Optimization of a Microplate Assay for Generating Listeria Monocytogenes, E. Coli O157:H7, and Salmonella Biofilms and Enzymatic Recovery for Enumeration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Microplate Adherence Assay
2.2.1. Fluorescence Substrate for Fluorescence Microplate Assay
2.2.2. Microplates as a Substrate for Attachment and Biofilm Formation
2.2.3. Microplate Washing
2.3. Enzymatic Detachment of Adhered Cells from Microplates
2.3.1. Enzymes for Microbial Detachment
2.3.2. Enzymatic Detachment and Enumeration Assay
2.4. Statistical Analysis
3. Results
3.1. Choice of Fluorescent Substrate
3.2. Screening of Listeria monocytogenes, Salmonella, and E. coli O157:H7 via Fluorescence Microplate Assay
3.3. Buffer Washes with Microplate Biofilms
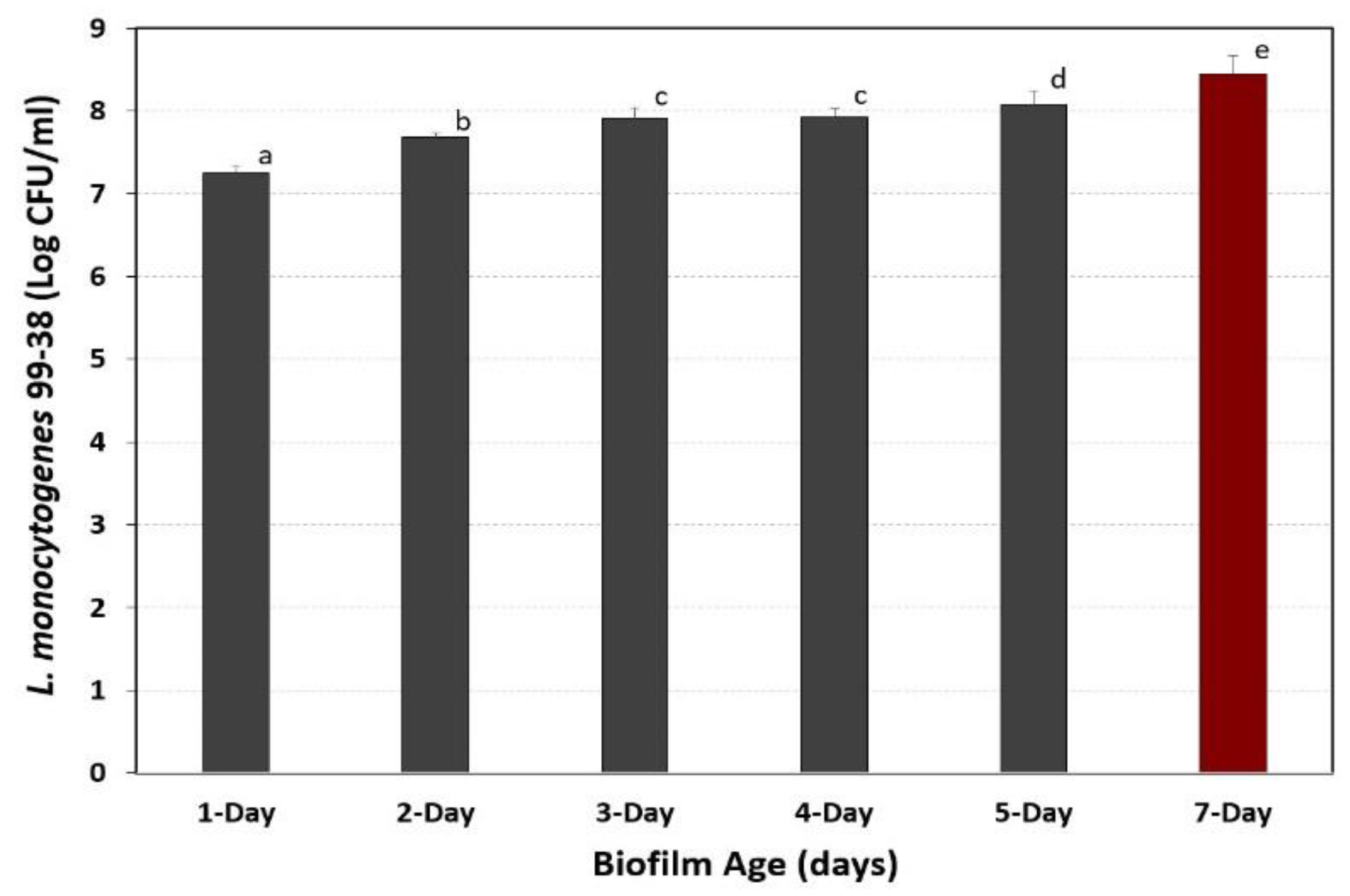
3.4. Development of Extended Biofilms in Microplates
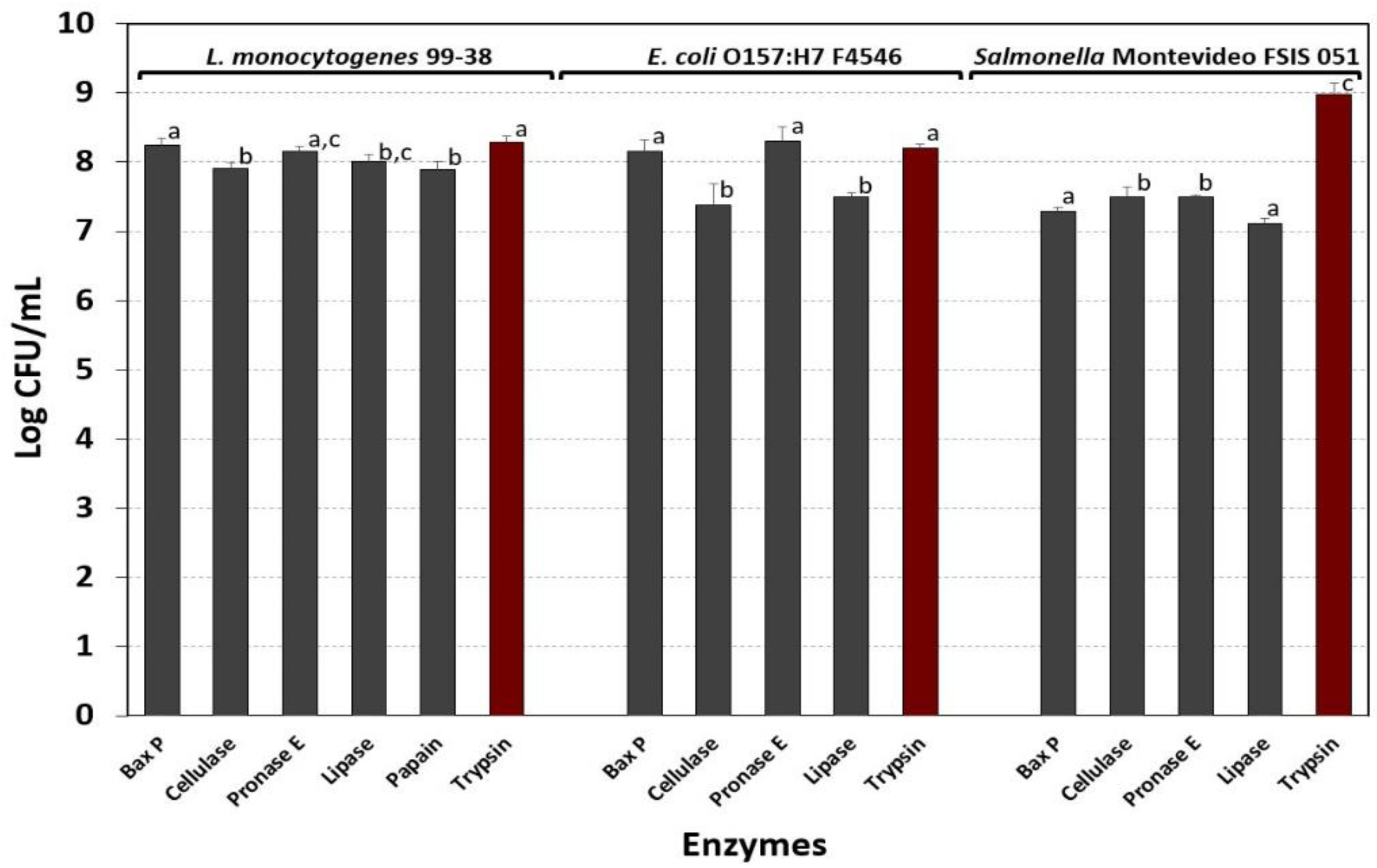
3.5. Evaluation of Various Enzymes for Bacterial Detachment and Enumeration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, X.; Zhao, F.; Wang, J.; Zhong, N. Biofilm formation and control strategies of foodborne pathogens: Food safety perspectives. RSC Adv. 2017, 7, 36670–36683. [Google Scholar] [CrossRef]
- Achinas, S.C.N.; Euverink, G.J.W. A brief recap of microbial adhesion and biofilms. Appl. Sci. 2019, 9, 2801. [Google Scholar] [CrossRef]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O’Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003, 49, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Hauck, D.; Hoffmann, M.; Sommer, R.; Joachim, I.; Müller, R.; Imberty, A.; Varrot, A.; Titz, A. Covalent lectin inhibition and application in bacterial biofilm imaging. Angew. Chem. Int. Ed. 2017, 56, 16559–16564. [Google Scholar] [CrossRef]
- Beloin, C.; Roux, A.; Ghigo, J.M. Escherichia coli biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 249–289. [Google Scholar]
- Kumar, C.G.; Anand, S.K. Significance of microbial biofilms in food industry: A review. Int. J. Food Microbiol. 1998, 42, 9–27. [Google Scholar] [CrossRef]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The Involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 1998, 280, 295–298. [Google Scholar] [CrossRef]
- Cao, S.; Wang, J.; Chen, H.; Chen, D. Progress of marine biofouling and antifouling technologies. Chin. Sci. Bull. 2011, 56, 598–612. [Google Scholar] [CrossRef]
- Hallam, N.B.; West, J.R.; Forster, C.F.; Simms, J. The potential for biofilm growth in water distribution systems. Water Res. 2001, 35, 4063–4071. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.D. Dental plaque as a biofilm and a microbial community—implications for health and disease. BMC Oral Health 2006, 6 (Suppl. 1), S14. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, R.A.N.; Frank, J.F. Biofilm formation and control in food processing facilities. Compr. Rev. Food Sci. Food Saf. 2003, 2, 22–32. [Google Scholar] [CrossRef]
- Lee Wong, A.C. Biofilms in food processing environments. J. Dairy Sci. 1998, 81, 2765–2770. [Google Scholar] [CrossRef]
- Alvarez-Ordóñez, A.; Coughlan, L.M.; Briandet, R.; Cotter, P.D. Biofilms in food processing environments: Challenges and opportunities. Annu. Rev. Food Sci. Technol. 2019, 10, 173–195. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005, 1, 1B.1.1–1B.1.17. [Google Scholar]
- Hartmann, R.; Singh, P.K.; Pearce, P.; Mok, R.; Song, B.; Díaz-Pascual, F.; Dunkel, J.; Drescher, K. Emergence of three-dimensional order and structure in growing biofilms. Nat. Phys. 2019, 15, 251–256. [Google Scholar] [CrossRef]
- Gamble, R.; Muriana, P.M. Microplate fluorescence assay for measurement of the ability of strains of Listeria monocytogenes from meat and meat-processing plants to adhere to abiotic surfaces. Appl. Environ. Microbiol. 2007, 73, 5235–5244. [Google Scholar] [CrossRef]
- Hood, S.K.; Zottola, E.A. Biofilms in food processing. Food Control 1995, 6, 9–18. [Google Scholar] [CrossRef]
- Kaplan, J.B. Biofilm dispersal: Mechanisms, clinical implications, and potential therapeutic uses. J. Dent. Res. 2010, 89, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Selan, L.; Berlutti, F.; Passariello, C.; Comodi-Ballanti, M.R.; Thaller, M.C. Proteolytic enzymes: A new treatment strategy for prosthetic infections? Antimicrob. Agents Chemother. 1993, 37, 2618–2621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ma, L.; Beuchat, L.R.; Erickson, M.C.; Phelan, V.H.; Doyle, M.P. Evaluation of treatments for elimination of foodborne pathogens on the surface of leaves and roots of lettuce (Lactuca sativa L.). J. Food Prot. 2009, 72, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, D.; Chen, J. Fate of various Salmonella enterica and enterohemorrhagic Escherichia coli cells attached to alfalfa, fenugreek, lettuce, and tomato seeds during germination. Food Control 2018, 88, 229–235. [Google Scholar] [CrossRef]
- Juneja, V.K.; Valenzuela Melendres, M.; Huang, L.; Gumudavelli, V.; Subbiah, J.; Thippareddi, H. Modeling the effect of temperature on growth of Salmonella in chicken. Food Microbiol. 2007, 24, 328–335. [Google Scholar] [CrossRef]
- Tiong, H.; Hartson, S.; Muriana, P.M. Comparison of surface proteomes of adherence variants of Listeria monocytogenes using LC-MS/MS for identification of potential surface adhesins. Pathogens 2016, 5, 40. [Google Scholar] [CrossRef]
- Kushwaha, K.; Muriana, P.M. Adherence Characteristics of Listeria Strains Isolated from Three Ready-to-Eat Meat Processing Plants. J. Food Prot. 2009, 72, 2125–2131. [Google Scholar] [CrossRef]
- Juneja, V.K.; Eblen, B.S.; Ransom, G.M. Thermal inactivation of Salmonella spp. in chicken broth, beef, pork, turkey, and chicken: Determination of D- and Z-values. J. Food Sci. 2001, 66, 146–152. [Google Scholar] [CrossRef]
- Alonso, E.P.C.; Gilliland, S.E.; Krehbiel, C.R. Incidence and toxin production ability of Escherichia coli O157:H7 isolated from cattle trucks. J. Food Prot. 2007, 70, 2383–2385. [Google Scholar] [CrossRef]
- Pericolini, E.; Colombari, B.; Ferretti, G.; Iseppi, R.; Ardizzoni, A.; Girardis, M.; Sala, A.; Peppoloni, S.; Blasi, E. Real-time monitoring of Pseudomonas aeruginosa biofilm formation on endotracheal tubes in vitro. BMC Microbiol. 2018, 18, 84. [Google Scholar] [CrossRef]
- Stepanović, S.; Ćirković, I.; Ranin, L.; Svabić-Vlahović, M. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 2004, 38, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Acta Pathol. Microbiol. Immunol. Scand. 2007, 115, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [PubMed]
- Borucki, M.K.; Peppin, J.D.; White, D.; Loge, F.; Call, D.R. Variation in biofilm formation among strains of Listeria monocytogenes. Appl. Environ. Microbiol. 2003, 69, 7336–7342. [Google Scholar] [CrossRef] [PubMed]
- Giaouris, E.; Chorianaopoulos, N.; Nychas, G.-J.E. Effect of temperature, pH, and water activity on biofilm formation by Salmonella enterica Enteritidis PT4 on stainless steel surfaces as indicated by the bead vortexing method and conductance measurements. J. Food Prot. 2005, 68, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Oulahal-Lagsir, N.; Martial-Gros, A.; Bonneau, M.; Blum, L.J. “Escherichia coli-milk” biofilm removal from stainless steel surfaces: Synergism between ultrasonic waves and enzymes. Biofouling 2003, 19, 159–168. [Google Scholar] [PubMed]
- Breeuwer, P.; Drocourt, J.L.; Bunschoten, N.; Zwietering, M.H.; Rombouts, F.M.; Abee, T. Characterization of uptake and hydrolysis of fluorescein diacetate and carboxyfluorescein diacetate by intracellular esterases in Saccharomyces cerevisiae, which result in accumulation of fluorescent product. Appl. Environ. Microbiol. 1995, 61, 1614–1619. [Google Scholar]
- Bunthof, C.J.; Bloemen, K.; Breeuwer, P.; Rombouts, F.M.; Abee, T. Flow cytometric assessment of viability of lactic acid bacteria. Appl. Environ. Microbiol. 2001, 67, 2326–2335. [Google Scholar] [CrossRef]
- Farinacci, M. Improved apoptosis detection in ovine neutrophils by annexin V and carboxyfluorescein diacetate staining. Cytotechnology 2007, 54, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.E.; Streger, S.H.; Rothmel, R.K.; Mailloux, B.J.; Hall, J.A.; Onstott, T.C.; Fredrickson, J.K.; Balkwill, D.L.; DeFlaun, M.F. Development of a vital fluorescent staining method for monitoring bacterial transport in subsurface environments. Appl. Environ. Microbiol. 2000, 66, 4486–4496. [Google Scholar] [CrossRef]
- Perez-Conesa, D.; McLandsborough, L.; Weiss, J. Inhibition and inactivation of Listeria monocytogenes and Escherichia coli O157:H7 colony biofilms by micellar-encapsulated eugenol and carvacrol. J. Food Prot. 2006, 69, 2947–2954. [Google Scholar] [CrossRef] [PubMed]
- Ostrov, I.P.T.; Shemesh, M. Robust biofilm-forming Bacillus isolates from the dairy environment demonstrate an enhanced resistance to cleaning-in-place procedures. Foods 2019, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, S.; Sandy, R.; Kota, K.; Lundh, L.; Gomba, G.; Recabo, K.; Duplantier, A.; Panchal, R.G. Robust biofilm assay for quantification and high throughput screening applications. J. Microbiol. Methods 2019, 159, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Phelan, K.; May, K.M. Basic techniques in mammalian cell tissue culture. Curr. Protoc. Cell Biol. 2015, 66, 1.1.1–1.1.22. [Google Scholar] [PubMed]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Nahar, S.; Mizan, M.F.R.; Ha, A.J.-W.; Ha, S.-D. Advances and future prospects of enzyme-based biofilm prevention approaches in the food industry. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1484–1502. [Google Scholar] [CrossRef]
- Daboor, S.M.; Raudonis, R.; Cohen, A.; Rohde, J.R.; Cheng, Z. Marine bacteria, a source for alginolytic enzyme to disrupt Pseudomonas aeruginosa biofilms. Mar. Drugs 2019, 17, 307. [Google Scholar] [CrossRef]
- Saggu, S.K.; Jha, G.; Mishra, P.C. Enzymatic degradation of biofilm by metalloprotease from Microbacterium sp. SKS10. Front. Bioeng. Biotechnol. 2019, 7, 192. [Google Scholar] [CrossRef]
- Lim, E.S.; Koo, O.K.; Kim, M.-J.; Kim, J.-S. Bio-enzymes for inhibition and elimination of Escherichia coli O157:H7 biofilm and their synergistic effect with sodium hypochlorite. Sci. Rep. 2019, 9, 9920. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aryal, M.; Pranatharthiharan, P.; Muriana, P.M. Optimization of a Microplate Assay for Generating Listeria Monocytogenes, E. Coli O157:H7, and Salmonella Biofilms and Enzymatic Recovery for Enumeration. Foods 2019, 8, 541. https://doi.org/10.3390/foods8110541
Aryal M, Pranatharthiharan P, Muriana PM. Optimization of a Microplate Assay for Generating Listeria Monocytogenes, E. Coli O157:H7, and Salmonella Biofilms and Enzymatic Recovery for Enumeration. Foods. 2019; 8(11):541. https://doi.org/10.3390/foods8110541
Chicago/Turabian StyleAryal, Manish, Preetty Pranatharthiharan, and Peter M. Muriana. 2019. "Optimization of a Microplate Assay for Generating Listeria Monocytogenes, E. Coli O157:H7, and Salmonella Biofilms and Enzymatic Recovery for Enumeration" Foods 8, no. 11: 541. https://doi.org/10.3390/foods8110541