Antioxidant and Anti-Apoptotic Properties of Oat Bran Protein Hydrolysates in Stressed Hepatic Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Oat Bran Protein Hydrolysates
2.3. Cell Culture, Cytotoxicity and Cytoprotective Experiments
2.4. Determination of Intracellular Reactive Oxygen Species
2.5. Preparation of Cell Extracts for Glutathione and Enzyme Assays
2.6. Glutathione Assay
2.7. Antioxidant Enzyme Assays
2.7.1. Catalase
2.7.2. Glutathione Peroxidase
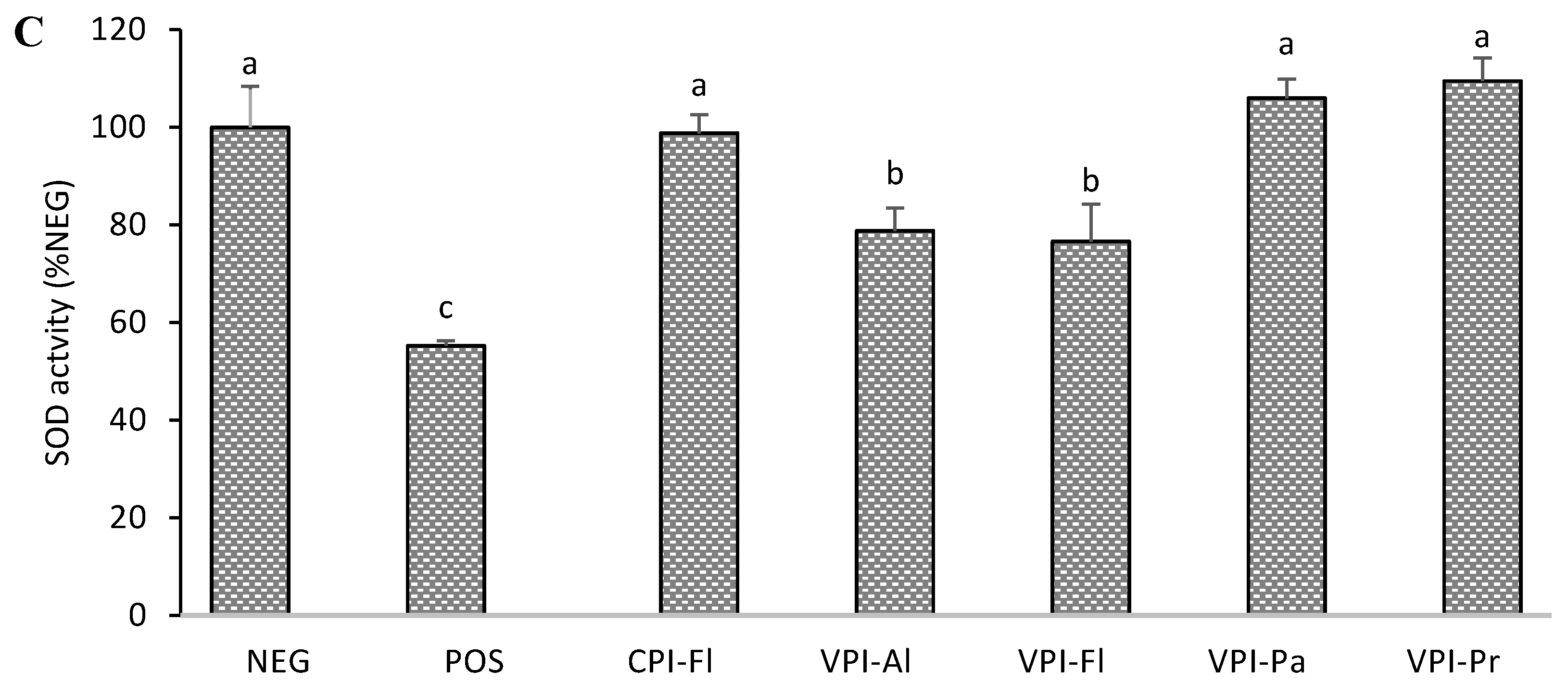
2.7.3. Superoxide Dismutase
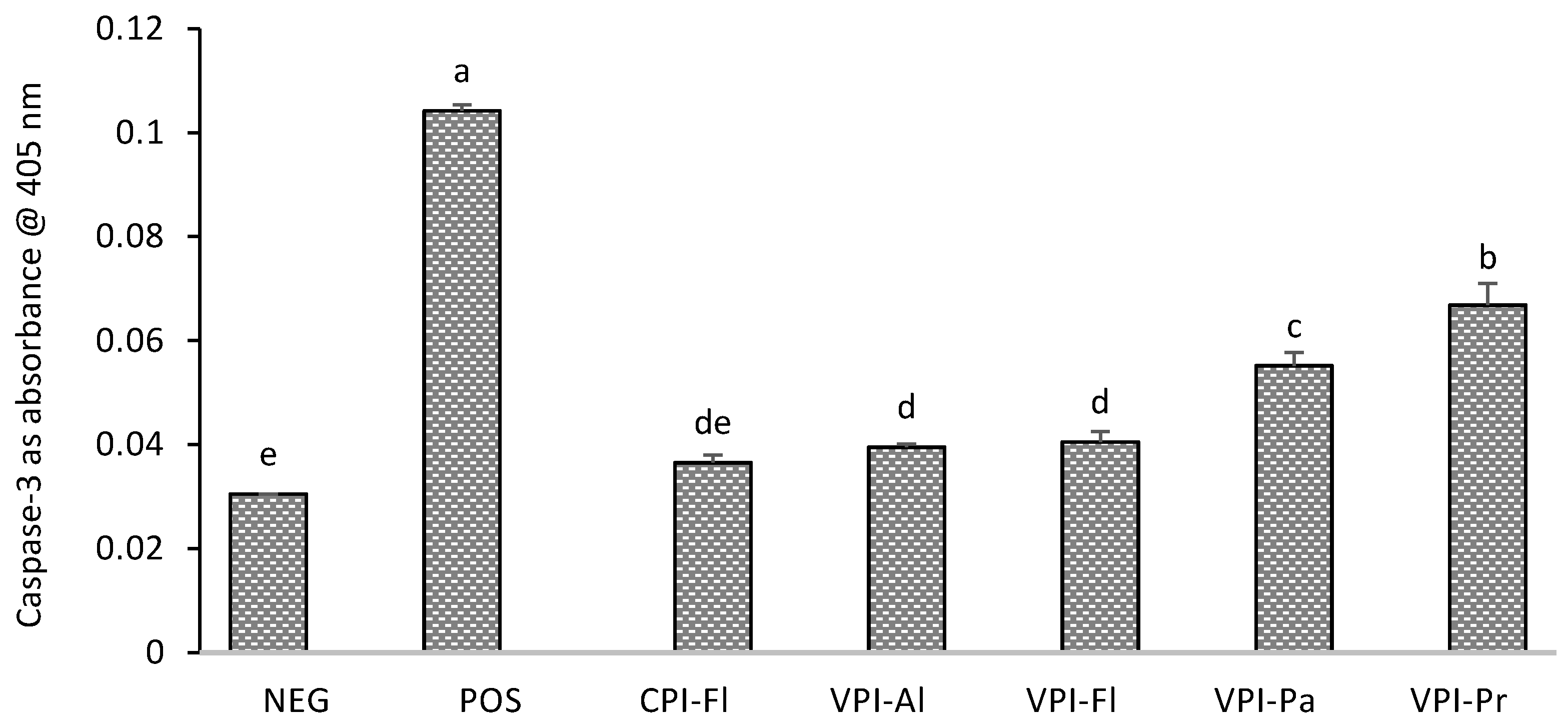
2.8. Caspase-3 Assay
2.9. Statistical Analysis
3. Results and Discussion
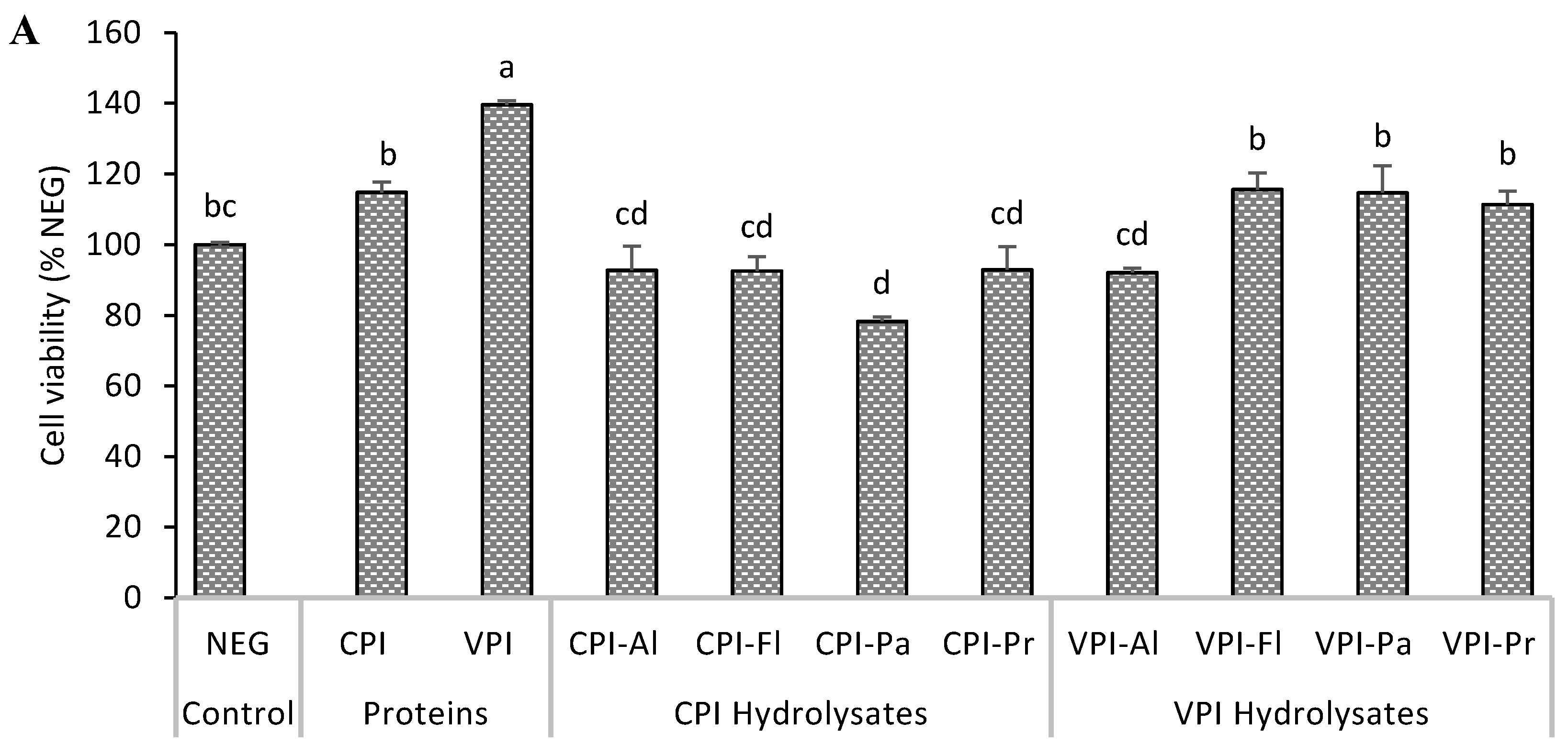
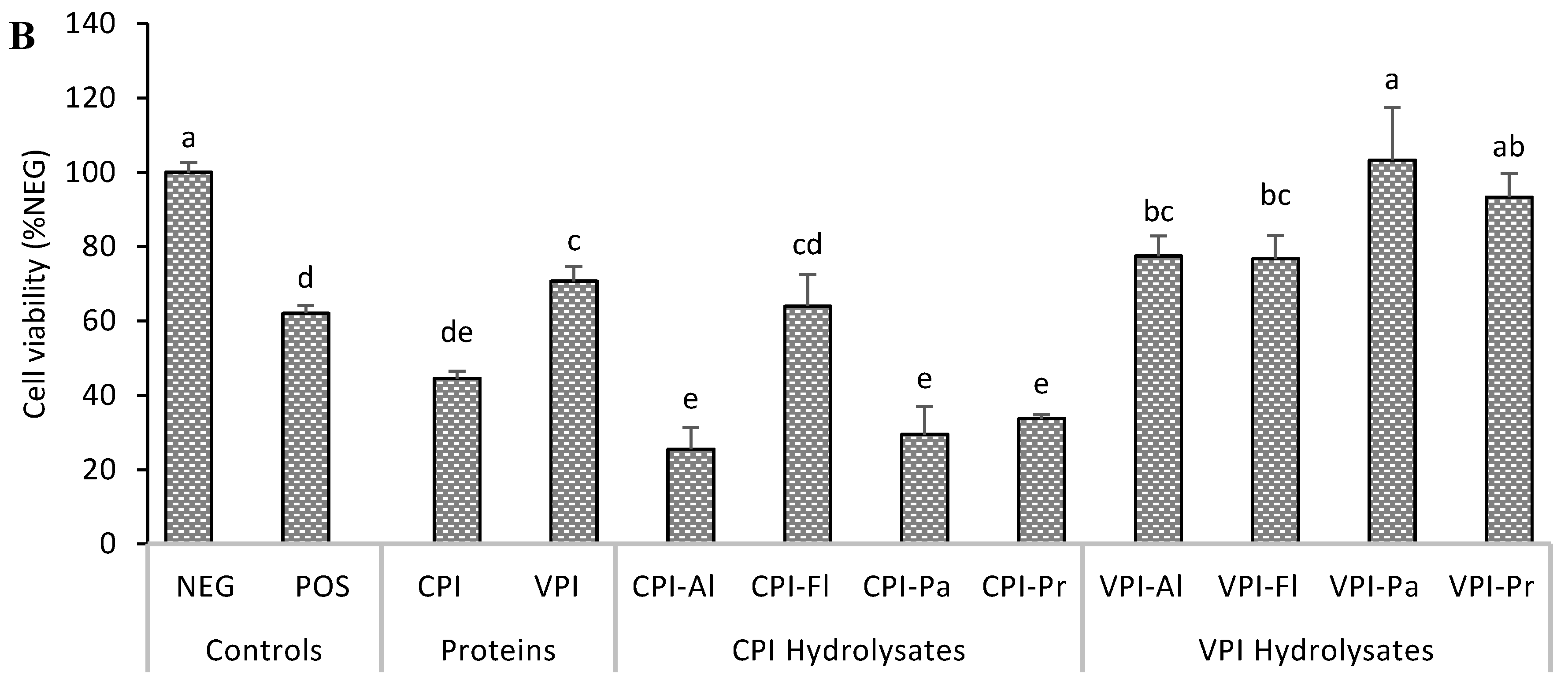
3.1. Cytotoxicity and Cytoprotection
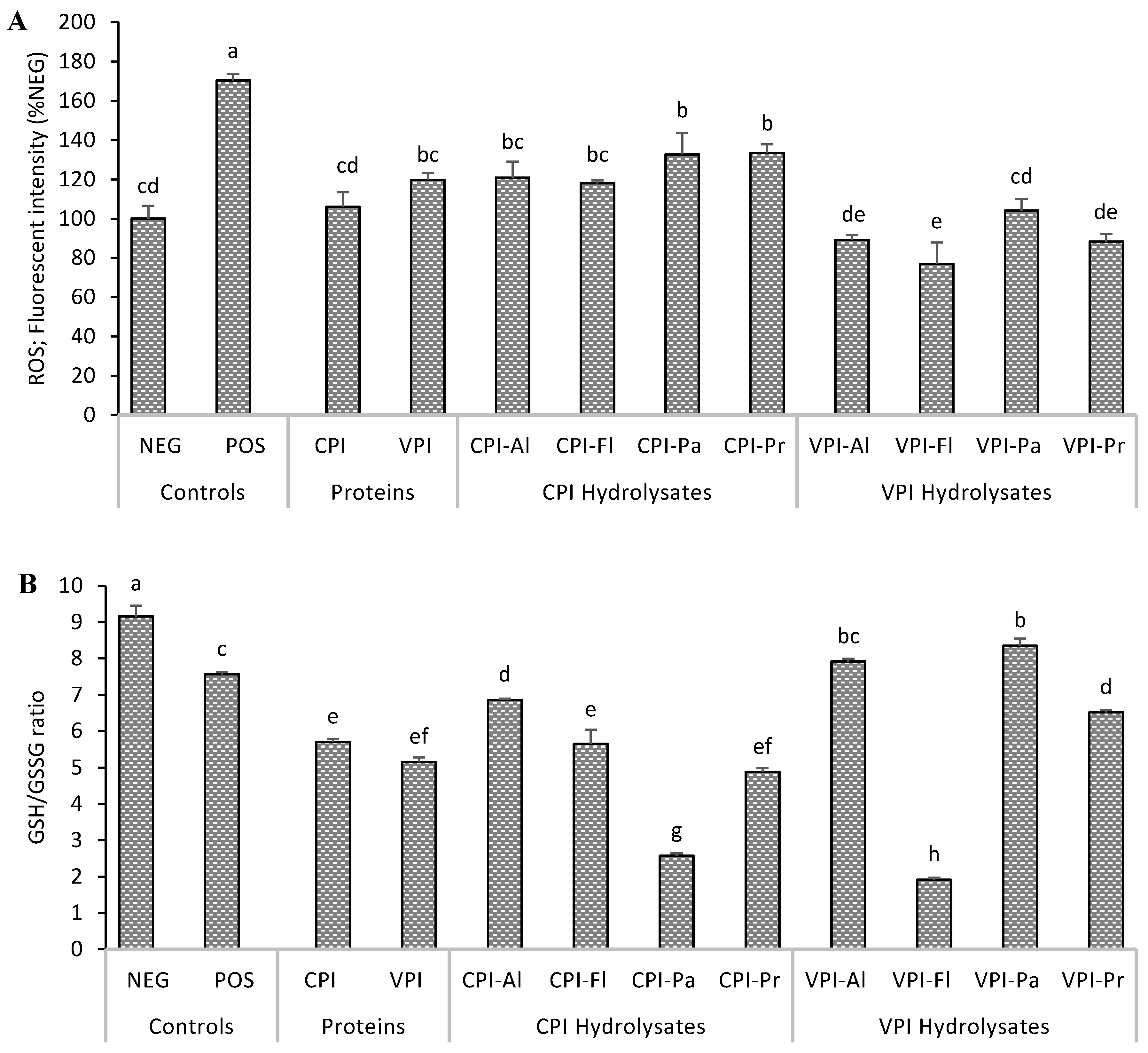
3.2. Determination of Intracellular Reactive Species and Glutathione
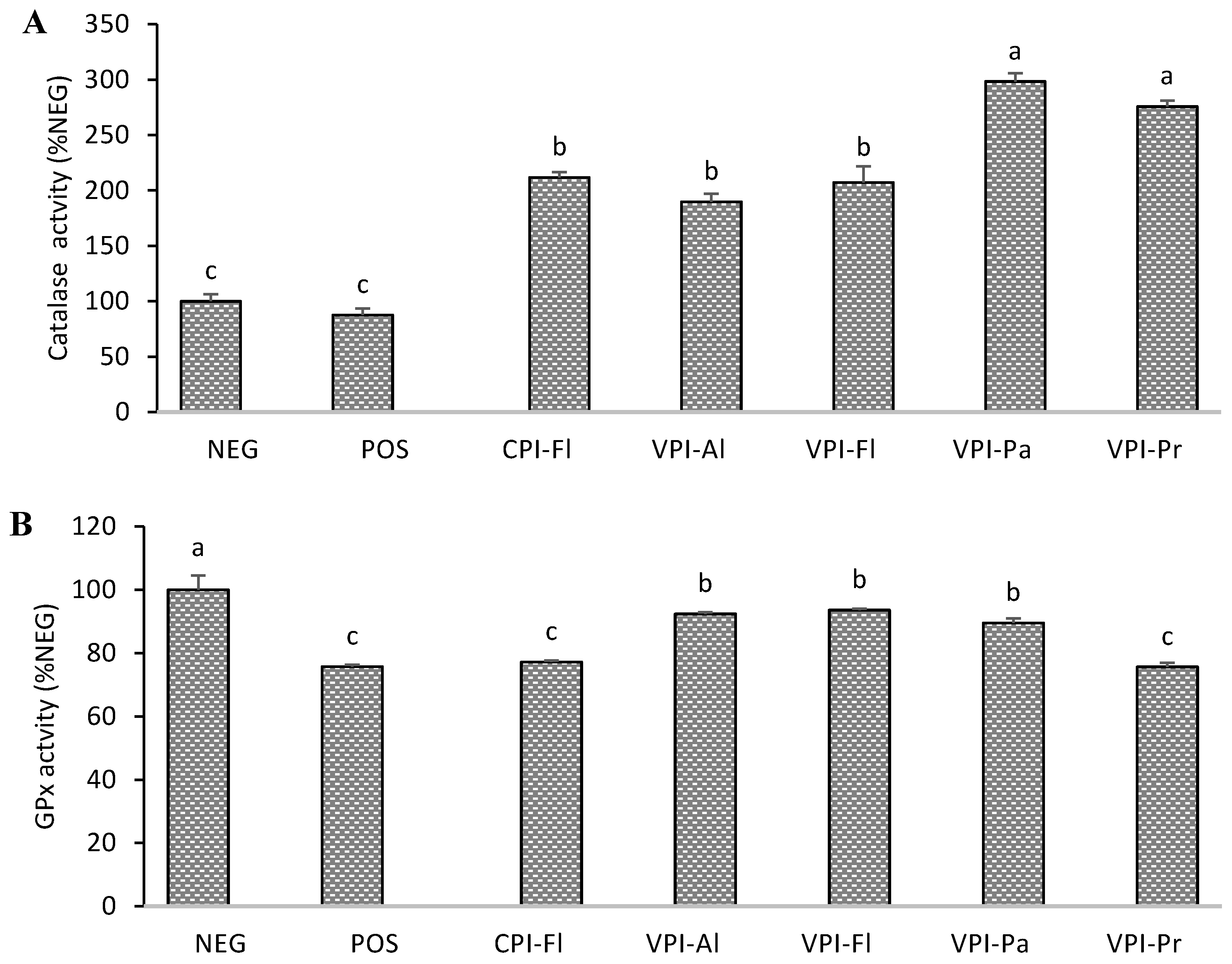
3.3. Antioxidant Enzymes Activity
3.4. Caspase-3 Determination and Cell Apoptosis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cichoz-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef]
- Leung, R.; Venus, C.; Zeng, T.; Tsopmo, A. Structure-function relationships of hydroxyl radical scavenging and chromium-VI reducing cysteine-tripeptides derived from rye secalin. Food Chem. 2018, 254, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Moronta, J.; Smaldini, P.L.; Docena, G.H.; Añón, M.C. Peptides of amaranth were targeted as containing sequences with potential anti-inflammatory properties. J. Funct. Foods 2016, 21, 463–473. [Google Scholar] [CrossRef]
- Yang, T.; Zhu, H.; Zhou, H.; Lin, Q.; Li, W.; Liu, J. Rice protein hydrolysate attenuates hydrogen peroxide induced apoptosis of myocardiocytes H9c2 through the Bcl-2/Bax pathway. Food Res. Int. 2012, 48, 736–741. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Cheung, F.; Hong, M.; Feng, Y. The potential and action mechanism of polyphenols in the treatment of liver diseases. Oxid. Med. Cell. Longev. 2018, 2018, 8394818. [Google Scholar] [CrossRef]
- Walters, M.E.; Esfandi, R.; Tsopmo, A. Potential of food hydrolyzed proteins and peptides to chelate iron or calcium and enhance their absorption. Foods 2018, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.-C.; Lv, J.; He, H.; Huang, W.; Han, Y. Hepatoprotective effects of corn peptides against carbon tetrachloride-induced liver injury in mice. J. Food Biochem. 2012, 36, 458–464. [Google Scholar] [CrossRef]
- Moritani, C.; Kawakami, K.; Fujita, A.; Kawakami, K.; Hatanaka, T.; Tsuboi, S. Anti-oxidative activity of hydrolysate from rice bran protein in HepG2 cells. Biol. Pharm. Bull. 2017, 40, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-H.; Qian, Z.-J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.-K. Antioxidant peptides from protein hydrolysate of microalgae Navicula incerta and their protective effects in Hepg2/CYP2E1 cells induced by ethanol. Phytother. Res. 2012, 26, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Tsopmo, A.; Cooper, A.; Jodayree, S. Enzymatic hydrolysis of oat flour protein isolates to enhance antioxidative properties. Adv. J. Food Sci. Technol. 2010, 2, 206–212. [Google Scholar]
- Vanvi, A.; Tsopmo, A. Pepsin digested oat bran proteins: separation, antioxidant activity, and identification of new peptides. J. Chem. 2016, 2016, 8216378. [Google Scholar] [CrossRef]
- Baakdah, M.M.; Tsopmo, A. Identification of peptides, metal binding and lipid peroxidation activities of HPLC fractions of hydrolyzed oat bran proteins. J. Food Sci. Technol. 2016, 53, 3593–3601. [Google Scholar] [CrossRef]
- Esfandi, R.; Willmore, W.G.; Tsopmo, A. Peptidomic analysis of hydrolyzed oat bran proteins, and their in vitro antioxidant and metal chelating properties. Food Chem. 2019, 279, 49–57. [Google Scholar] [CrossRef]
- Liu, Y.; Nair, M.G. An efficient and economical MTT assay for determining the antioxidant activity of plant. J. Nat. Prod. 2010, 73, 1193–1195. [Google Scholar] [CrossRef] [PubMed]
- Vieira, E.F.; da Silva, D.D.; Carmo, H.; Ferreira, I.M.P.L.V.O. Protective ability against oxidative stress of brewers’ spent grain protein hydrolysates. Food Chem. 2017, 228, 602–609. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Rui, H.L. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [PubMed]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Spitz, D.R.; Oberley, L.W. An assay for superoxide dismutase activity in mammalian tissue homogenates. Anal. Biochem. 1989, 179, 8–18. [Google Scholar] [CrossRef]
- Franek, F.; Hohenwarter, O.; Katinger, H. Plant protein hydrolysates: Preparation of defined peptide fractions promoting growth and production in animal cells cultures. Biotechnol. Prog. 2000, 16, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Karnjanapratum, S.; O’Callaghan, Y.C.; Benjakul, S.; O’Brien, N. Antioxidant, immunomodulatory and antiproliferative effects of gelatin hydrolysate from unicorn leatherjacket skin. J. Sci. Food Agric. 2016, 96, 3220–3226. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Esfandi, R.; Willmore, W.G.; Tsopmo, A. Antioxidant activity of oat proteins derived peptides in stressed hepatic hepg2 cells. Antioxidants 2016, 5, 39. [Google Scholar] [CrossRef]
- Yarnpakdee, S.; Benjakul, S.; Kristinsson, H.G.; Bakken, H.E. Preventive effect of Nile tilapia hydrolysate against oxidative damage of HepG2 cells and DNA mediated by H2O2 and AAPH. J. Food Sci. Technol. 2015, 52, 6194–6205. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Castilla, J.; Hernández-Álvarez, A.J.; Jiménez-Martínez, C.; Jacinto-Hernández, C.; Alaiz, M.; Girón-Calle, J.; Dávila-Ortiz, G. Antioxidant and metal chelating activities of Phaseolus vulgaris L. var. Jamapa protein isolates, phaseolin and lectin hydrolysates. Food Chem. 2012, 131, 1157–1164. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Aspects Med. 2009, 30, 1–12. [Google Scholar] [CrossRef]
- Tao, J.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Bioactive Peptides from Cartilage Protein Hydrolysate of Spotless Smoothhound and Their Antioxidant Activity In Vitro. Marine Drugs 2018, 16, 100. [Google Scholar] [CrossRef]
- Han, B.K.; Park, Y.; Choi, H.S.; Suh, H.J. Hepatoprotective effects of soluble rice protein in primary hepatocytes and in mice. J. Sci. Food Agric. 2016, 96, 685–694. [Google Scholar] [CrossRef]
- Phelan, M.; Aherne-Bruce, S.A.; O’Sullivan, D.; FitzGerald, R.J.; O’Brien, N.M. Potential bioactive effects of casein hydrolysates on human cultured cells. Int. Dairy J. 2009, 19, 279–285. [Google Scholar] [CrossRef]
- Meilhac, O.; Zhou, M.; Santanam, N.; Parthasarathy, S. Lipid peroxides induce expression of catalase in cultured vascular cells. J. Lipid Res. 2000, 41, 1205–1213. [Google Scholar]
- Homayouni-Tabrizi, M.; Asoodeh, A.; Soltani, M. Cytotoxic and antioxidant capacity of camel milk peptides: Effects of isolated peptide on superoxide dismutase and catalase gene expression. J. Food Drug Anal. 2017, 25, 567–575. [Google Scholar] [CrossRef]
- Richardson, D.R.; Ponka, P. Identification of a mechanism of iron uptake by cells which is stimulated by hydroxyl radicals generated via the iron-catalysed Haber-Weiss reaction. Biochim. Biophys. Acta 1995, 1269, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Bourre, J.-M.; Dumont, O.; Clément, M.; Dinh, L.; Droy-Lefaix, M.-T.; Christen, Y. Vitamin E deficiency has different effects on brain and liver phospholipid hydroperoxide glutathione peroxidase activities in the rat. Neurosci. Lett. 2000, 286, 87–90. [Google Scholar] [CrossRef]
- Harrington, H.A.; Ho, K.L.; Ghosh, S.; Tung, K. Construction and analysis of a modular model of caspase activation in apoptosis. Theor. Biol. Med. Model. 2008, 5, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.M.; Zhang, L.; Yu, H.X.; Meng, J.; Sun, Z.; Lu, R.R. Protective effect of whey protein hydrolysates on H2O2-induced PC12 cells oxidative stress via a mitochondria-mediated pathway. Food Chem. 2013, 141, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Ahn, C.B.; Yoon, N.Y.; Nam, K.H.; Kim, Y.K.; Je, J.Y. Protective effect of enzymatic hydrolysates from seahorse (Hippocampus abdominalis) against H2O2-mediated human umbilical vein endothelial cell injury. Biomed. Pharmacother. 2018, 108, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esfandi, R.; Willmore, W.G.; Tsopmo, A. Antioxidant and Anti-Apoptotic Properties of Oat Bran Protein Hydrolysates in Stressed Hepatic Cells. Foods 2019, 8, 160. https://doi.org/10.3390/foods8050160
Esfandi R, Willmore WG, Tsopmo A. Antioxidant and Anti-Apoptotic Properties of Oat Bran Protein Hydrolysates in Stressed Hepatic Cells. Foods. 2019; 8(5):160. https://doi.org/10.3390/foods8050160
Chicago/Turabian StyleEsfandi, Ramak, William G. Willmore, and Apollinaire Tsopmo. 2019. "Antioxidant and Anti-Apoptotic Properties of Oat Bran Protein Hydrolysates in Stressed Hepatic Cells" Foods 8, no. 5: 160. https://doi.org/10.3390/foods8050160