The Influence of pH, NaCl, and the Deacidifying Yeasts Debaryomyces hansenii and Kluyveromyces marxianus on the Production of Pigments by the Cheese-Ripening Bacteria Arthrobacter arilaitensis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultures
2.1.1. Yeast and Bacterial Strains
2.1.2. Cultures
2.2. Deacidified Cheese-Based Media and Factors Controlled
2.2.1. pH Adjustment
Chemical Deacidification
Biological Deacidification
2.2.2. Inoculation of Arthrobacter arilaitensis and Cultivation Conditions
2.3. Color Measurements
2.4. Statistical Analysis
3. Results
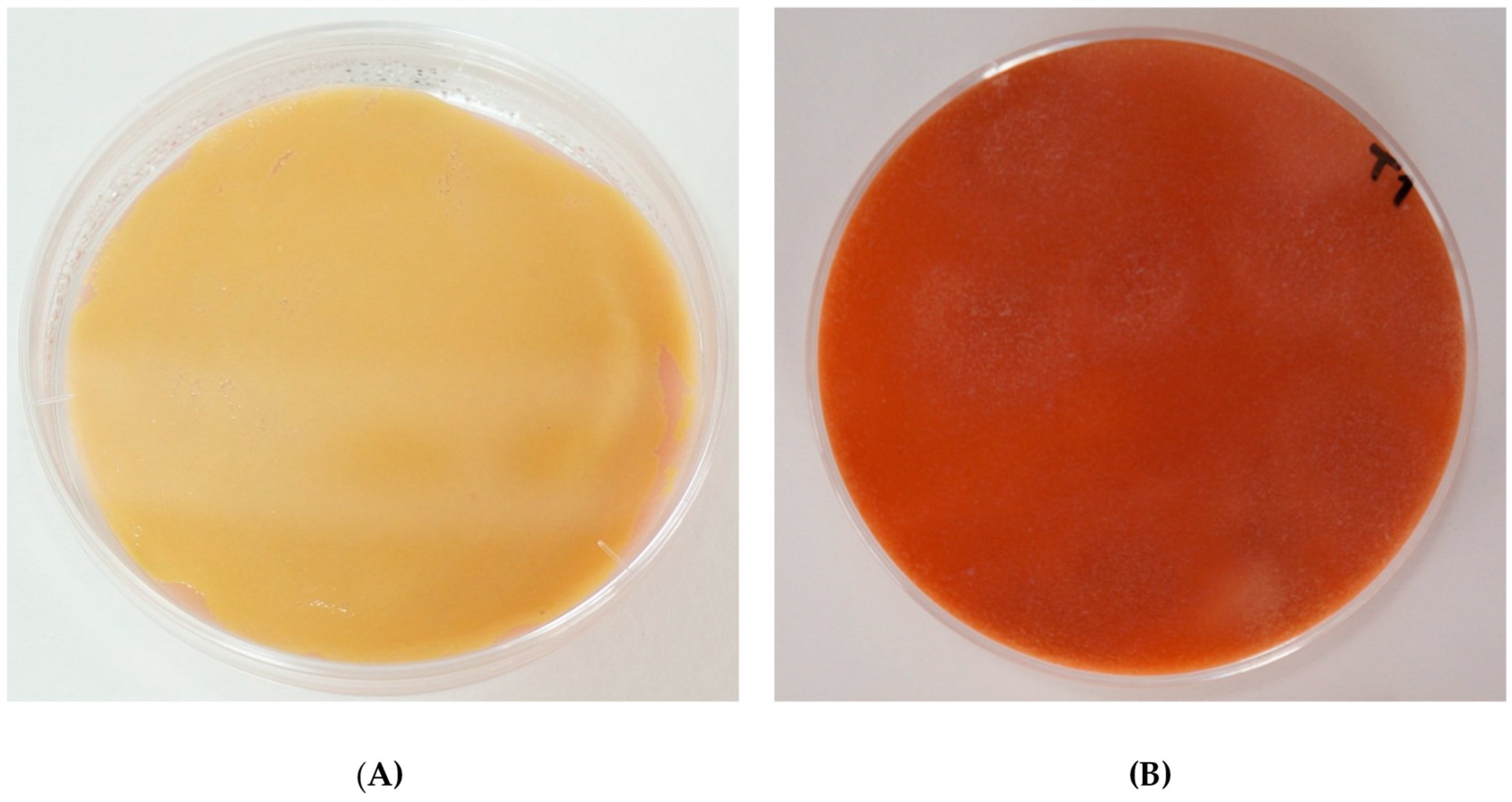
3.1. Diversity of Coloration by Arthrobacter arilaitensis Po102 on Different Deacidified Cheese-Based Media
3.2. The Effects of pH, NaCl, and Deacidification on the Coloration of Arthrobacter arilaitensis Po102
3.2.1. Changes in Color Development of Arthrobacter arilaitensis Po102
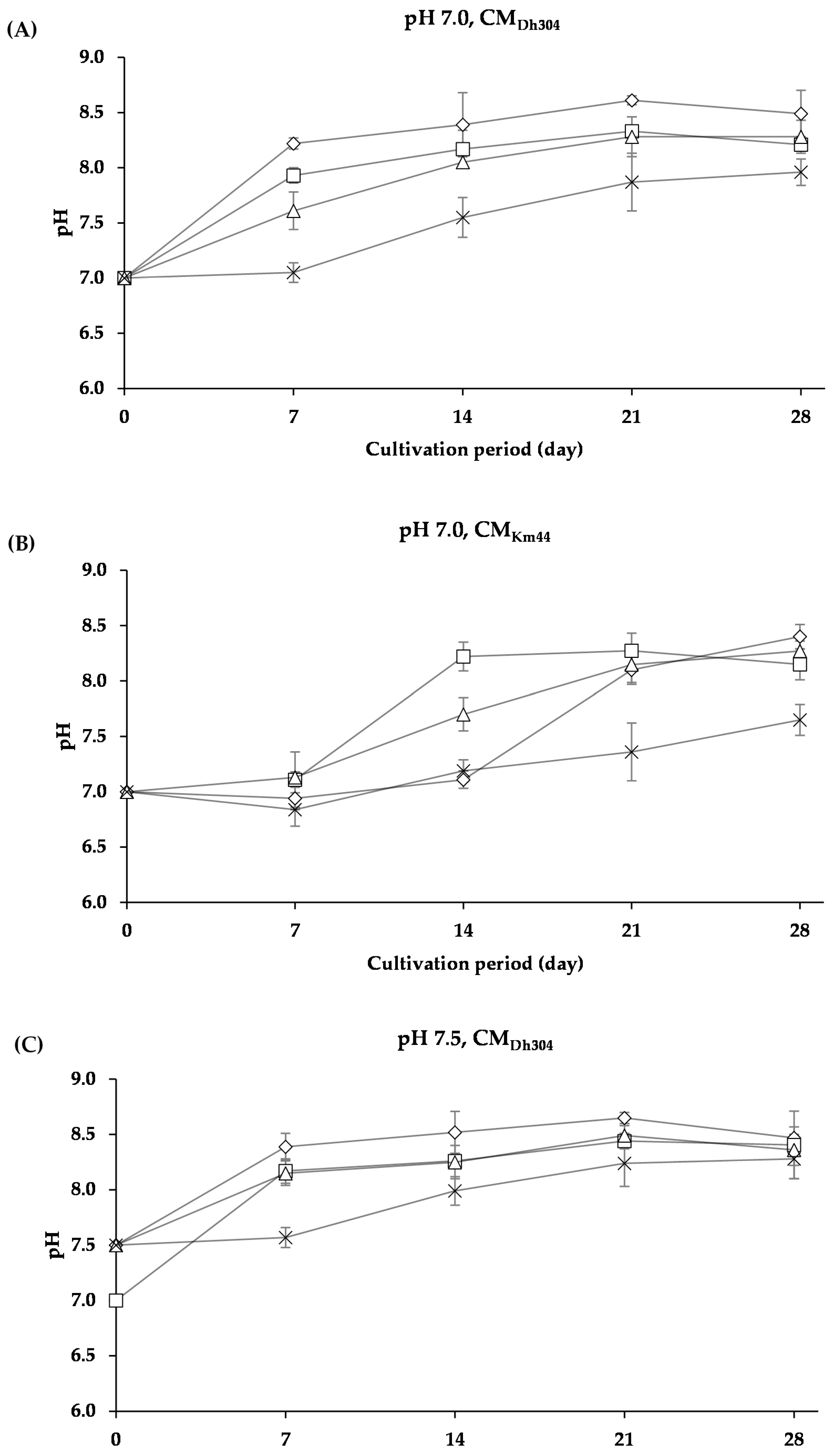
3.2.2. Changes in pH of Arthrobacter arilaitensis Po102 Cultures
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dufossé, L.; Mabon, P.; Binet, A. Assessment of the coloring strength of Brevibacterium linens strains: Spectrocolorimetry versus total carotenoid extraction/quantification. J. Dairy Sci. 2001, 84, 354–360. [Google Scholar] [CrossRef]
- Irlinger, F.; Layec, S.; Hélinck, S.; Dugat-Bony, E. Cheese rind microbial communities: Diversity, composition and origin. FEMS Microbiol. Lett. 2015, 362, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Eliskases-Lechner, F.; Ginzinger, W. The bacterial flora of surface-ripened chesses with special regard to corynforms. Le Lait 1995, 75, 571–584. [Google Scholar] [CrossRef]
- Bockelmann, W. Development of defined surface starter cultures for the ripening of smear cheeses. Int. Dairy J. 2002, 12, 123–131. [Google Scholar] [CrossRef]
- Brennan, N.; Ward, A.C.; Berford, T.; Fox, P.; Goodfellow, M.; Cogan, T. Biodiversity of the bacteria flora on the surface of a smear cheese. Appl. Environ. Microbiol. 2002, 68, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Feurer, C.; Vallaeys, T.; Corrieu, G.; Irlinger, F. Does smearing inoculum reflect the bacterial composition of the smear at the end of the ripening of a French soft, red smear cheese? J. Dairy Sci. 2004, 87, 3189–3197. [Google Scholar] [CrossRef]
- Larpin-Laborde, S.; Imran, M.; Bonaïti, C.; Bora, N.; Gelsomino, R.; Goerges, S.; Irlinger, F.; Goodfellow, M.; Ward, A.C.; Vancanneyt, M.; et al. Surface microbial consortia from Livarot, a French smear-ripened cheese. Can. J. Microbiol. 2011, 57, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Irlinger, F.; Bimet, G.; Delettre, J.; Lefèvre, M.; Grimont, P.A.D. Arhtrobacter bergerei sp. nov. and Arthrobacter arilaitensis sp. nov., novel corynform species isolated from the surface of cheeses. Int. J. Syst. Evol. Microbiol. 2005, 55, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Sutthiwong, N.; Caro, Y.; Milhau, C.; Valla, A.; Fouillaud, M.; Dufossé, L. Arthrobacter arilaitensis strains isolated from ripened cheeses: Characterization of their pigmentation using spectrocolorimetry. Food Res. Int. 2014, 65, 184–192. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Dufossé, L. Production of carotenoids by Arthrobacter arilaitensis strains isolated from smear-ripened cheeses. FEMS Res. Lett. 2014, 360, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Guiffrida, D.; Sutthiwong, N.; Dugo, P.; Donato, P.; Cacciola, F.; Girad-Valenciennes, E.; Mao, Y.; Monnet, C.; Fouillaud, M.; Caro, Y.; et al. Characterisation of the C50 carotenoids produced by strains of the cheese-ripening bacterium Arthrobacter arilaitensis. Int. Dairy J. 2016, 55, 10–16. [Google Scholar] [CrossRef]
- Reps, A. Bacterial smear-ripened cheese. In Cheese: Chemistry, Physics and Microbiology. Volume 2. Major Cheese Group; Fox, P.F., Ed.; Chapman and Hall: London, UK; pp. 137–172.
- Kindstedt, P.S. The basics of cheesemaking. Microbiol. Spectr. 2013, 1, CM-0002. [Google Scholar] [CrossRef] [PubMed]
- Leclercq-Perlat, M.N.; Oumer, A.; Buono, F.; Bergere, J.L.; Spinnler, H.E.; Corrieu, G. Behavior of Brevibacterium linens and Debaryomyces hansenii as ripening flora in controlled production of soft smear cheese from reconstituted milk: Protein degradation. J. Dairy Sci. 2000, 83, 1674–1683. [Google Scholar] [CrossRef]
- Dugat-Bony, E.; Straub, C.; Teissandier, A.; Onésime, D.; Loux, V.; Monnet, C.; Irlinger, F.; Landaud, S.; Leclercq-Perlat, M.-N.; Bento, P.; et al. Overview of a surface-ripened cheese community functioning by meta-omics anylyses. PLoS ONE 2015, 10, e0124360. [Google Scholar] [CrossRef] [PubMed]
- Irlinger, F.; Mounier, J. Microbial interactions in cheese: Implications for cheese quality and safety. Curr. Opin. Biotechnol. 2009, 20, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclercq-Perlat, M.N.; Corrieu, G.; Spinnler, H.E. The color of Brevibacterium linens depends on the yeast used for cheese deacidification. J. Dairy Sci. 2004, 87, 1536–1544. [Google Scholar] [CrossRef]
- Masoud, W.; Jakobson, M. The combined effects of pH, NaCl and temperature on growth of cheese ripening cultures of Debaryomyces hansenii and coryneform bacteria. Int. Dairy J. 2005, 15, 69–77. [Google Scholar] [CrossRef]
- Nedomová, Š.; Kilián, L.; Pytel, R.; Kumbár, V. Effect of ripening time on colour and texture properties in cheese. Slovak J. Food. Sci. 2017, 11, 296–301. [Google Scholar] [CrossRef]
- Leclercq-Perlat, M.N.; Sicard, M.; Trelea, I.C.; Picque, D.; Corrieu, G. Temperature and relative humidity influence the ripening descriptors of Camembert-type cheeses throughout ripening. J. Diary Sci. 2015, 98, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masoud, W.; Jakobsen, M. Surface ripened cheeses: The effects of Debaryomyces hansenii, NaCl and pH on the intensity of pigmentation produced by Brevibacterium linens and Corynebacterium flavescens. Int. Dairy J. 2003, 13, 231–237. [Google Scholar] [CrossRef]
- Mounier, J.; Irlinger, F.; Leclercq-Perlat, M.-N.; Sarthou, A.-S.; Spinnler, H.-E.; Fitzgerald, G.F.; Cogan, T.M. Growth and colour development of some surface ripening bacteria with Debaryomyces hansenii on aseptic cheese curd. J. Dairy Res. 2006, 73, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Rea, M.C.; Goerges, S.; Gelsomino, R.; Brennan, N.M.; Mounier, J.; Vancanneyt, M.; Scherer, S.; Swings, J.; Cogan, T.M. Stability of the biodiversity of the surface consortia of Gubbeen, a red-smear cheese. J. Dairy Sci. 2007, 90, 2200–2210. [Google Scholar] [CrossRef] [PubMed]
- Martìnez-Cuesta, M.C.; Palencia, P.F.D.; Requena, T.; Pelácz, C. Enzymatic ability of Lactobacillus casei subsp. casei IFPL731 for flavor development in cheese. Int. Dairy J. 2001, 11, 577–585. [Google Scholar] [CrossRef]
- Swearingen, P.A.; O’Sullivan, D.J.; Warthsen, J.J. Isolation, characterization, and influence of native, nonstarter lactic acid bacteria on Chedda cheese quality. J. Dairy Sci. 2001, 84, 50–59. [Google Scholar] [CrossRef]
- Bockelmann, W.; Fuehr, C.; Martin, D.; Heller, K.J. Color development by Red-Smear surface bacteria. Kiel. Milchwirt. Forsch. 1997, 49, 285–292. [Google Scholar]
- Sutthiwong, N.; Fouillaud, M.; Valla, A.; Caro, Y.; Dufossé, L. Bacteria belonging to the extremely versatile genus Arthrobacter as novel source of natural pigments with extended hue range. Food Res. Int. 2014, 65, 156–162. [Google Scholar] [CrossRef]
- Cleary, J.L.; Kolachina, S.; Wolfe, B.E.; Sanchez, L.M. Coproporphyrin III produced by the bacterium Glutamicibacter arilaitensis binds zinc and is upregulated by fungi cheese rinds. Appl. Environ. Sci. 2018, 3, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Leclercq-Perlat, M.N.; Spinnler, H.E. The type of cheese curds determined the colouring capacity of Brevibacterium linens and Arthrobacter species. J. Dairy Res. 2010, 77, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Roostita, R.; Fleet, G.H. Growth of yeasts in milk and associated changes to milk composition. Int. J. Food Microbiol. 1996, 31, 205–219. [Google Scholar] [CrossRef]
- Daly, D.F.M.; MacSweeney, P.L.H.; Sheehan, J.J. Pink discolouration defect in commercial cheese: A review. Dairy Sci. Technol. 2012, 92, 439–453. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, D.J.; Daly, D.; O’Sullivan, O.; Burdikova, Z.; Vana, R.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; McSweeney, P.L.H.; et al. Thermus and the pink discoloration defect in cheese. Appl. Environ. Sci. 2016, 1. [Google Scholar] [CrossRef] [PubMed]
 = color of blank agar (without inoculating with A. arilaitensis Po102).
= color of blank agar (without inoculating with A. arilaitensis Po102).
= color of blank agar (without inoculating with A. arilaitensis Po102).
= color of blank agar (without inoculating with A. arilaitensis Po102).

 ), 2% (
), 2% (  ), 4% (
), 4% (  ), and 8% (
), and 8% (  ).
), 2% ( ), 4% ( ), and 8% ( ).
).
), 2% ( ), 4% ( ), and 8% ( ).
 ), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
 ), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ). ), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).
), 2% ( ), 4% ( ), and 8% ( ).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | NaCl (w/v) | CMNaOH | CMDh304 | CMKm44 | |||
|---|---|---|---|---|---|---|---|
| h° | C* | h° | C* | h° | C* | ||
| 5.8 | 0% | 87.04 ± 1.95 m | 24.53 ± 0.51 b | 80.53 ± 1.15 bc | 38.54 ± 1.48 a | 82.20 ± 2.20 b | 37.12 ± 1.12 a |
| 2% | 100.26 ± 1.02 a | 14.46 ± 0.13 cd | 104.19 ± 0.39 n | 7.71 ± 0.62 m | 98.37 ± 1.09 a | 13.60 ± 1.72 c | |
| 4% | 102.59 ± 0.83 o | 9.66 ± 1.04 n | 104.46 ± 0.88 p | 7.88 ± 1.08 o | 104.26 ± 1.28 q | 6.79 ± 0.54 p | |
| 8% | 105.53 ± 2.68 r | 7.42 ± 0.25 q | 105.74 ± 0.73 s | 6.71 ± 0.18 r | 105.25 ± 0.63 t | 6.56 ± 1.44 s | |
| 7.0 | 0% | 93.76 ± 0.76 u | 31.41 ± 0.69 d | 86.13 ± 0.72 de | 31.24 ± 2.23 t | 84.80 ± 1.67 d | 33.43 ± 1.17 u |
| 2% | 92.44 ± 1.43 i | 17.10 ± 0.25 v | 87.69 ± 2.13 v | 33.33 ± 2.31 w | 85.05 ± 2.31 w | 26.91 ± 0.14 e | |
| 4% | 95.35 ± 0.28 j | 14.81 ± 0.34 x | 87.08 ± 0.13 g | 35.17 ± 3.28 y | 89.13 ± 0.13 gh | 29.22 ± 2.58 l | |
| 8% | 95.12 ± 0.18 kl | 12.53 ± 0.94 g | 95.18 ± 0.38 l | 18.08 ± 2.42 h | 100.35 ± 1.25 x | 11.15 ± 1.55 g | |
| 7.5 | 0% | 80.86 ± 0.67 y | 25.37 ± 0.96 b | 83.04 ± 1.83 z | 26.18 ± 1.05 z | 83.47 ± 0.74 a* | 29.50 ± 1.53 a* |
| 2% | 94.46 ± 1.16 i | 24.17 ± 0.17 b* | 88.51 ± 0.37 ef | 30.19 ± 1.39 f | 88.85 ± 2.23 f | 29.86 ± 0.94 ef | |
| 4% | 96.33 ± 0.63 j | 23.70 ± 2.47 c* | 81.56 ± 0.56 b* | 31.89 ± 1.35 k | 89.52 ± 1.39 h | 29.84 ± 1.85 kl | |
| 8% | 99.55 ± 1.42 k | 22.16 ± 0.55 d* | 83.22 ± 2.05 c* | 17.46 ± 1.74 hi | 91.17 ± 0.33 d* | 19.87 ± 0.97 ij | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutthiwong, N.; Fouillaud, M.; Dufossé, L. The Influence of pH, NaCl, and the Deacidifying Yeasts Debaryomyces hansenii and Kluyveromyces marxianus on the Production of Pigments by the Cheese-Ripening Bacteria Arthrobacter arilaitensis. Foods 2018, 7, 190. https://doi.org/10.3390/foods7110190
Sutthiwong N, Fouillaud M, Dufossé L. The Influence of pH, NaCl, and the Deacidifying Yeasts Debaryomyces hansenii and Kluyveromyces marxianus on the Production of Pigments by the Cheese-Ripening Bacteria Arthrobacter arilaitensis. Foods. 2018; 7(11):190. https://doi.org/10.3390/foods7110190
Chicago/Turabian StyleSutthiwong, Nuthathai, Mireille Fouillaud, and Laurent Dufossé. 2018. "The Influence of pH, NaCl, and the Deacidifying Yeasts Debaryomyces hansenii and Kluyveromyces marxianus on the Production of Pigments by the Cheese-Ripening Bacteria Arthrobacter arilaitensis" Foods 7, no. 11: 190. https://doi.org/10.3390/foods7110190