Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor)

 , ,
, , 
Abstract
:1. Introduction
2. Experimental Procedure
2.1. Yellow Mealworm Protein Extract
2.2. Patient Sera
2.3. Dot-Blot Screening
2.4. SDS-PAGE and Western Blot Experiments
2.5. Degranulation Test
2.6. Digestion and Nano-LC-MS/MS Analysis
2.7. Bioinformatics
3. Results
3.1. Dot-Blot Screening of Shrimp-Allergic Patient Sera Towards a Yellow Mealworm Protein Extract
3.2. Identification of IgE-Binding Cross-Reactive Allergens in the Yellow Mealworm Protein Extract
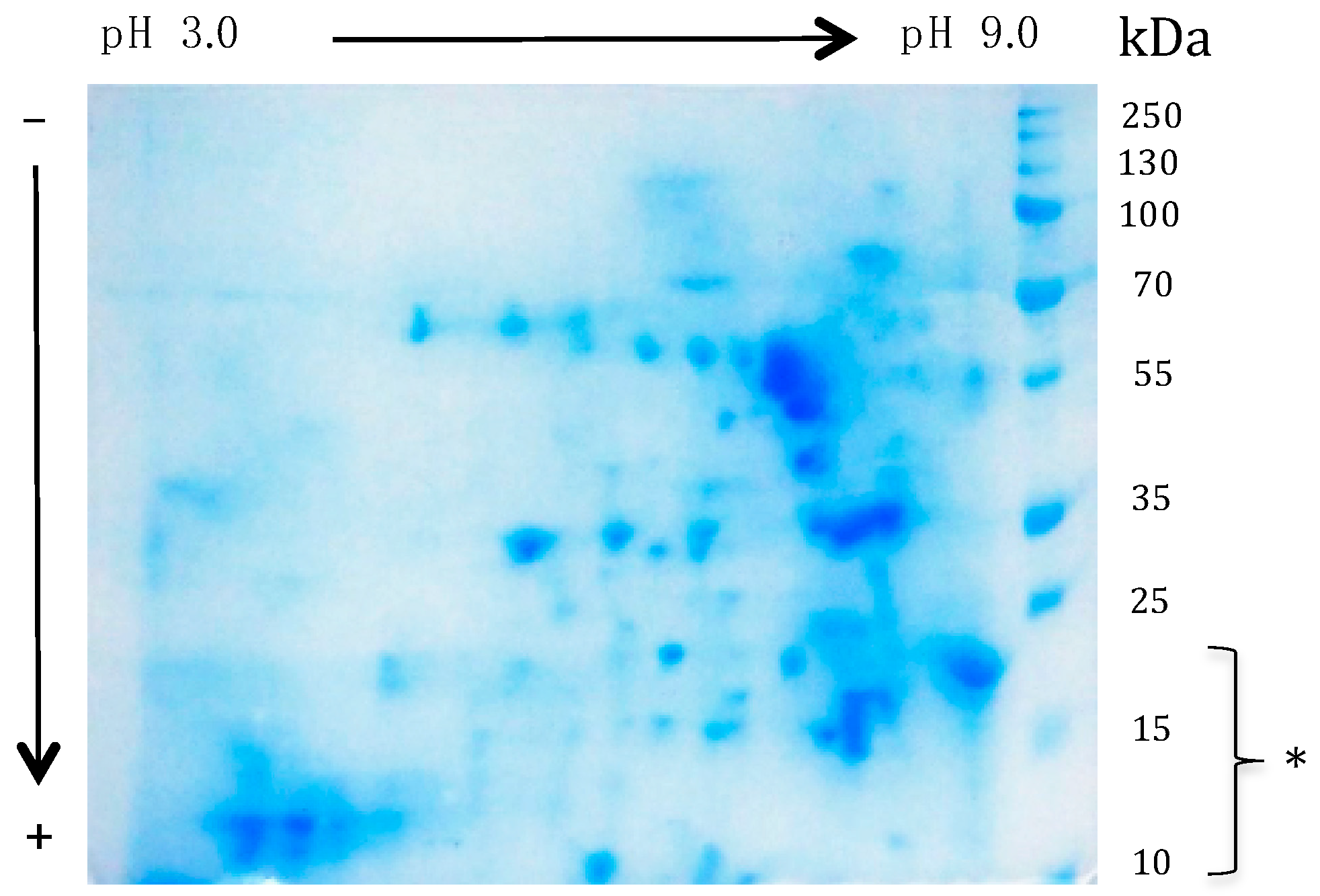
3.3. Characterization of the Potential Allergenic Protein Repertoire of the Mealworm Protein Extract
3.4. The Cross-Reacting Allergens of the Mealworm Protein Extract Are Functional in Degranulation Tests In Vitro
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seum: | Gender: | Shrimp Specific IgE (kU·mL−1) | Der p 10 Specific IgE (kU·mL−1) |
|---|---|---|---|
| 1a | Female | 81.1 | |
| 2a | Female | 45.3 | |
| 3a | Female | 49.8 | |
| 4a | Male | 28.2 | >100 |
| 5a | Female | 21.1 | |
| 6a | Male | >100 | |
| 7a | Female | 66.8 | |
| 8a | Male | >100 | |
| 9a | Male | 33.9 | |
| 10a | Female | 24.4 | |
| 11a | Male | 61.8 | |
| 12a | Female | 54.8 | |
| 13a | Male | 11.2 | 40.8 |
| 14a | Female | >100 | |
| 15a | Female | 18.8 | |
| 16a | Female | 66.8 | |
| 17a | Male | 11.8 | |
| 18a | Male | 29.4 | |
| 19a | Female | >100 | |
| 20a | Male | 52.1 | >100 |
| 21a | Female | 10.2 | |
| 1b | Male | >100 | |
| 2b | Female | >100 | |
| 3b | Female | >100 | |
| 4b | Female | 17.0 | >100 |
| 5b | Female | >100 | |
| 6b | Female | >100 | |
| 7b | Female | >100 | |
| 8b | Male | >100 | |
| 9b | Male | 1.4 | >100 |
| 10b | Female | 27.3 | >100 |
| 11b | Male | 2.4 | 84.6 |
| 12b | Female | 0.1 | 13.1 |
| 13b | Male | >100 |
References
- DeFoliart, G.; Nakagaki, B.; Sunde, M. Protein quality of the house cricket Acheta domestica when fed to broiler chicks. Poult. Sci. 1987, 66, 1367–1371. [Google Scholar]
- Oonincx, D.G.; de Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans—A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef] [PubMed]
- Rumpold, B.A.; Schluter, O.K. Potential and challenges of insects as an innovative source for food and feed production. Innov. Food Sci. Emerg. Technol. 2013, 17, 1–11. [Google Scholar] [CrossRef]
- Rumpold, B.A.; Schluter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, Z.; Liu, H. Feasibility of feeding yellow mealworm (Tenebrio molitor L.) in bioregenerative life support systems as a source of animal protein for humans. Acta Astron. 2013, 92, 103–109. [Google Scholar] [CrossRef]
- Oonincx, D.G.; van Broekhoven, S.; van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- Van Huis, A. Edible insects are the future? Proc. Nutr. Soc. 2016, 75, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Chen, X.M.; Zhao, M.; He, Z.; Sun, L.; Wang, C.Y.; Ding, W.F. Edible insects in China: Utilization and prospects. Insect Sci. 2017. [Google Scholar] [CrossRef]
- Van Broekhoven, S.; Oonincx, D.G.; van Huis, A.; van Loon, J.J. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef]
- Bußler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insects: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Helivon 2016, 2, e00218. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.P.; van den Broek, L.A.; Fogliano, V.; Lakemond, C.M. Nitrogen-to-protein conversion factors for thee edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible insects in a food safety and nutritional perspective: A critical review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; FAO Forestry Paper 171; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 978-92-5-107595-1. [Google Scholar]
- Charlton, A.J.; Dickinson, M.; Wakefield, M.E.; Fitches, E.; Kenis, M.; Han, R.; Zhu, F.; Kone, N.; Grant, M.; Devic, E.; et al. Exploring the chemical safety of fly larvae as a source of protein for animal feed. J. Insect Food Feed 2015, 1, 7–16. [Google Scholar] [CrossRef]
- Schlüter, O.; Rumpold, B.; Holzhauser, T.; Roth, A.; Vogel, R.F.; Quasigroch, W.; Vogel, S.; Heinz, V.; Jäger, H.; Bandick, N.; et al. Safety aspects of the production of foods and food ingredients from insects. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Testa, M.; Stillo, L.; Maffei, G.; Andriolo, V.; Gardois, P.; Zotti, C.M. Ugly but tasty: A systematic review of possible human and animal health risks related to entomophagy. Crit. Rev. Food Nutr. 2017, 57, 3747–3759. [Google Scholar] [CrossRef]
- Bernton, H.S.; Brown, H. Insects as potential sources of ingestant allergens. Ann. Allergy 1967, 25, 381–387. [Google Scholar]
- Phillips, J.; Burkholder, W. Allergies related to food insect production and consumption. Food Insect Newsl. 1995, 8, 1–2. [Google Scholar]
- Van der Brempt, X.; Moneret-Vautrin, D.A. The allergic risk of Tenebrio molitor for human consumption (article in French). Rev. Fr. Allergol. 2014, 54, 34–36. [Google Scholar] [CrossRef]
- Barre, A.; Caze-Subra, S.; Gironde, C.; Bienvenu, F.; Bienvenu, J.; Rougé, P. Entomophagy and the risk of allergy (article in French). Rev. Fr. Allergol. 2014, 54, 315–321. [Google Scholar] [CrossRef]
- Van der Brempt, X.; Beaudoin, E.; Lavaud, F. Is eating insects risky for allergic patients? Rev. Fr. Allergol. 2016, 56, 186–188. (In French) [Google Scholar] [CrossRef]
- Ribeiro, J.C.; Cunha, L.M.; Sousa-Pinto, B.; Fonseca, J. Allergic risks of consuming edible insects: A systematic review. Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Teranishi, H.; Kawai, K.; Murakami, G.; Miyao, M.; Kasuya, M. Occupational allergy to adult chironomid midges among environmental researchers. Jpn. Int. Arch. Allergy Immunol. 1995, 106, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Freye, H.B.; Esch, R.E.; Litwin, C.M.; Sorkin, L. Anaphylaxis to the ingestion and inhalation of Tenebrio molitor (mealworm) and Zophobas morio (superworm). Allergy Asthma Proc. 1996, 17, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Aldunate, M.T.; Echechipía, S.; Gómez, B.; García, B.E.; Olaguibel, J.M.; Rodríguez, A.; Moneo, I.; Tabat, A.I. Chironomids and other causes of fish food allergy. Allergol. Immunol. Clin. 1999, 14, 140–145. [Google Scholar]
- Ji, K.; Chen, J.; Li, M.; Liu, Z.; Wang, C.; Zhan, Z.; Wu, X.; Xia, Q. Anaphylactic shock and lethal anaphylaxis caused by food consumption in China. Trends Food Sci. Technol. 2009, 20, 227–231. [Google Scholar] [CrossRef]
- Choi, G.S.; Shin, Y.; Kim, J.E.; Ye, Y.M.; Park, H.S. Five cases of food allergy to vegetable worm (Cordyceps sinensis) showing cross-reactivity with silkworm pupæ. Allergy 2010, 65, 1196–1197. [Google Scholar] [CrossRef]
- Okezie, O.A.; Kgomotso, K.K.; Letswiti, M.M. Mopane worm allergy in a 36-year-old woman: A case report. J. Med. Case Rep. 2010, 4, 42. [Google Scholar] [CrossRef]
- Yew, K.L.; Kok, V.S.L. Exotic food anaphylaxis and the broken heart: Sago worm and Takotsubo cardiomyopathy. Med. J. Malays. 2012, 5, 540–541. [Google Scholar]
- Pener, M.P. Allergy to crickets: A review. J. Orthoptera Res. 2016, 25, 91–95. [Google Scholar] [CrossRef]
- Santos, A.B.R.; Chapman, M.D.; Aalberse, R.C.; Vailes, L.D.; Ferriani, V.P.L.; Oliver, C.; Rizzo, M.C.; Naspitz, C.K.; Arruda, L.K. Cockroach allergens and asthma in Brazil: Identification of tropomyosin as a major allergen with potential cross-reactivity with mite and shrimp allergens. J. Allergy Clin. Immunol. 1999, 104, 329–337. [Google Scholar] [CrossRef]
- Reese, G.; Ayuso, R.; Lehrer, S.B. Tropomyosin: An invertebrate pan-allergen. Int. Arch. Allergy Immunol. 1999, 119, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Galindo, P.A.; Lombardero, M.; Borja, J.E.G.; Feo, F.; Barber, D.; García, R. A new arthropod panallergen? Allergy 2001, 56, 195–197. [Google Scholar] [CrossRef] [PubMed]
- Shafique, R.H.; Inam, M.; Ismail, M.; Chaudhary, F.R. Group 10 allergens (tropomyosins) from house-dust mites may cause covariation of sensitization to allergens from other invertebrates. Allergy Rhinol. 2010, 3, e74–e90. [Google Scholar] [CrossRef] [PubMed]
- Binder, M.; Mahler, V.; Hayek, B.; Sperr, W.R.; Schöller, M.; Prozell, S.; Wiedermann, G.; Valent, P.; Valenta, R.; Duchêne, M. Molecular and immunological characterization of arginine kinase from the Indianmeal moth, Plodia interpunctella, a novel cross-reactive invertebrate pan-allergen. J. Immunol. 2001, 167, 5470–5477. [Google Scholar] [CrossRef]
- Verhoeckx, K.C.M.; van Broekhoven, S.; den Hartog-Jager, C.F.; Gaspari, M.; de Jong, G.A.H.; Wichers, H.J.; van Hoffen, E.; Houben, G.F.; Knulst, A.C. House dust mite (Der p 10) and crustacean allergic patients may react to food containing yellow mealworm proteins. Food Chem. Toxicol. 2014, 65, 364–373. [Google Scholar] [CrossRef]
- Broekman, H.; Knulst, A.; den Hartog Jager, S.; Monteleone, F.; Gaspari, M.; de Jong, G.; Houben, G.; Verhoeckx, K. Effect of thermal processing on mealworm allergenicity. Mol. Nutr. Food Res. 2015, 59, 1855–1864. [Google Scholar] [CrossRef]
- Van Broekhoven, S.; Bastiaan-Net, S.; de Jong, N.W.; Wichers, H.J. Influence of processing and in vitro digestion on the allergic cross-reactivity of three mealworm species. Food Chem. 2016, 196, 1075–1083. [Google Scholar] [CrossRef]
- Debaugnies, F.; Francis, F.; Delporte, C.; Doyen, V.; Lendent, C.; Mairesse, M.; Van Antwerpen, P.; Corraza, F. Identification de l’α-amylase comme allergène du ver de farine chez des patients professionnellement exposés. Rev. Fr. Allergol. 2016, 56, 281. (In French) [Google Scholar] [CrossRef]
- Srinroch, C.; Srisomsap, C.; Chokchaichamnankit, D.; Punyarit, P.; Phiriyangkul, P. Identification of novel allergen in edible insect, Gryllus bimaculatus and its cross-reactivity with Macrobrachium spp. allergens. Food Chem. 2015, 184, 160–166. [Google Scholar] [CrossRef]
- Just, N.; Lièvre, K.; Lallemand, K.; Leduc, V. Implication de l’hexamérine dans un cas d’allergie aux larves de mouches. Rev. Fr. Allergol. 2012, 52, 258. (In French) [Google Scholar] [CrossRef]
- De Gier, S.; Verhoeckx, K. Insect (food) allergy and allergens. Mol. Immunol. 2018, 100, 82–106. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, T.W.; Williams, P.B.; Dreskin, S.C.; Jouvin, M.H.; Kinet, J.P.; Tasset, D. High-affinity oligonucleotide ligands to human IgE inhibit binding to Fc epsilon receptor I. J. Immunol. 1996, 157, 221–230. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL-X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tool. Nucleic Acids Res. 1999, 15, 4876–4882. [Google Scholar]
- Risler, J.-L.; Delorme, M.O.; Delacroix, H.; Henaut, A. Amino acid substitutions in structurally related proteins. A pattern recognition approach. Determination of a new an efficient scoring matrix. J. Mol. Biol. 1998, 204, 1019–1029. [Google Scholar] [CrossRef]
- Breiter, D.R.; Kanost, M.R.; Benning, M.M.; Wesenberg, G.; Law, J.H.; Wells, M.A.; Rayment, I.; Holden, H.M. Molecular structure of an aloplipoprotein determined at 2.5-Å resolution. Biochemistry 1991, 30, 603–608. [Google Scholar] [CrossRef]
- Fan, D.; Zheng, Y.; Yang, D.; Wang, J. NMR solution structure and dynamics of an exchanh-geable apolipoprotein, Locusta migratoria apolipophorin III. J. Biol. Chem. 2003, 278, 21212–21220. [Google Scholar] [CrossRef]
- Wang, J.; Sykes, B.D.; Ryan, R.O. Structural basis for the conformational adaptability of apolipophorin III, a helix-bundle exchangeable apolipoprotein. Proc. Natl. Acad. Sci. USA 2002, 99, 1188–1193. [Google Scholar] [CrossRef] [Green Version]
- Rothemund, S.; Liou, Y.C.; Davies, P.L.; Krause, E.; Sönnichsen, F.D. A new class of hexahelical insect proteins revealed as putative carriers of small hydrophobic ligands. Structure 1999, 7, 1325–1332. [Google Scholar] [CrossRef] [Green Version]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the precision of comparative models with YASARA NOVA—A self-parameterizing force field. Proteins 2002, 47, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, S.; Lagarde, A.; Iovinella, P.; Legrand, P.; Tegoni, M.; Pelosi, P.; Cambillau, C. Crystal structure of Apis mellifera OBP14, a C-minus odorant-binding protein, and its complexes with odorant molecules. Insect Biochem. Mol. Biol. 2012, 42, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, A.; Spinelli, S.; Tegoni, M.; He, X.; Field, L.; Zhou, J.J.; Cambillau, C. The crystal structure of odorant binding protein 7 from Anopheles gambiae exhibits an outstanding adaptability of its binding site. J. Mol. Biol. 2011, 414, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Di Luccio, E.; Ishjida, Y.; Leal, W.S.; Wilson, D.K. Crystallographic observation of pH-induced conformational changes in the Amyelois transitella pheromone-binding protein AtraPBP1. PLoS ONE 2013, 8, e53840. [Google Scholar] [CrossRef]
- Damberger, F.F.; Ishida, Y.; Leal, W.S.; Wüthrich, K. Structural basis of ligand binding and release in insect pheromone-binding protein: NMR structure of Antheraea polyphemus PBP1 at pH 450. J. Mol. Biol. 2007, 373, 811–819. [Google Scholar] [CrossRef]
- Zhou, J.J.; Robertson, G.; He, X.; Dufour, S.; Hooper, A.M.; Pickett, J.A.; Keep, N.H.; Field, L.M. Characterisation of Bombyx mori odorant-binding proteins reveals that a general odorant-binding protein discriminates between sex pheromone components. J. Mol. Biol. 2009, 389, 529–545. [Google Scholar] [CrossRef]
- Campanacci, V.; Lartigue, A.; Hällberg, B.M.; Jones, T.A.; Giudici-Orticoni, M.T.; Tegoni, M.; Cambillau, C. Moth chemosensory protein exhibits drastic conformational changes and cooperativity on ligand binding. Proc. Natl. Acad. Sci. USA 2003, 100, 5069–5074. [Google Scholar] [CrossRef] [Green Version]
- Jansen, S.; Chmelik, J.; Zidek, L.; Padrta, P.; Novák, P.; Zdráhal, Z.; Picimbon, J.F.; Löfstedt, C.; Sklenár, V. Structure of Bombyx mori chemosensory protein 1 in solution. Arch. Insect Biochem. Physiol. 2007, 66, 135–145. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemistry of protein structures. J. Appl. Cryst. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Melo, F.; Feytmans, E. Assessing protein structures with a non-local atomic interaction energy. J. Mol. Biol. 1998, 277, 1141–1152. [Google Scholar] [CrossRef]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Glaser, F.; Pupko, T.; Bell, R.E.; Bechor, D.; Martz, E.; Ben-Tal, N. ConSurf: Identification of functional regions in proteins by surface-mapping of phylogenetic informations. Bioinformatics 2003, 19, 163–164. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Radauer, C.; Bublin, M.; Wagner, S.; Mari, A.; Breiteneder, H. Allergens are distributed into few protein families and possess a restricted number of biochemical functions. J. Allergy Clin. Immunol. 2008, 121, 847–852. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Borges, M.; Capriles-Hulett, A.; Fernández-Caldas, E.; Suárez-Chacón, R.; Caballero, F.; Castillo, S.; Sotillo, E. Mite-contaminated foods as a cause of anaphylaxis. J. Allergy Clin. Immunol. 1997, 99, 738–743. [Google Scholar] [CrossRef]
- Sánchez-Borges, M.; Suárez-Chacón, R.; Capriles-Hulett, A.; Caballero-Fonseca, F.; Iraola, V.; Fernández-Caldas, E. Anaphylaxis from ingestion of mites: Pancake anaphylaxis. J. Allergy Clin. Immunol. 2013, 131, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Y.; Liu, G.M.; Cai, Q.F.; Weng, W.Y.; Maleki, S.J.; Su, W.J.; Cao, M.J. Stability of major allergen tropomyosin and other food proteins of mud crab (Scylla serrata) by in vitro gastrointestinal digestion. Food. Chem. Toxicol. 2010, 48, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Sahabudin, S.; Misnan, R.; Yadzir, Z.H.; Mohamad, J.; Abdullah, N.; Bakhtiar, F.; Murad, S. Identification of major and minor allergens of black tiger prawn (Penaeus monodon) and king prawn (Penaeus latisulcatus). Malays. J. Med. Sci. 2011, 18, 2732. [Google Scholar]
- Yadzir, Z.H.M.; Misnan, R.; Abdullah, N.; Bakhtiar, F.; Arip, M.; Murad, S. Identification of the major allergen of Macrobrachium rosenbergii (giant freshwater prawn). Asian Pac. J. Trop. Biomed. 2012, 2, 50–54. [Google Scholar] [CrossRef]
- Rosmilah, M.; Shahnaz, M.; Zailatul, H.M.Y.; Noormalin, A.; Normilah, I. Identification of tropomyosin and arginine kinase as major allergens of Portunus pelagicus (blue swimming crab). Trop. Biomed. 2012, 29, 467–478. [Google Scholar] [PubMed]
- Misnan, R.; Murad, S.; Yadzir, Z.H.; Abdullah, N. Identification of the major allergens of Charybdis feriatus (red crab) and its cross-reactivity with Portunus pelagicus (blue crab). Asian Pac. J. Allergy Immunol. 2012, 30, 285–293. [Google Scholar] [PubMed]
- Meseguer Arce, J.; Sánchez-Guerrero Villajos, I.M.; Iraola, V.; Carnés, J.; Fernández Caldas, E. Occupational allergy to aquarium fish food: Red midge larva, freshwater shrimp, and earthworm. A clinical and immunological study. J. Investig. Allergol. Clin. Immunol. 2013, 23, 462–470. [Google Scholar]
- Yadzir, Z.H.; Misnan, R.; Bakhtiar, F.; Abdullah, N.; Murad, S. Tropomyosin, the major tropical oyster Crassostrea belcheri allergen and effect of cooking on its allergenicity. Allergy Asthma Clin. Immunol. 2015, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Broekman, H.; Verhoeckx, K.C.; den Hartog Jager, C.F.; Kruizinga, A.G.; Pronk-Kleinjan, M.; Remington, B.C.; Bruijnzeel-Koomen, C.A.; Houben, G.F.; Knulst, A.C. Majority of shrimp-allergic patients are allergic to mealworm. Allergy Clin Immunol. 2016, 137, 1261–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondot, I.; Quennedey, B.; Delachambre, J. Structure, organization and expression of two clustered cuticle protein genes during the metamorphosis of an insect, Tenebrio molitor. Eur. J. Biochem. 1998, 254, 304–312. [Google Scholar] [CrossRef]
- Rougé, P.; Barre, A. Allergy to edible insects: A computational identification of the IgE-binding cross-reacting allergen repertoire of edible insects. In Future Foods; Mikkola, H., Ed.; INTECH: Croatia, Balkans, 2017; Volume 4, pp. 71–88. [Google Scholar]
- Nadeau, L.; Nadeau, I.; Franklin, F.; Dunkel, F. The potential for entomophagy to address undernutrition. Ecol. Food Nutr. 2015, 54, 200–208. [Google Scholar] [CrossRef]
- Barennes, H.; Phimmasane, M.; Rajaonarivo, C. Insect consumption to address undernutrition, a national survey of the prevalence of insect consumption among adults and vendors in Laos. PLoS ONE 2015, 10, e0136458. [Google Scholar] [CrossRef]
- Payne, C.L.; Scarborough, P.; Rayner, M.; Nonaka, K. Are edible insects more or less ‘healthy’ than commonly consumed meats? A comparison using two nutrient profiling models developed to combat over- and undernutrition. Eur. J. Clin. Nutr. 2016, 70, 285–291. [Google Scholar] [CrossRef]
- Laar, A.; Kotoh, A.; Parker, M.; Milani, P.; Tawiah, C.; Soor, S.; Anankware, J.P.; Kalra, N.; Manu, G.; Tandoh, A.; et al. An exploration of edible palm weevil larvae (akokono) as a source of nutrition and livelihood: Perspectives from Ghanaian stakeholders. Food Nutr. Bull. 2017, 38, 455–467. [Google Scholar] [CrossRef]
- Dai, C.; Ma, H.; Luo, L.; Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur. Food Res. Technol. 2013, 236, 681–689. [Google Scholar] [CrossRef]
- Cito, A.; Dreassi, E.; Frosinini, R.; Zanfini, A.; Pianigiani, C.; Botta, M.; Francardi, V. The potential beneficial effects of Tenebrio molitor (Coleoptera: Tenebrionidae) and Galleria mellonella (Lepidoptera: Pyralidae) on human health. Redia 2017, 100, 125–133. [Google Scholar]
- Lee, W.; Kim, M.A.; Park, I.; Hwang, J.S.; Na, M.; Bae, J.S. Novel direct factor Xa inhibitory compounds from Tenebrio molitor with anti-platelet aggregation activity. Food Chem. Toxicol. 2017, 109, 19–27. [Google Scholar] [CrossRef]
- Dutta, P.; Dey, T.; Manna, P.; Kalita, J. Antioxidant potential of Vespa affinis L., a traditional edible insect species of North East India. PLoS ONE 2016, 11, e0156107. [Google Scholar] [CrossRef]
- Dutta, P.; Dey, T.; Dihingia, A.; Manna, P.; Kalita, J. Antioxidant and glucose metabolizing potential of edible insect, Brachytrupes orientalis via modulating Nrf2/AMPK/GLUT4 signaling pathway. Biomed. Pharmacother. 2017, 95, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Wendin, K.; Olsson, V.; Langton, M. Mealworms as food ingredient—Sensory investigation of a model system. Foods 2019, 8, 319. [Google Scholar] [CrossRef] [PubMed]
- Pali-Schöll, I.; Meinlschmidt, P.; Larenas-Linnemann, D.; Purschke, B.; Hofstetter, G.; Rodríguez-Monroy, F.A.; Einhorn, L.; Mothes-Luksch, N.; Jensen-Jarolim, E.; Jäger, H. Edible insects: Cross-recognition of IgE from crustacean- and house dust mite allergic patients, and reduction of allergenicity by food processing. World Allergy Organ. J. 2019, 12, 100006. [Google Scholar] [CrossRef] [Green Version]









| Score: | Protein: | Score: | Protein: |
|---|---|---|---|
| 2984.019 | α-Myosin | 222.675 | 28 kDa dessication stress protein |
| 1214.674 | Aldehyde oxidase | 219.963 | Masquerade-like serine protease |
| 1066.813 | Ca-transporting ATPase | 213.041 | 13 kDa hemolymph protein d |
| 1025.416 | Actin 87E | 211.853 | Larval cuticle protein A3A |
| 940.811 | 86 kDa Early stage encapsulation | 211.417 | 12 kDa hemolymph protein c |
| inducing protein | 210.607 | Myosin light chain alkali | |
| 895.841 | Tropomyosin 2 | 209.594 | Fructose-biphosphate aldolase |
| 806.565 | Actin 5C | 209.332 | Serine proteinase |
| 765.02 | Actinin α | 207.818 | Glutathione S-transferase |
| 740.12 | Tubulin β | 201.438 | 13 kDa hemolymph protein b |
| 664.062 | Prophenoloxidase | 198.188 | Vitellogenin receptor |
| 609.664 | Tropomyosin 1 | 190.799 | Troponin C |
| ATP synthase subunit β | 189.968 | Calreticulin | |
| 504.441 | α-Amylase | 187.099 | 4.5 LIM domains protein 5 |
| 493.123 | Tubulin α | 184.454 | Cytochrome c2 |
| 474.845 | Odorant binding protein OBP 14 | 179.106 | Nucleoside triphosphatase A |
| 474.053 | Apolipophorin-III | 177.997 | Muscle-specific protein 20 |
| 446.015 | Myosin heavy chain, non-muscle | 176.202 | Muscle LIM protein Mlp84B |
| 445.547 | Arginine kinase | 171.809 | Enolase |
| 439.842 | HSP 70 | 171.39 | Obstractor C2 |
| 423.306 | Filamin-A | 168.27 | Protein disulfide-isomerase |
| 395.328 | 12 kDa hemolymph protein a | 164.625 | 13 kDa hemolymph protein c |
| 386.89 | Twitchin-like protein | 161.351 | Phosphoglycerate kinase |
| 370.18 | Cockroach allergen-like protein | 159.53 | Chemosensory protein CSP12 |
| 363.618 | Serpin 1 | 157.539 | α-Spectrin |
| 359.558 | ATP synthase subunit α | 157.443 | 14.3.3. ζ |
| 340.168 | Hexamerin 2 | 155.443 | Glycogenin-1-like protein |
| 338.385 | Glyceraldehyde-3-phosphate | 151.351 | Phosphoglycerate kinase |
| dehydrogenase | 147.874 | Ferritin | |
| 337.266 | 56 kDa Early stage encapsulation | 142.987 | Calumenin |
| inducing protein | 141.375 | Cathepsin L11 | |
| 326.698 | Voltage-dependant anion-selective | 140.034 | Ribosomal protein S3 |
| channel protein | 132.912 | Nucleobindin-2 like protein | |
| 315.75 | Paramyosin long form | 130.118 | Trypsin |
| 313.141 | Serpin 93 kDa | 129.432 | G protein-coupled receptor kinase 1 |
| 296.596 | Larval cuticle protein A1A | 124.835 | THP isoform 84aa-XY |
| 286.543 | Serpin 40 | 122.826 | Chitin deacetylase 1 |
| 280.057 | Cathepsin F10 | 122.326 | Myosin regulatory light chain-2 |
| 279.884 | α-1,4-glucan phosphorylase | 122.024 | Malate dehydrogenase |
| 278.386 | Larval cuticle protein A2B | 121.525 | Cuticular protein |
| 278.215 | Troponin T | 118.956 | Chitinase |
| 276.285 | Melanization-related protein | 116.322 | Glucose-6-phosphate isomerase |
| 273.213 | Paramyosin short form | 99.944 | Carbonyl reductase 1 |
| 269.133 | Peptidyl-prolyl cis-trans isomerase | 98.68 | Protein lethal/essential for life |
| 260.731 | Apolipophorin-like protein | 96.268 | Transketolase-like protein 2 |
| 259.533 | Myosin heavy chain, muscle | 89.762 | Multiple coagulation factor |
| 259.092 | HSP 60 | deficiency protein 2 | |
| 254.399 | V-type proton ATPase subunit β | 87.931 | 60S ribosomal protein L31 |
| 251.439 | Pyruvate kinase | 87.574 | ATP carrier protein |
| 249.266 | Melanin-inhibiting protein | 87.104 | Lysosomal aspartic protease |
| 247.353 | Serpin 48 | 78.888 | Aspartate aminotransferase |
| 246.2 | Superoxide dismutase(Cu-Zn) | 76.674 | Elongation factor 1-α |
| 237.263 | 12 kDa hemolymph protein b | 69.44 | Adenosylhomocysteinase |
| 229.745 | RNA-binding protein squid-like | 59.528 | Muscular protein 20 |
| 227.752 | Sarcalumenin | 34.596 | Reticulon-like protein |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barre, A.; Pichereaux, C.; Velazquez, E.; Maudouit, A.; Simplicien, M.; Garnier, L.; Bienvenu, F.; Bienvenu, J.; Burlet-Schiltz, O.; Auriol, C.; et al. Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor). Foods 2019, 8, 515. https://doi.org/10.3390/foods8100515
Barre A, Pichereaux C, Velazquez E, Maudouit A, Simplicien M, Garnier L, Bienvenu F, Bienvenu J, Burlet-Schiltz O, Auriol C, et al. Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor). Foods. 2019; 8(10):515. https://doi.org/10.3390/foods8100515
Chicago/Turabian StyleBarre, Annick, Carole Pichereaux, Esmeralda Velazquez, Agathe Maudouit, Mathias Simplicien, Lorna Garnier, Françoise Bienvenu, Jacques Bienvenu, Odile Burlet-Schiltz, Cédric Auriol, and et al. 2019. "Insights into the Allergenic Potential of the Edible Yellow Mealworm (Tenebrio molitor)" Foods 8, no. 10: 515. https://doi.org/10.3390/foods8100515