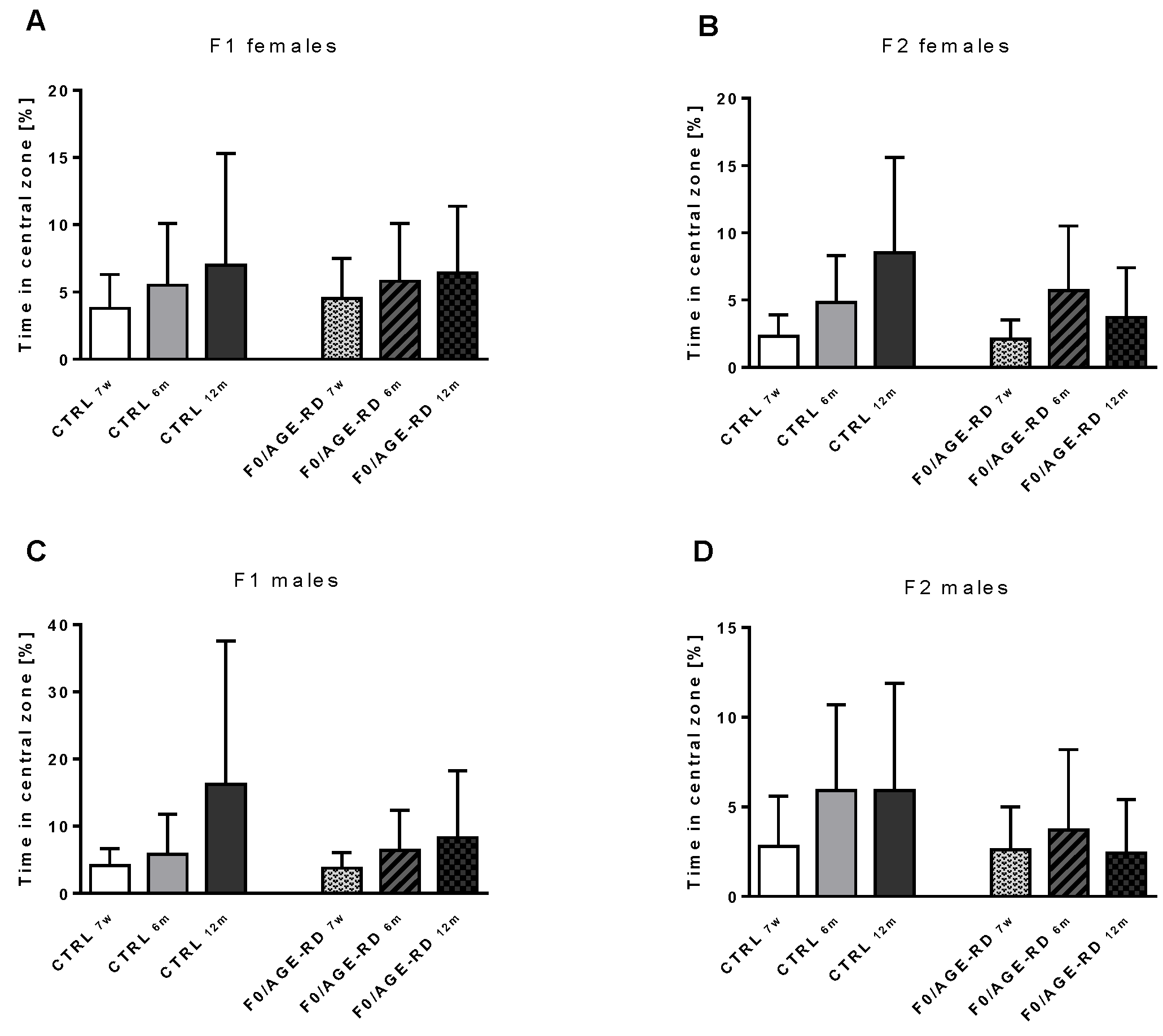
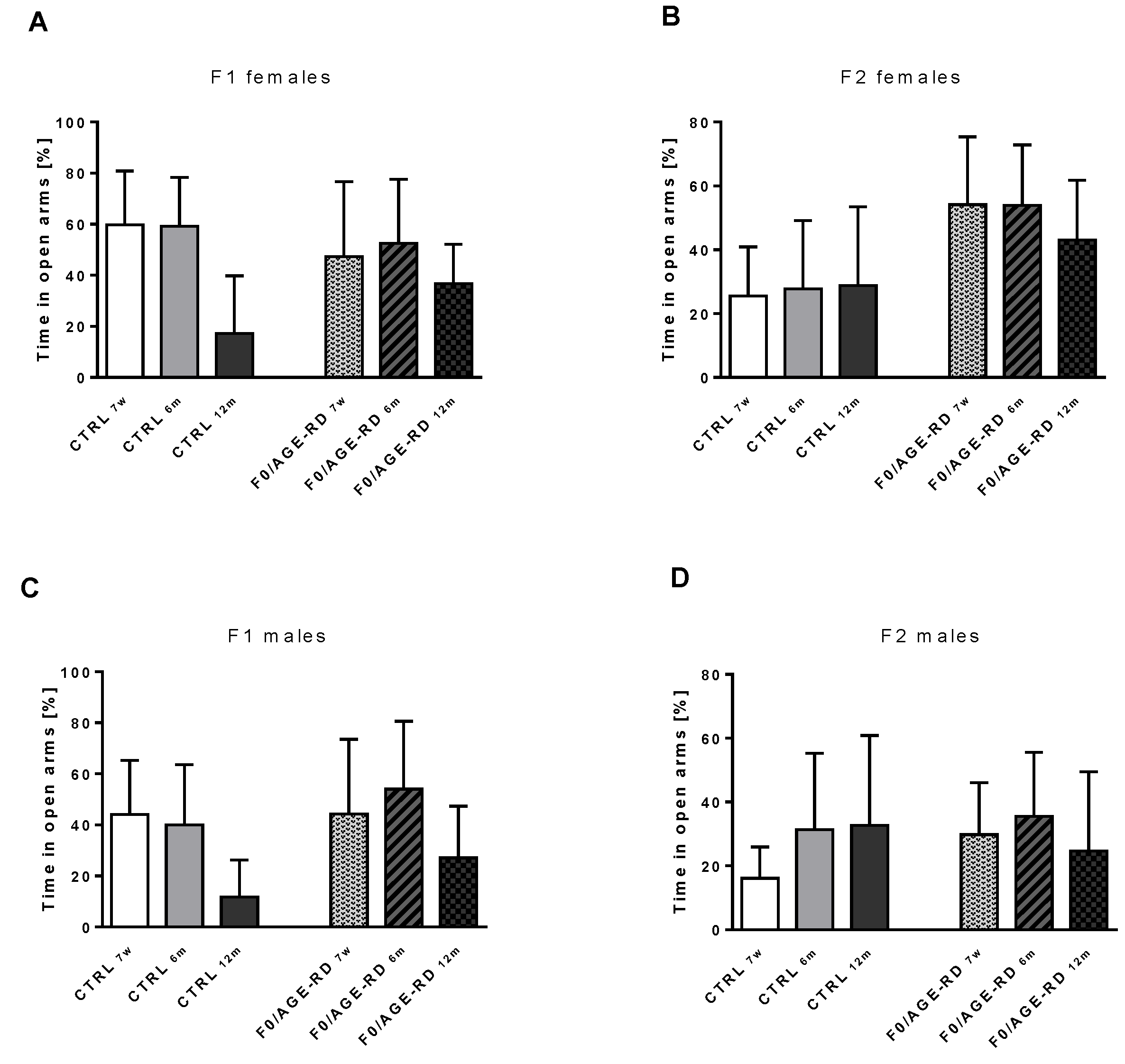
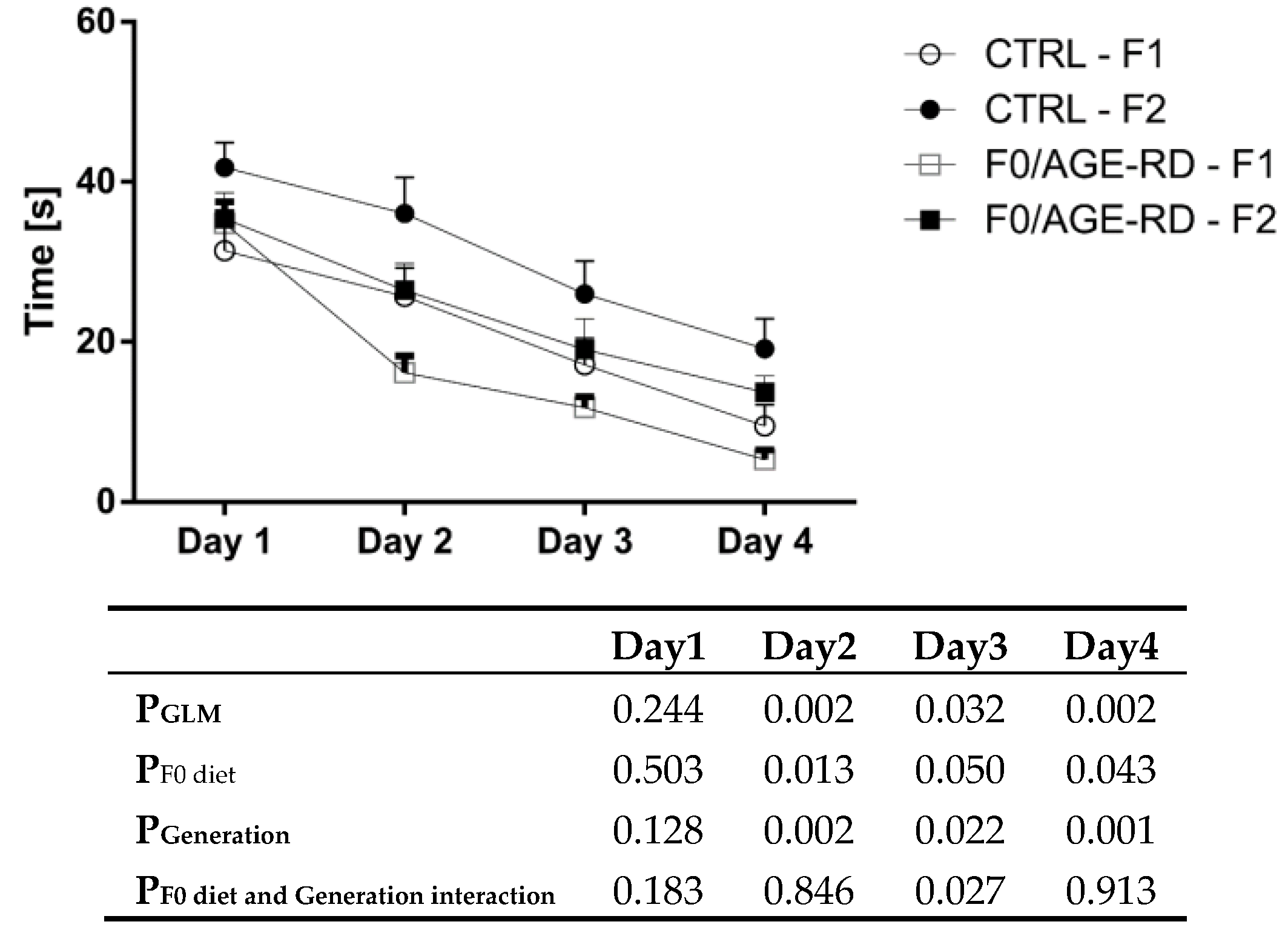
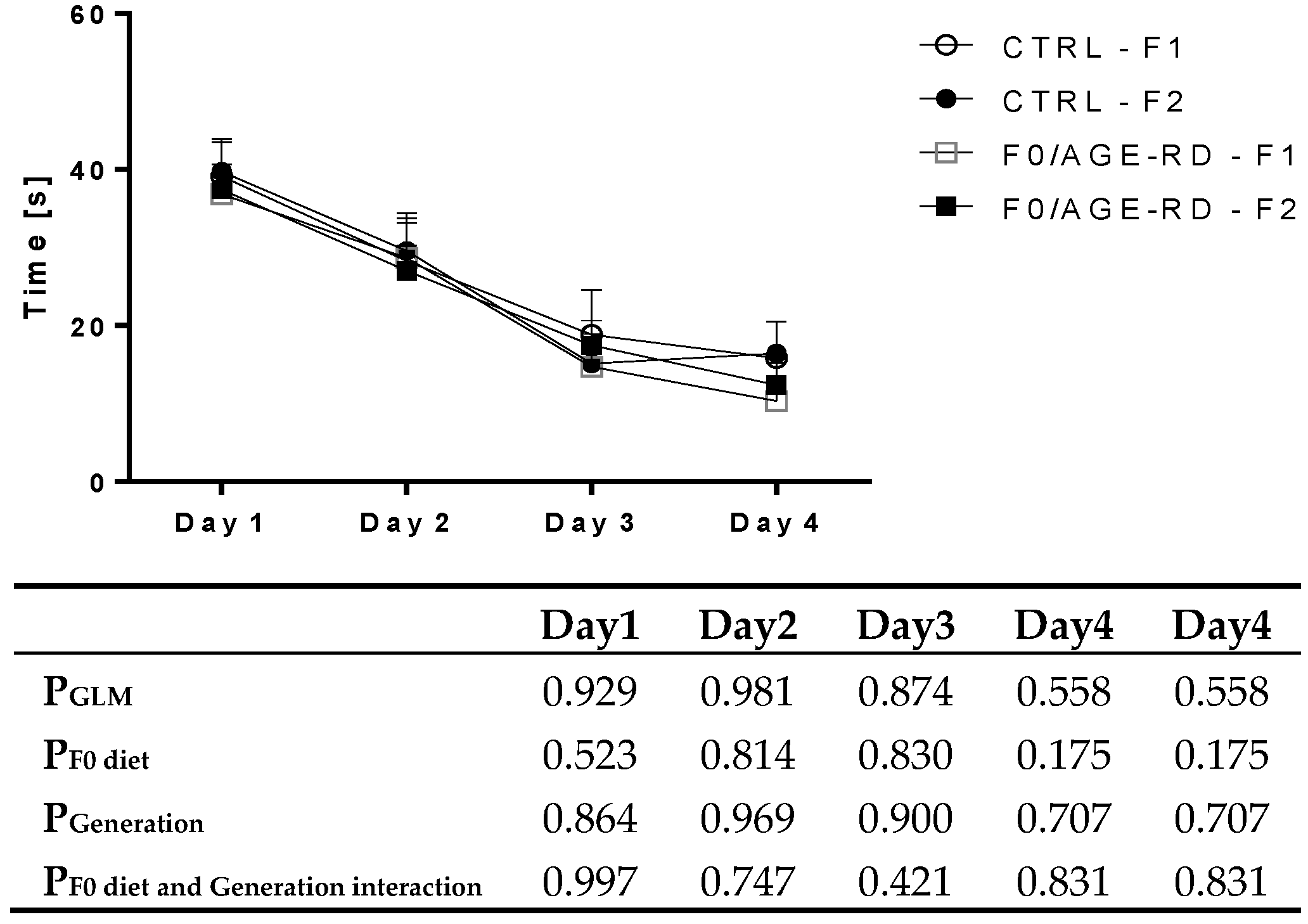
3.3.3. Effects of F0 Diet on Behavioral Phenotype of F1 and F2 Offspring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pellegrini, N.; Fogliano, V. Cooking, industrial processing and caloric density of foods. Curr. Opin. Food Sci. 2017, 14, 98–102. [Google Scholar] [CrossRef]
- Wrangham, R.W.; Conklin-Brittain, N.L. Cooking as a biological trait. Comp. Biochem. Physiol. Part A 2003, 136, 35–46. [Google Scholar] [CrossRef]
- Delgado-Andrade, C. New Knowledge in Analytical, Technological, and Biological Aspects of the Maillard Reaction. Foods 2017, 6, 40. [Google Scholar] [CrossRef]
- Maillard, L.C. Actions des acides amines sur les sucres. Formation des melanoides par voi methodique. C. R. Acad. Sci. Ser. 1912, 145, 66–68. [Google Scholar]
- Thornalley, P.J.; Langborg, A.; Minhas, H.S. Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochem. J. 1999, 344, 109–116. [Google Scholar] [CrossRef]
- Li, M.; Zeng, M.M.; He, Z.Y.; Zheng, Z.P.; Qin, F.; Tao, G.J.; Zhang, S.; Chen, J. Increased Accumulation of Protein-Bound N-epsilon-(Carboxymethyl)lysine in Tissues of Healthy Rats after Chronic Oral N-epsilon-(Carboxymethyl)lysine. J. Agric. Food Chem. 2015, 63, 1658–1663. [Google Scholar] [CrossRef]
- Kislinger, T.; Fu, C.F.; Huber, B.; Qu, W.; Taguchi, A.; Yan, S.D.; Hofmann, M.; Yan, S.F.; Pischetsrieder, M.; Stern, D.; et al. N-epsilon-(carboxymethyl)lysine adducts of proteins are ligands for receptor for advanced glycation end products that activate cell signaling pathways and modulate gene expression. J. Biol. Chem. 1999, 274, 31740–31749. [Google Scholar] [CrossRef]
- Xie, J.; Reverdatto, S.; Frolov, A.; Hoffmann, R.; Burz, D.S.; Shekhtman, A. Structural basis for pattern recognition by the receptor for advanced glycation end products (RAGE). J. Biol. Chem. 2008, 283, 27255–27269. [Google Scholar] [CrossRef]
- Bierhaus, A.; Humpert, P.M.; Morcos, M.; Wendt, T.; Chavakis, T.; Arnold, B.; Stern, D.M.; Nawroth, P.P. Understanding RAGE, the receptor for advanced glycation end products. J. Mol. Med. 2005, 83, 876–886. [Google Scholar] [CrossRef]
- Schmidt, A.M.; Yan, S.D.; Yan, S.F.; Stern, D.M. The biology of the receptor for advanced glycation end products and its ligands. Biochim. Biophys. Acta 2000, 1498, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Heidland, A.; Sebekova, K.; Schinzel, R. Advanced glycation end products and the progressive course of renal disease. Am. J. Kidney Dis. 2001, 38, S100–S106. [Google Scholar] [CrossRef]
- Goh, S.Y.; Cooper, M.E. The role of advanced glycation end products in progression and complications of diabetes. J. Clin. Endocrinol. Metab. 2008, 93, 1143–1152. [Google Scholar] [CrossRef]
- Deluyker, D.; Evens, L.; Bito, V. Advanced glycation end products (AGEs) and cardiovascular dysfunction: Focus on high molecular weight AGEs. Amino Acids 2017, 49, 1535–1541. [Google Scholar] [CrossRef]
- Reynaert, N.L.; Gopal, P.; Rutten, E.P.A.; Wouters, E.F.M.; Schalkwijk, C.G. Advanced glycation end products and their receptor in age-related, non-communicable chronic inflammatory diseases; Overview of clinical evidence and potential contributions to disease. Int. J. Biochem. Cell Biol. 2016, 81, 403–418. [Google Scholar] [CrossRef]
- Munch, G.; Westcott, B.; Menini, T.; Gugliucci, A. Advanced glycation endproducts and their pathogenic roles in neurological disorders. Amino Acids 2012, 42, 1221–1236. [Google Scholar] [CrossRef]
- Yamagishi, S.I.; Matsui, T. Role of Hyperglycemia-Induced Advanced Glycation End Product (AGE) Accumulation in Atherosclerosis. Ann. Vasc. Dis. 2018, 11, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Khan, H.; Siddiqui, Z.; Khan, M.Y.; Rehman, S.; Shahab, U.; Godovikova, T.; Silnikov, V.; Moinuddin. AGEs, RAGEs and s-RAGE; friend or foe for cancer. Semin. Cancer Biol. 2018, 49, 44–55. [Google Scholar] [CrossRef]
- Lund, M.N.; Ray, C.A. Control of Maillard Reactions in Foods: Strategies and Chemical Mechanisms. J. Agric. Food Chem. 2017, 65, 4537–4552. [Google Scholar] [CrossRef]
- Finot, P.A.; Magnenat, E. Metabolic transit of early and advanced Maillard products. Prog. Food Nutr. Sci. 1981, 5, 193–207. [Google Scholar]
- Seiquer, I.; Diaz-Alguacil, J.; Delgado-Andrade, C.; Lopez-Frias, M.; Hoyos, A.M.; Galdo, G.; Navarro, M.P. Diets rich in Maillard reaction products affect protein digestibility in adolescent males aged 11–14 y. Am. J. Clin. Nutr. 2006, 83, 1082–1088. [Google Scholar] [CrossRef]
- Alamir, I.; Niquet-Leridon, C.; Jacolot, P.; Rodriguez, C.; Orosco, M.; Anton, P.M.; Tessier, F.J. Digestibility of extruded proteins and metabolic transit of N epsilon-carboxymethyllysine in rats. Amino Acids 2013, 44, 1441–1449. [Google Scholar] [CrossRef]
- Grunwald, S.; Krause, R.; Bruch, M.; Henle, T.; Brandsch, M. Transepithelial flux of early and advanced glycation compounds across CaCO2 cell monolayers and their interaction with intestinal amino acid and peptide transport systems. Br. J. Nutr. 2006, 95, 1221–1228. [Google Scholar] [CrossRef]
- Hellwig, M.; Geissler, S.; Matthes, R.; Peto, A.; Silow, C.; Brandsch, M.; Henle, T. Transport of Free and Peptide-Bound Glycated Amino Acids: Synthesis, Transepithelial Flux at CaCO2 Cell Monolayers, and Interaction with Apical Membrane Transport Proteins. ChemBioChem 2011, 12, 1270–1279. [Google Scholar] [CrossRef]
- Somoza, V.; Wenzel, E.; Weiss, C.; Clawin-Radecker, I.; Grubel, N.; Erbersdobler, H.F. Dose-dependent utilisation of casein-linked lysinoalanine, N(epsilon)-fructoselysine and N(epsilon)-carboxymethyllysine in rats. Mol. Nutr. Food Res. 2006, 50, 833–841. [Google Scholar] [CrossRef]
- Sebekova, K.; Saavedra, G.; Zumpe, C.; Somoza, V.; Klenovicsova, K.; Birlouez-Aragon, I. Plasma concentration and urinary excretion of N-epsilon-(carboxymethyl)lysine in breast milk- and formula-fed infants. Ann. N. Y. Acad. Sci. 2008, 1126, 177–180. [Google Scholar] [CrossRef]
- Baye, E.; Kiriakova, V.; Uribarri, J.; Moran, L.J.; de Courten, B. Consumption of diets with low advanced glycation end products improves cardiometabolic parameters: Meta-analysis of randomised controlled trials. Sci. Rep. 2017, 7, 2266. [Google Scholar] [CrossRef]
- Roncero-Ramos, I.; Delgado-Andrade, C.; Tessier, F.J.; Niquet-Leridon, C.; Strauch, C.; Monnier, V.M.; Navarro, M.P. Metabolic transit of N(epsilon)-carboxymethyl-lysine after consumption of AGEs from bread crust. Food Funct. 2013, 4, 1032–1039. [Google Scholar] [CrossRef]
- Tessier, F.J.; Niquet-Leridon, C.; Jacolot, P.; Jouquand, C.; Genin, M.; Schmidt, A.M.; Grossin, N.; Boulanger, E. Quantitative assessment of organ distribution of dietary protein-bound C-13-labeled N-epsilon-carboxymethyllysine after a chronic oral exposure in mice. Mol. Nutr. Food Res. 2016, 60, 2446–2456. [Google Scholar] [CrossRef]
- Diamanti-Kandarakis, E.; Piperi, C.; Korkolopoulou, P.; Kandaraki, E.; Levidou, G.; Papalois, A.; Patsouris, E.; Papavassiliou, A.G. Accumulation of dietary glycotoxins in the reproductive system of normal female rats. J. Mol. Med. 2007, 85, 1413–1420. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J.P. The developmental origins of adult disease. Eur. J. Epidemiol. 2003, 18, 733–736. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Lillycrop, K.A.; Vickers, M.H.; Pleasants, A.B.; Phillips, E.S.; Beedle, A.S.; Burdge, G.C.; Hanson, M.A. Metabolic plasticity during mammalian development is directionally dependent on early nutritional status. Proc. Natl. Acad. Sci. USA 2007, 104, 12796–12800. [Google Scholar] [CrossRef] [Green Version]
- Gluckman, P.D.; Hanson, M.A. Developmental plasticity and human disease: Research directions. J. Intern. Med. 2007, 261, 461–471. [Google Scholar] [CrossRef]
- Gungor, N.K. Overweight and obesity in children and adolescents. J. Clin. Res. Pediatr. Endocrinol. 2014, 6, 129–143. [Google Scholar] [CrossRef]
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and Obesity: Prevalence, Consequences, and Causes of a Growing Public Health Problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef]
- Yeomans, M.R. Adverse effects of consuming high fat-sugar diets on cognition: Implications for understanding obesity. Proc. Nutr. Soc. 2017, 76, 455–465. [Google Scholar] [CrossRef]
- Mitanchez, D.; Chavatte-Palmer, P. Review shows that maternal obesity induces serious adverse neonatal effects and is associated with childhood obesity in their offspring. Acta Paediatr. 2018, 107, 1156–1165. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Reynolds, R.M.; Prescott, S.L.; Nyirenda, M.; Jaddoe, V.W.; Eriksson, J.G.; Broekman, B.F. Influence of maternal obesity on the long-term health of offspring. Lancet Diabetes Endocrinol. 2017, 5, 53–64. [Google Scholar] [CrossRef]
- Contu, L.; Hawkes, C.A. A Review of the Impact of Maternal Obesity on the Cognitive Function and Mental Health of the Offspring. Int. J. Mol. Sci. 2017, 18, 1093. [Google Scholar] [CrossRef]
- Edlow, A.G. Maternal obesity and neurodevelopmental and psychiatric disorders in offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef]
- Tozuka, Y.; Kumon, M.; Wada, E.; Onodera, M.; Mochizuki, H.; Wada, K. Maternal obesity impairs hippocampal BDNF production and spatial learning performance in young mouse offspring. Neurochem. Int. 2010, 57, 235–247. [Google Scholar] [CrossRef]
- Robb, J.L.; Messa, I.; Lui, E.; Yeung, D.; Thacker, J.; Satvat, E.; Mielke, J.G. A maternal diet high in saturated fat impairs offspring hippocampal function in a sex-specific manner. Behav. Brain Res. 2017, 326, 187–199. [Google Scholar] [CrossRef]
- Winther, G.; Elfving, B.; Muller, H.K.; Lund, S.; Wegener, G. Maternal High-fat Diet Programs Offspring Emotional Behavior in Adulthood. Neuroscience 2018, 388, 87–101. [Google Scholar] [CrossRef]
- Zieba, J.; Uddin, G.M.; Youngson, N.A.; Karl, T.; Morris, M.J. Long-term behavioural effects of maternal obesity in C57BL/6J mice. Physiol. Behav. 2019, 199, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.; Langley-Evans, S.C.; Voigt, J.P. The impact of maternal cafeteria diet on anxiety-related behaviour and exploration in the offspring. Physiol. Behav. 2011, 103, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.M.; King, M.V.; Davey, W.G.; Langley-Evans, S.C.; Voigt, J.P. Impact of cafeteria feeding during lactation in the rat on novel object discrimination in the offspring. Br. J. Nutr. 2014, 112, 1933–1937. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, A.; Batista, T.H.; Veronesi, V.B.; Giusti-Paiva, A.; Vilela, F.C. Cafeteria diet during the gestation period programs developmental and behavioral courses in the offspring. Int. J. Dev. Neurosci. 2018, 68, 45–52. [Google Scholar] [CrossRef]
- Mucellini, A.B.; Laureano, D.P.; Silveira, P.P.; Sanvitto, G.L. Maternal and post-natal obesity alters long-term memory and hippocampal molecular signaling of male rat. Brain Res. 2019, 1708, 138–145. [Google Scholar] [CrossRef]
- Clouard, C.; Kemp, B.; Val-Laillet, D.; Gerrits, W.J.J.; Bartels, A.C.; Bolhuis, J.E. Prenatal, but not early postnatal, exposure to a Western diet improves spatial memory of pigs later in life and is paired with changes in maternal prepartum blood lipid levels. FASEB J. 2016, 30, 2466–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Val-Laillet, D.; Besson, M.; Guerin, S.; Coquery, N.; Randuineau, G.; Kanzari, A.; Quesnel, H.; Bonhomme, N.; Bolhuis, J.E.; Kemp, B.; et al. A maternal Western diet during gestation and lactation modifies offspring’s microbiota activity, blood lipid levels, cognitive responses, and hippocampal neurogenesis in Yucatan pigs. FASEB J. 2017, 31, 2037–2049. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.A.; Javurek, A.B.; Painter, M.S.; Murphy, C.R.; Conard, C.M.; Gant, K.L.; Howald, E.C.; Ellersieck, M.R.; Wiedmeyer, C.E.; Vieira-Potter, V.J.; et al. Effects of a maternal high-fat diet on offspring behavioral and metabolic parameters in a rodent model. J. Dev. Orig. Health Dis. 2017, 8, 75–88. [Google Scholar] [CrossRef]
- Speight, A.; Davey, W.G.; McKenna, E.; Voigt, J.W. Exposure to a maternal cafeteria diet changes open-field behaviour in the developing offspring. Int. J. Dev. Neurosci. 2017, 57, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Clouard, C.; Gerrits, W.J.; Kemp, B.; Val-Laillet, D.; Bolhuis, J.E. Perinatal Exposure to a Diet High in Saturated Fat, Refined Sugar and Cholesterol Affects Behaviour, Growth, and Feed Intake in Weaned Piglets. PLoS ONE 2016, 11, e0154698. [Google Scholar] [CrossRef]
- Winther, G.; Eskelund, A.; Bay-Richter, C.; Elfving, B.; Muller, H.K.; Lund, S.; Wegener, G. Grandmaternal high-fat diet primed anxiety-like behaviour in the second-generation female offspring. Behav. Brain Res. 2019, 359, 47–55. [Google Scholar] [CrossRef]
- Sarker, G.; Peleg-Raibstein, D. Maternal Overnutrition Induces Long-Term Cognitive Deficits across Several Generations. Nutrients 2019, 11, 7. [Google Scholar] [CrossRef]
- Peppa, M.; Brem, H.; Ehrlich, P.; Zhang, J.G.; Cai, W.; Li, Z.; Croitoru, A.; Thung, S.; Vlassara, H. Adverse effects of dietary glycotoxins on wound healing in genetically diabetic mice. Diabetes 2003, 52, 2805–2813. [Google Scholar] [CrossRef]
- Troise, A.D.; Wiltafsky, M.; Fogliano, V.; Vitaglione, P. The quantification of free Amadori compounds and amino acids allows to model the bound Maillard reaction products formation in soybean products. Food Chem. 2018, 247, 29–38. [Google Scholar] [CrossRef]
- Krause, R.; Knoll, K.; Henle, T. Studies in the formation of furosine and pyridosine during acid hydrolysis of different Amadori products of lysine. Eur. Food Res. Technol. 2003, 216, 277–283. [Google Scholar] [CrossRef]
- Domonkos, E.; Borbelyova, V.; Csongova, M.; Bosy, M.; Kacmarova, M.; Ostatnikova, D.; Hodosy, J.; Celec, P. Sex differences and sex hormones in anxiety-like behavior of aging rats. Horm. Behav. 2017, 93, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Luine, V. Recognition memory tasks in neuroendocrine research. Behav. Brain Res. 2015, 285, 158–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalcin, I.; Aksu, F.; Belzung, C. Effects of desipramine and tramadol in a chronic mild stress model in mice are altered by yohimbine but not by pindolol. Eur. J. Pharmacol. 2005, 514, 165–174. [Google Scholar] [CrossRef]
- Horvath, G.; Reglodi, D.; Farkas, J.; Vadasz, G.; Mammel, B.; Kvarik, T.; Bodzai, G.; Kiss-Illes, B.; Farkas, D.; Matkovits, A.; et al. Perinatal positive and negative influences on the early neurobehavioral reflex and motor development. In Perinatal Programming of Neurodevelopment. Advances in Neurobiology; Antonelli, M., Ed.; Springer: New York, NY, USA, 2015; Volume 10, pp. 149–167. [Google Scholar]
- Hodosy, J.; Ostatnikova, D.; Riljak, V.; Myslivecek, J.; Celec, P. The effect of rapid and depot testosterone and estradiol on spatial performance in water maze. Cent. Eur. J. Biol. 2012, 7, 596–602. [Google Scholar] [CrossRef] [Green Version]
- Nursten, H.E. The Chemistry of Nonenzymic Browning. In The Maillard Reaction, Chemistry, Biology and Implications; Nursten, H.E., Ed.; RCS: Oxford, UK, 2005; pp. 5–30. [Google Scholar]
- Haucke, E.; Santos, A.N.; Simm, A.; Henning, C.; Glomb, M.A.; Gurke, J.; Schindler, M.; Fischer, B.; Santos, A.N. Accumulation of advanced glycation end products in the rabbit blastocyst under maternal diabetes. Reproduction 2014, 148, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Mericq, V.; Piccardo, C.; Cai, W.J.; Chen, X.; Zhu, L.; Striker, G.E.; Vlassara, H.; Uribarri, J. Maternally Transmitted and Food-Derived Glycotoxins A factor preconditioning the young to diabetes? Diabetes Care 2010, 33, 2232–2237. [Google Scholar] [CrossRef]
- Goldberg, T.; Cai, W.J.; Peppa, M.; Dardaine, V.; Baliga, B.S.; Uribarri, J.; Vlassara, H. Advanced glycoxidation end products in commonly consumed foods. J. Am. Diet. Assoc. 2004, 104, 1287–1291. [Google Scholar] [CrossRef]
- Uribarri, J.; Woodruff, S.; Goodman, S.; Cai, W.J.; Chen, X.; Pyzik, R.; Yong, A.; Striker, G.E.; Vlassara, H. Advanced Glycation End Products in Foods and a Practical Guide to Their Reduction in the Diet. J. Am. Diet. Assoc. 2010, 110, 911–916. [Google Scholar] [CrossRef]
- Poulsen, M.W.; Hedegaard, R.V.; Andersen, J.M.; de Courten, B.; Bugel, S.; Nielsen, J.; Skibsted, L.H.; Dragsted, L.O. Advanced glycation endproducts in food and their effects on health. Food Chem. Toxicol. 2013, 60, 10–37. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Fogliano, V. Dietary Advanced Glycosylation End-Products (dAGEs) and Melanoidins Formed through the Maillard Reaction: Physiological Consequences of their Intake. Ann. Rev. Food Sci. Technol. 2018, 9, 271–291. [Google Scholar] [CrossRef]
- Scheijen, J.L.J.M.; Clevers, E.; Engelen, L.; Dagnelie, P.C.; Brouns, F.; Stehouwer, C.D.A.; Schalkwijk, C.G. Analysis of advanced glycation endproducts in selected food items by ultra-performance liquid chromatography tandem mass spectrometry: Presentation of a dietary AGE database. Food Chem. 2016, 190, 1145–1150. [Google Scholar] [CrossRef]
- Birlouez-Aragon, I.; Pischetsrieder, M.; Leclere, J.; Morales, F.J.; Hasenkopf, K.; Kientsch-Engel, R.; Ducauze, C.J.; Rutledge, D. Assessment of protein glycation markers in infant formulas. Food Chem. 2004, 87, 253–259. [Google Scholar] [CrossRef] [Green Version]
- De Courten, B.; de Courten, M.P.J.; Soldatos, G.; Dougherty, S.L.; Straznicky, N.; Schlaich, M.; Sourris, K.C.; Chand, V.; Scheijen, J.L.J.M.; Kingwell, B.A.; et al. Diet low in advanced glycation end products increases insulin sensitivity in healthy overweight individuals: A double-blind, randomized, crossover trial. Am. J. Clin. Nutr. 2016, 103, 1426–1433. [Google Scholar] [CrossRef]
- Birlouez-Aragon, I.; Saavedra, G.; Tessier, F.J.; Galinier, A.; Ait-Ameur, L.; Lacoste, F.; Niamba, C.N.; Alt, N.; Somoza, V.; Lecerf, J.M. A diet based on high-heat-treated foods promotes risk factors for diabetes mellitus and cardiovascular diseases. Am. J. Clin. Nutr. 2010, 91, 1220–1226. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Andrade, C.; Tessier, F.J.; Niquet-Leridon, C.; Seiquer, I.; Pilar Navarro, M. Study of the urinary and faecal excretion of N (epsilon)-carboxymethyllysine in young human volunteers. Amino Acids 2012, 43, 595–602. [Google Scholar] [CrossRef]
- Sebekova, K.; Klenovics, K.S.; Boor, P.; Celec, P.; Behuliak, M.; Schieberle, P.; Heidland, A.; Palkovits, M.; Somoza, V. Behaviour and hormonal status in healthy rats on a diet rich in Maillard reaction products with or without solvent extractable aroma compounds. Physiol. Behav. 2012, 105, 693–701. [Google Scholar] [CrossRef]
- Mendes-da-Silva, C.; Giriko, C.A.; Mennitti, L.V.; Hosoume, L.F.; Souto, T.D.; da Silva, A.V. Maternal high-fat diet during pregnancy or lactation changes the somatic and neurological development of the offspring. Arq. Neuropsiquiatr. 2014, 72, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Csongova, M.; Gurecka, R.; Koborova, I.; Celec, P.; Domonkos, E.; Ulicna, O.; Somoza, V.; Sebekova, K. The effects of a maternal advanced glycation end product-rich diet on somatic features, reflex ontogeny and metabolic parameters of offspring mice. Food Funct. 2018, 9, 3432–3446. [Google Scholar] [CrossRef]
- Santillan, M.E.; Vincenti, L.M.; Martini, A.C.; de Cuneo, M.F.; Ruiz, R.D.; Mangeaud, A.; Stutz, G. Developmental and neurobehavioral effects of perinatal exposure to diets with different ω-6:ω-3 ratios in mice. Nutrition 2010, 26, 423–431. [Google Scholar] [CrossRef]
- Dierssen, M.; Fotaki, V.; de Lagran, M.M.; Gratacos, M.; Arbones, M.; Fillat, C.; Estivill, X. Neurobehavioral development of two mouse lines commonly used in transgenic studies. Pharmacol. Biochem. Behav. 2002, 73, 19–25. [Google Scholar] [CrossRef]
- Munch, G.; Thome, J.; Foley, P.; Schinzel, R.; Riederer, P. Advanced glycation endproducts in ageing and Alzheimer’s disease. Brain Res. Rev. 1997, 23, 134–143. [Google Scholar] [CrossRef]
- Peleg-Raibstein, D.; Luca, E.; Wolfrum, C. Maternal high-fat diet in mice programs emotional behavior in adulthood. Behav. Brain Res. 2012, 233, 398–404. [Google Scholar] [CrossRef]
- Sprott, R.L.; Eleftheriou, B.E. Open-field behavior in ageing inbred mice. Gerontologia 1974, 20, 155–162. [Google Scholar] [CrossRef]
- Meyza, K.Z.; Boguszewski, P.M.; Nikolaev, E.; Zagrodzka, J. Age increases anxiety and reactivity of the fear/anxiety circuit in Lewis rats. Behav. Brain Res. 2011, 225, 192–200. [Google Scholar] [CrossRef]
- Kanoski, S.E.; Davidson, T.L. Western diet consumption and cognitive impairment: Links to hippocampal dysfunction and obesity. Physiol. Behav. 2011, 103, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, I.; Ricny, J.; Atrakchi-Baranes, D.; Shemesh, C.; Kravitz, E.; Liraz-Zaltsman, S.; Maksin-Matveev, A.; Cooper, I.; Leibowitz, A.; Uribarri, J.; et al. High dietary advanced glycation end products are associated with poorer spatial learning and accelerated A deposition in an Alzheimer mouse model. Aging Cell 2016, 15, 309–316. [Google Scholar] [CrossRef]
- Madeira, M.D.; Lieberman, A.R. Sexual dimorphism in the mammalian limbic system. Prog. Neurobiol. 1995, 45, 275–333. [Google Scholar] [CrossRef]
- Srikanth, V.; Westcott, B.; Forbes, J.; Phan, T.G.; Beare, R.; Venn, A.; Pearson, S.; Greenaway, T.; Parameswaran, V.; Munch, G. Methylglyoxal, Cognitive Function and Cerebral Atrophy in Older People. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 68–73. [Google Scholar] [CrossRef]
- Beeri, M.S.; Moshier, E.; Schmeidler, J.; Godbold, J.; Uribarri, J.; Reddy, S.; Sano, M.; Grossman, H.T.; Cai, W.J.; Vlassara, H.; et al. Serum concentration of an inflammatory glycotoxin, methylglyoxal, is associated with increased cognitive decline in elderly individuals. Mech. Ageing Dev. 2011, 132, 583–587. [Google Scholar] [CrossRef]
- West, R.K.; Moshier, E.; Lubitz, I.; Schmeidler, J.; Godbold, J.; Cai, W.J.; Uribarri, J.; Vlassara, H.; Silverman, J.M.; Beeri, M.S. Dietary advanced glycation end products are associated with decline in memory in young elderly. Mech. Ageing Dev. 2014, 140, 10–12. [Google Scholar] [CrossRef] [Green Version]
- Degen, J.; Vogel, M.; Richter, D.; Hellwig, M.; Henle, T. Metabolic transit of dietary methylglyoxal. J. Agric. Food Chem. 2013, 61, 10253–10260. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Dicarbonyl stress in cell and tissue dysfunction contributing to ageing and disease. Biochem. Biophys. Res. Commun. 2015, 458, 221–226. [Google Scholar] [CrossRef] [Green Version]
- Southern, L.; Williams, J.; Esiri, M.M. Immunohistochemical study of N-epsilon-carboxymethyl lysine (CML) in human brain: Relation to vascular dementia. BMC Neurol. 2007, 7, 35. [Google Scholar] [CrossRef]
- Yaffe, K.; Lindquist, K.; Schwartz, A.V.; Vitartas, C.; Vittinghoff, E.; Satterfield, S.; Simonsick, E.M.; Launer, L.; Rosano, C.; Cauley, J.A.; et al. Advanced glycation end product level, diabetes, and accelerated cognitive aging. Neurology 2011, 77, 1351–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawangjit, A.; Oyanedel, C.N.; Niethard, N.; Salazar, C.; Born, J.; Inostroza, M. The hippocampus is crucial for forming non-hippocampal long-term memory during sleep. Nature 2018, 564, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Moreton, E.; Baron, P.; Tiplady, S.; McCall, S.; Clifford, B.; Langley-Evans, S.C.; Fone, K.C.F.; Voigt, J.P. Impact of early exposure to a cafeteria diet on prefrontal cortex monoamines and novel object recognition in adolescent rats. Behav. Brain Res. 2019, 363, 191–198. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTRL F | AGE-RD F | CTRL M | AGE-RD M | pDiet | pSex | pDiet and Sex intercation | |
|---|---|---|---|---|---|---|---|
| n = 13 | n = 16 | n = 11 | n = 16 | ||||
| Body weight (g) | |||||||
| Newborns | 7.0 ± 0.8 | 7.4 ± 0.8 | 7.2 ± 0.6 | 7.6 ± 0.9 | 0.070 | 0.301 | 0.867 |
| PND21 | 53.2 ± 8.9 | 57.6 ± 8.7 | 55.2 ± 8.0 | 60.4 ± 9.1 | 0.049 | 0.317 | 0.886 |
| 7 weeks | 209 ± 35 | 173 ± 28 | 293 ± 17 | 240 ± 56 | <0.001 | <0.001 | 0.467 |
| 6 months | 335 ± 38 | 318± 32 | 583 ± 70 | 551 ± 67 | 0.112 | <0.001 | 0.609 |
| 12 months | 402 ± 57 | 379 ± 42 | 743 ± 97 | 715 ± 109 | 0.278 | <0.001 | 0.914 |
| Fat pads/body weight (%) | 5.5 ± 2.1 | 4.3 ± 2.0 | 6.5 ± 2.5 | 5.1 ± 2.7 | 0.112 | 0.294 | 0.925 |
| Body weight gain (g/d) | 1.0 ± 0.2 | 0.9 ± 0.1 | 1.9 ± 0.3 | 1.8 ± 0.3 | 0.222 | <0.001 | 0.879 |
| Chow consumption-at week 7 (g) | 16.7 ± 4.5 | 17.9 ± 5.3 | 23.3 ± 5.8 | 16.9 ± 5.3 | 0.159 | 0.131 | 0.043 |
| Chow consumption-at 12 months (g) | 21.7 ± 1.1 | 27.3 ± 1.7 | 30.5 ± 0.8 | 31.2 ± 1.8 | <0.001 | <0.001 | <0.01 |
| CTRL F | AGE-RD F | CTRL M | AGE-RD M | pDie | pSex | pD*S | |
|---|---|---|---|---|---|---|---|
| n = 12 | n = 15 | n = 14 | n = 15 | ||||
| Body weight (g) | |||||||
| Newborns | 7.3 ± 1.0 | 7.4 ± 0.5 | 7.4 ± 0.8 | 7.5 ± 0.6 | 0.270 | 0.260 | 0.699 |
| PND21 | 50.8 ± 4.4 | 52.3 ± 8.3 | 55.6 ± 4.9 | 49.5 ± 4.9 | 0.174 | 0.566 | 0.024 |
| 7 weeks | 223 ± 14 | 235 ± 45 | 326 ± 22 | 292 ± 43 | 0.245 | <0.001 | 0.018 |
| 6 months | 330 ± 26.0 | 336 ± 85 | 592 ± 39 | 536 ± 78 | 0.159 | <0.001 | 0.084 |
| 12 months | 387 ± 30 | 386 ± 94 | 730 ± 56 | 668 ± 92 | 0.138 | <0.001 | 0.142 |
| Fat pads/body weight (%) | 5.3 ± 1.6 | 4.0 ± 1.6 | 4.8 ± 1.0 | 4.0 ± 1.8 | 0.061 | 0.609 | 0.710 |
| Body weight gain (g/d) | 1.0 ± 0.08 | 1.0 ± 0.2 | 1.9 ± 0.1 | 1.7 ± 0.2 | 0.146 | <0.001 | 0.166 |
| Chow consumption-at week 7 (g) | 21.3 ± 5.6 | 17.8 ± 4.1 | 26.5 ± 7.6 | 24.9 ± 7.3 | 0.263 | 0.010 | 0.673 |
| Chow consumption-at 12 months (g) | 22.4 ± 3.8 | 21.0 ± 3.4 | 32.2 ± 5.1 | 31.5 ± 5.6 | 0.614 | <0.001 | 0.867 |
| F1 Females | F1 Males | F2 Females | F2 Males | |
|---|---|---|---|---|
| Hindlimb placing | 23.4 | −7.5 | 25.3 | 25.3 |
| Forelimb grasp | 24.6 | 17.5 | 2.0 | 5.0 |
| Hindlimb grasp | 23.4 | 17.9 | 13.0 | 5.6 |
| Negative geotaxis | 11.2 | 47.6 | −10.1 | −4.9 |
| Surface righting | 31.7 | 27.8 | 31.0 | 14.4 |
| Gener/Sex | CTRL | AGE-RD | |||||
|---|---|---|---|---|---|---|---|
| 7w | 6M | 12M | 7w | 6M | 12M | ||
| F1/F | 5.0 ± 3.0 | 3.6 ± 4.3 | 3.1 ± 1.8 | 4.7 ± 2.9 | 3.8 ± 3.2 | 5.1 ± 1.6 | F(2, 85) = 13.1, p < 0.001; Age and diet interaction: NS; age, diet and generation interaction: NS |
| F2/F | 7.4 ± 4.5 | 4.0 ± 3.8 | 1.3 ± 1.7 | 7.0 ± 2.9 | 4.6 ± 3.1 | 2.7 ± 2.7 | |
| F1/M | 4.1 ± 2.2 | 3.8 ± 4.0 | 2.2 ± 1.8 | 5.4 ± 3.3 | 2.8 ± 3.4 | 3.7 ± 1.9 | F(2, 76) = 21.0, p < 0.001; Age and diet interaction: NS; age, diet and generation interaction: p = 0.042 |
| F2/M | 6.1 ± 2.3 | 3.7 ± 2.8 | 1.1 ± 1.3 | 3.7 ± 2.4 | 3.7 ± 1.9 | 0.9 ± 0.7 | |
| F1 | F2 | PGLM | |||
|---|---|---|---|---|---|
| CTRL | AGE-RD | CTRL | AGE-RD | ||
| Females | 18.7 ± 7.1 | 18.2 ± 4.0 | 14.8 ± 3.5 | 16.4 ± 4.1 | 0.187 |
| Males | 18.8 ± 2.4 | 20.1 ± 4.9 | 17.6 ± 4.9 | 16.2 ± 4.9 | 0.142 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csongová, M.; Renczés, E.; Šarayová, V.; Mihalovičová, L.; Janko, J.; Gurecká, R.; Troise, A.D.; Vitaglione, P.; Šebeková, K. Maternal Consumption of a Diet Rich in Maillard Reaction Products Accelerates Neurodevelopment in F1 and Sex-Dependently Affects Behavioral Phenotype in F2 Rat Offspring. Foods 2019, 8, 168. https://doi.org/10.3390/foods8050168
Csongová M, Renczés E, Šarayová V, Mihalovičová L, Janko J, Gurecká R, Troise AD, Vitaglione P, Šebeková K. Maternal Consumption of a Diet Rich in Maillard Reaction Products Accelerates Neurodevelopment in F1 and Sex-Dependently Affects Behavioral Phenotype in F2 Rat Offspring. Foods. 2019; 8(5):168. https://doi.org/10.3390/foods8050168
Chicago/Turabian StyleCsongová, Melinda, Emese Renczés, Veronika Šarayová, Lucia Mihalovičová, Jakub Janko, Radana Gurecká, Antonio Dario Troise, Paola Vitaglione, and Katarína Šebeková. 2019. "Maternal Consumption of a Diet Rich in Maillard Reaction Products Accelerates Neurodevelopment in F1 and Sex-Dependently Affects Behavioral Phenotype in F2 Rat Offspring" Foods 8, no. 5: 168. https://doi.org/10.3390/foods8050168