
Impact of Molecular Weight Variations in Dendrobium officinale Polysaccharides on Antioxidant Activity and Anti-Obesity in Caenorhabditis elegans

Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Polysaccharides Extraction and Modification
2.3. Molecular Weight Determination of Polysaccharides
2.4. Analysis of Polysaccharide Components
2.5. Antioxidant Ability Detection In Vitro
2.5.1. Detection of the DPPH Radical (DPPH·) Scavenging Rate
2.5.2. Detection of the Hydroxyl Radical (·OH) Scavenging Rate
2.5.3. Total Antioxidant Capacity (T-AOC) Assay
2.6. C. elegans Assays
2.6.1. Culture and Synchronization of C. elegans
2.6.2. Lifespan Assay
2.6.3. Body Bend Assay
2.6.4. Pharyngeal Pumping Assay
2.6.5. Assessment of Antioxidant Stress Ability in C. elegans
2.6.6. Detection of Endogenous Antioxidant Enzymes
2.6.7. Analysis of Body Fat Accumulation in C. elegans Obesity Model
Oil Red O Staining Assay
TG Quantification
2.6.8. Gene Detection via q-PCR
2.7. Statistical Analysis
3. Results and Discussion
3.1. Molecular Weight Detection
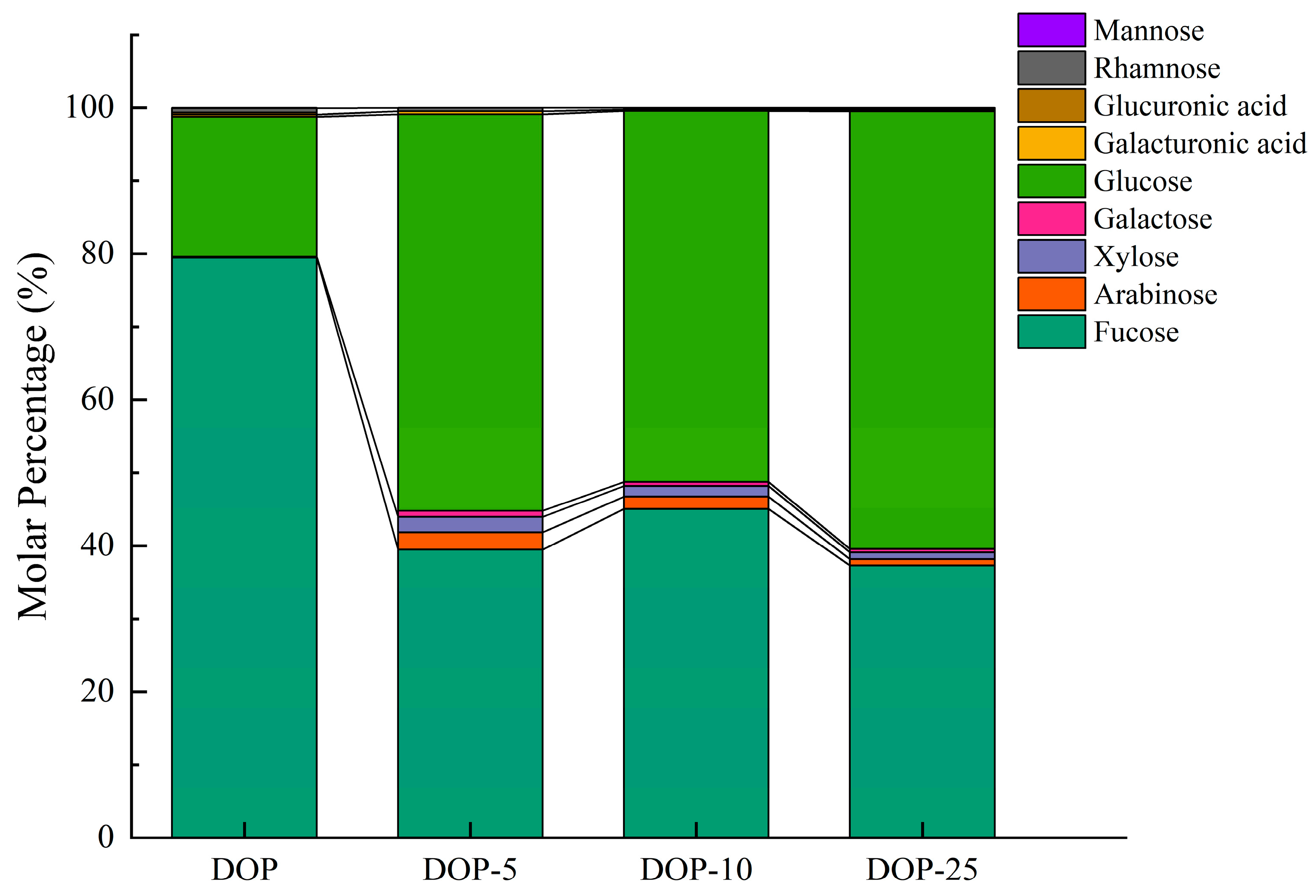
3.2. Polysaccharide Composition
3.3. C. elegans Assays
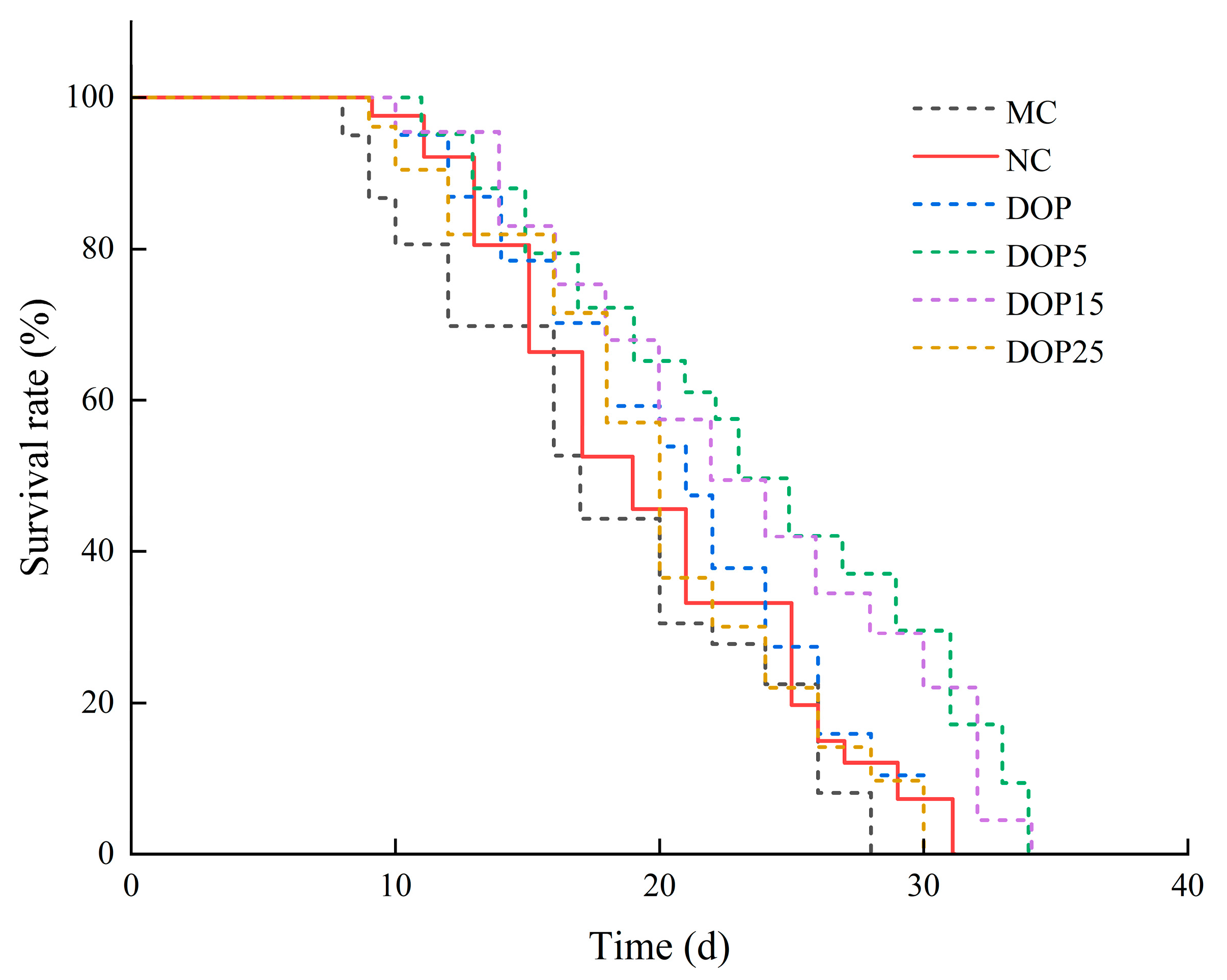
3.3.1. Effects of Polysaccharides on the Lifespan of C. elegans
3.3.2. Effects of Polysaccharides on the Locomotion Capacity of C. elegans
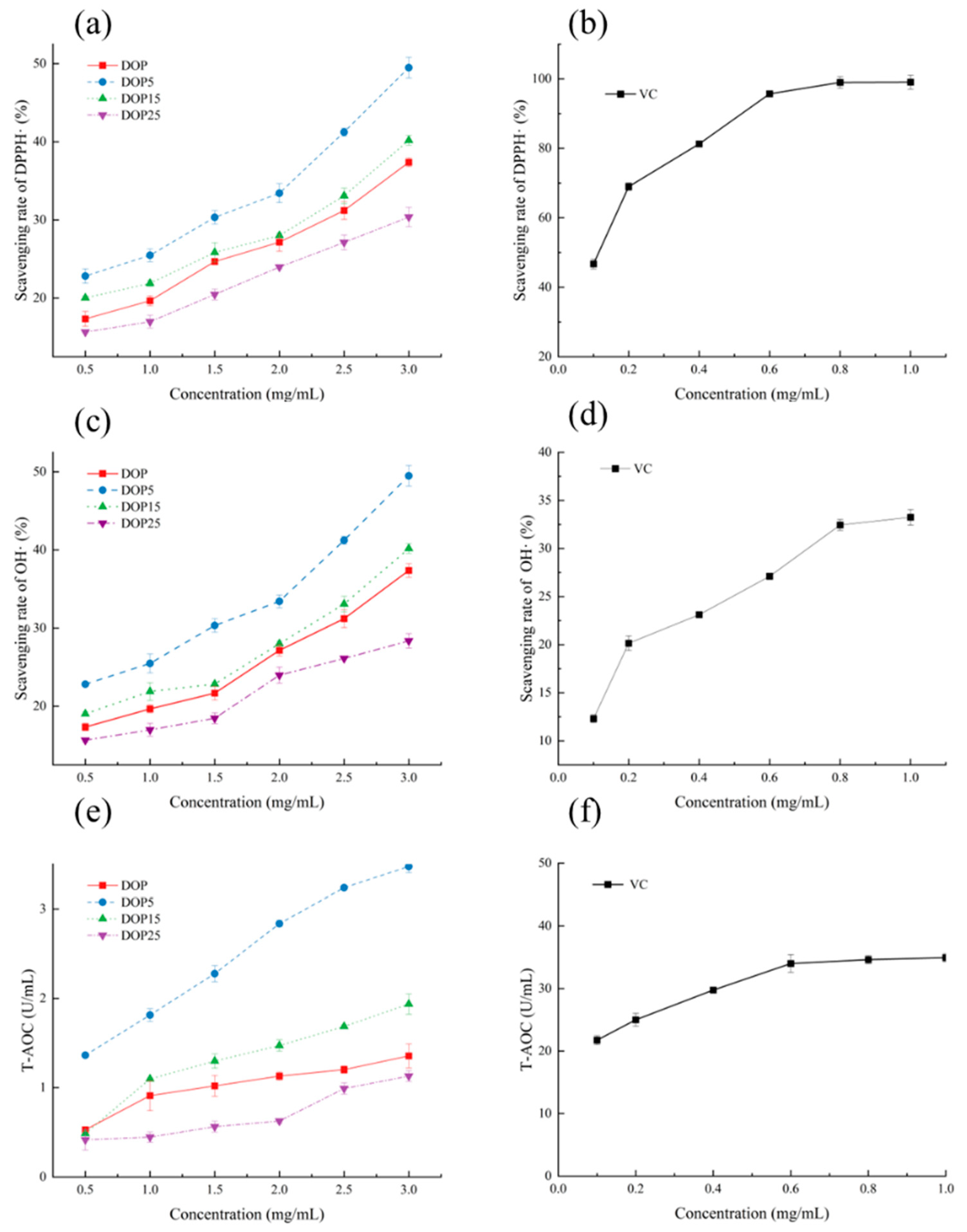
3.3.3. The Ability to Resist Oxidative Stress
3.3.4. Antioxidant Enzyme Activities in C. elegans
3.3.5. Effects of Polysaccharides of Different Molecular Weights in Fat Accumulation
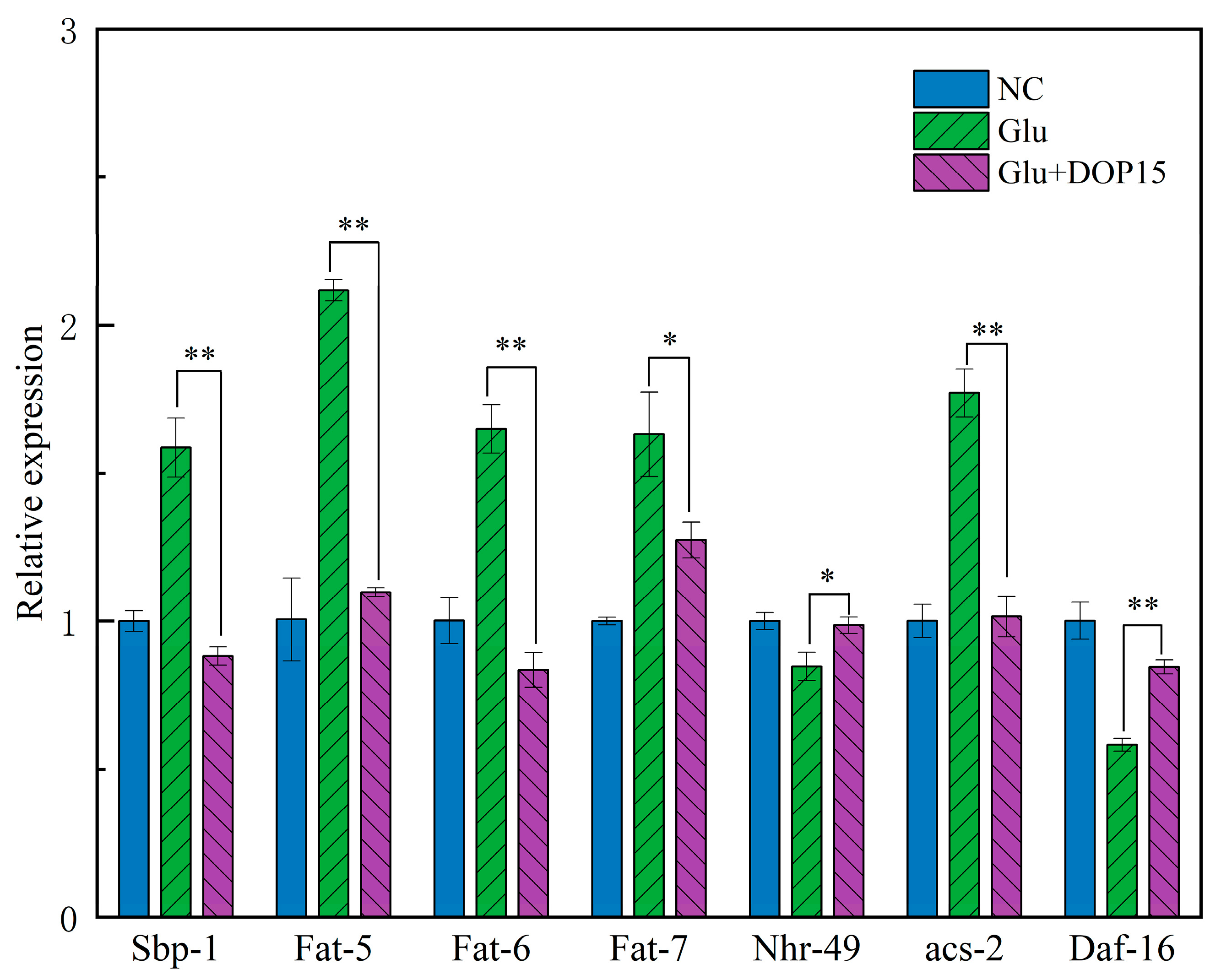
3.3.6. Effect of DOP15 on the Gene Expression of C. elegans
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moliner, C.; Núñez, S.; Cásedas, G.; Valero, M.S.; Dias, M.I.; Barros, L.; López, V.; Gómez-Rincón, C. Flowers of Allium cepa L. as Nutraceuticals: Phenolic Composition and Anti-Obesity and Antioxidant Effects in Caenorhabditis elegans. Antioxidants 2023, 12, 720. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrova, K.; Koelman, L.; Rodrigues, C.E. Dietary patterns and biomarkers of oxidative stress and inflammation: A systematic review of observational and intervention studies. Redox Biol 2021, 42, 101869. [Google Scholar] [CrossRef]
- Sandner, G.; König, A.; Wallner, M.; Weghuber, J. Functional foods—Dietary or herbal products on obesity: Application of selected bioactive compounds to target lipid metabolism. Curr. Opin. Food Sci. 2020, 34, 9–20. [Google Scholar] [CrossRef]
- Chen, W.-H.; Wu, J.-J.; Li, X.-F.; Lu, J.-M.; Wu, W.; Sun, Y.-Q.; Zhu, B.; Qin, L.-P. Isolation, structural properties, bioactivities of polysaccharides from Dendrobium officinale Kimura et. Migo: A review. Int. J. Biol. Macromol. 2021, 184, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Li, Y.; Tao, S.; Wei, G.; Huang, Y.; Chen, D.; Wu, C. Purification, characterization and biological activity of polysaccharides from Dendrobium officinale. Molecules 2016, 21, 701. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Trapika, I.G.S.C.; Tang, S.Y.S.; Cho, J.L.; Qi, Y.; Li, C.G.; Li, Y.; Yao, M.; Yang, D.; Liu, B.; et al. Mechanisms and active compounds polysaccharides and bibenzyls of medicinal Dendrobiums for diabetes management. Front. Nutr. 2022, 8, 811870. [Google Scholar] [CrossRef]
- Liu, Z.; Guo, Y.; Iu, J.J.; Si, J.; Wu, L.; Zhang, X. Effect of strains and parts on amino acids of Dendrobium officinale. Zhongguo Zhong Yao Za Zhi = Zhongguo Zhongyao Zazhi = China J. Chin. Mater. Medica 2015, 40, 1468–1472. [Google Scholar]
- Zhou, C.; Xie, Z.; Lei, Z.; Huang, Y.; Wei, G. Simultaneous identification and determination of flavonoids in Dendrobium officinale. Chem. Cent. J. 2018, 12, 40. [Google Scholar] [CrossRef]
- Liang, J.; Li, H.; Chen, J.; He, L.; Du, X.; Zhou, L.; Xiong, Q.; Lai, X.; Yang, Y.; Huang, S.; et al. Dendrobium officinale polysaccharides alleviate colon tumorigenesis via restoring intestinal barrier function and enhancing anti-tumor immune response. Pharmacol. Res. 2019, 148, 104417. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.; Han, J.; Shu, H.; Liu, K. Isolation of polysaccharides from Dendrobium officinale leaves and anti-inflammatory activity in LPS-stimulated THP-1 cells. Chem. Cent. J. 2018, 12, 40. [Google Scholar] [CrossRef]
- Kim, S.; Jo, K.; Byun, B.S.; Han, S.H.; Yu, K.-W.; Suh, H.J.; Hong, K.-B. Chemical and biological properties of puffed Dendrobium officinale extracts: Evaluation of antioxidant and anti-fatigue activities. J. Funct. Foods 2020, 73, 104144. [Google Scholar] [CrossRef]
- Wang, K.; Wang, H.; Liu, Y.; Shui, W.; Wang, J.; Cao, P.; Wang, H.; You, R.; Zhang, Y. Dendrobium officinale polysaccharide attenuates type 2 diabetes mellitus via the regulation of PI3K/Akt-mediated glycogen synthesis and glucose metabolism. J. Funct. Foods 2018, 40, 261–271. [Google Scholar] [CrossRef]
- Yang, L.-C.; Lu, T.-J.; Hsieh, C.-C.; Lin, W.-C. Characterization and immunomodulatory activity of polysaccharides derived from Dendrobium tosaense. Carbohydr. Polym. 2014, 111, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Qiu, D.; Liu, X.; Zhang, L.; Cai, S.; Zhang, X. Antiproliferative effect of Dendrobium catenatum Lindley polypeptides against human liver, gastric and breast cancer cell lines. Food Funct. 2015, 6, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Chen, S.; Hu, Y.; Yang, Y.; Yuan, J.; Wu, Y.; Li, S.; Lin, J.; He, L.; Hou, S.; et al. Protective roles and mechanisms of Dendrobium officinal polysaccharides on secondary liver injury in acute colitis. Int. J. Biol. Macromol. 2018, 107, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Miao, Y.; Xu, Y.; Meng, X.; Cui, W.; Wang, Y.; Zhu, L.; Sha, Z.; Wei, K.; Zhu, R. Taishan Pinus Massoniana pollen polysaccharide inhibits the replication of acute tumorigenic ALV-J and its associated tumor growth. Vet. Microbiol. 2019, 236, 108376. [Google Scholar] [CrossRef]
- Ren, D.; Wang, N.; Guo, J.; Yuan, L.; Yang, X. Chemical characterization of Pleurotus eryngii polysaccharide and its tumor-inhibitory effects against human hepatoblastoma HepG-2 cells. Carbohydr. Polym. 2016, 138, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Li, X.; Feng, S.; Liu, J.; Zhou, L.; Yuan, M.; Ding, C. Characteristics and bioactivities of different molecular weight polysaccharides from camellia seed cake. Int. J. Biol. Macromol. 2016, 91, 1025–1032. [Google Scholar] [CrossRef]
- Zhang, X.; Luo, Y.; Wei, G.; Li, Y.; Huang, Y.; Huang, J.; Liu, C.; Huang, R.; Liu, G.; Wei, Z.; et al. Physicochemical and Antioxidant Properties of the Degradations of Polysaccharides from Dendrobium officinale and Their Suitable Molecular Weight Range on Inducing HeLa Cell Apoptosis. Evid. Based Complement. Altern. Med. 2019, 2019, 4127360. [Google Scholar] [CrossRef]
- Tao, S.; Ren, Z.; Yang, Z.; Duan, S.; Wan, Z.; Huang, J.; Liu, C.; Wei, G. Effects of Different Molecular Weight Polysaccharides From Dendrobium officinale Kimura & Migo on Human Colorectal Cancer and Transcriptome Analysis of Differentially Expressed Genes. Front. Pharmacol. 2021, 12, 704486. [Google Scholar] [CrossRef]
- Yu, S.; Zheng, C.; Zhou, F.; Baillie, D.L.; Rose, A.M.; Deng, Z.; Chu, J.S.-C. Genomic identification and functional analysis of essential genes in Caenorhabditis elegans. BMC Genom. 2018, 19, 871. [Google Scholar] [CrossRef] [PubMed]
- Son, H.G.; Altintas, O.; Kim, E.J.E.; Kwon, S.; Lee, S.J.V. Age-dependent changes and biomarkers of aging in Caenorhabditis elegans. Aging Cell 2019, 18, e12853. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, Z.; Zhou, H.; Sun, X.; Chen, X.; Xu, N. The anti-aging effects of Gracilaria lemaneiformis polysaccharide in Caenorhabditis elegans. Int. J. Biol. Macromol. 2019, 140, 600–604. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, K.; Chang, F.Y.; Watts, J.L.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Ruvkun, G. Genome-wide RNAi analysis of Caenorhabditis elegans fat regulatory genes. Nature 2003, 421, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.-X.; Xie, X.S. Coherent Anti-Stokes Raman Scattering Microscopy: Instrumentation, Theory, and Applications. J. Phys. Chem. B 2003, 108, 827–840. [Google Scholar] [CrossRef]
- Meng, J.-K.; Zhan, N.; Wu, H.; Zhang, X.-Q. Optimization of high-pressure extraction of polysaccharide from Dendrobium candidum and analysis of its antioxidant activity. Food Mach. 2023, 39, 157–163. [Google Scholar]
- Sevag, M.; Lackman, D.B.; Smolens, J. The isolation of the components of streptococcal nucleoproteins in serologically active form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar] [CrossRef]
- Sun, Y.; Qin, Y.; Li, H.; Peng, H.; Chen, H.; Xie, H.-R.; Deng, Z. Rapid characterization of chemical constituents in Radix Tetrastigma, a functional herbal mixture, before and after metabolism and their antioxidant/antiproliferative activities. J. Funct. Foods 2015, 18, 300–318. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Fan, S.T.; Huang, D.F.; Yu, Q.; Liu, X.Z.; Li, C.; Wang, S.; Xiong, T.; Nie, S.P.; Xie, M.Y. Effect of Lactobacillus plantarum NCU116 fermentation on Asparagus officinalis polysaccharide: Characterization, antioxidative, and immunoregulatory activities. J. Agr. Food Chem. 2018, 66, 10703–10711. [Google Scholar] [CrossRef]
- Shi, H.; Hu, X.; Zheng, H.; Li, C.; Sun, L.; Guo, Z.; Huang, W.; Yu, R.; Song, L.; Zhu, J. Two novel antioxidant peptides derived from Arca subcrenata against oxidative stress and extend lifespan in Caenorhabditis elegans. J. Funct. Foods 2021, 81, 104462. [Google Scholar] [CrossRef]
- Yang, Z.-Z.; Yu, Y.-T.; Lin, H.-R.; Liao, D.-C.; Cui, X.-H.; Wang, H.-B. Lonicera japonica extends lifespan and healthspan in Caenorhabditis elegans. Free. Radic. Biol. Med. 2018, 129, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Wang, W.; Chu, W. Antioxidant and reducing lipid accumulation effects of rutin in Caenorhabditis elegans. BioFactors 2021, 47, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ching, T. Oil Red O Staining for Lipid Content in Caenorhabditis elegans. Bio-Protoc. 2021, 11, e4124. [Google Scholar] [CrossRef] [PubMed]
- Farias-Pereira, R.; Oshiro, J.; Kim, K.-H.; Park, Y. Green coffee bean extract and 5-O-caffeoylquinic acid regulate fat metabolism in Caenorhabditis elegans. J. Funct. Foods 2018, 48, 586–593. [Google Scholar] [CrossRef]
- Zheng, H.; Cui, S.; Sun, B.; Zhang, B.; Tao, D.; Wang, Z.; Zhang, Y.; Ma, F. Synergistic effect of discrete ultrasonic and H2O2 on physicochemical properties of chitosan. Carbohydr. Polym. 2022, 291, 119598. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Chen, F.; Yang, W.; Huang, H. Preparation, deproteinization and comparison of bioactive polysaccharides. Trends Food Sci. Technol. 2021, 109, 564–568. [Google Scholar] [CrossRef]
- Chen, F.; Huang, S.; Huang, G. Preparation, activity, and antioxidant mechanism of rice bran polysaccharide. Food Funct. 2021, 12, 834–839. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Yang, W.; Huang, G.; Huang, H. The antioxidant activities of balsam pear polysaccharide. Int. J. Biol. Macromol. 2020, 142, 232–236. [Google Scholar] [CrossRef]
- Wei, Q.; Zhang, Y.-H. Ultrasound-assisted polysaccharide extraction from Cercis chinensis and properites, antioxidant activity of polysaccharide. Ultrason. Sonochem. 2023, 96, 106422. [Google Scholar] [CrossRef]
- Georgiev, V.G.; Weber, J.; Kneschke, E.-M.; Denev, P.N.; Bley, T.; Pavlov, A.I. Antioxidant Activity and Phenolic Content of Betalain Extracts from Intact Plants and Hairy Root Cultures of the Red Beetroot Beta vulgaris cv. Detroit Dark Red. Plant Foods Hum. Nutr. 2010, 65, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Alcántar-Fernández, J.; González-Maciel, A.; Reynoso-Robles, R.; Andrade, M.E.P.; Hernández-Vázquez, A.d.J.; Velázquez-Arellano, A.; Miranda-Ríos, J. High-glucose diets induce mitochondrial dysfunction in Caenorhabditis elegans. PLoS ONE 2019, 14, e0226652. [Google Scholar] [CrossRef] [PubMed]
- Fitzenberger, E.; Boll, M.; Wenzel, U. Impairment of the proteasome is crucial for glucose-induced lifespan reduction in the mev-1 mutant of Caenorhabditis elegans. Biochim. Et Biophys. Acta (BBA)—Mol. Basis Dis. 2013, 1832, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Zhao, Z.; Bi, J.; Dai, H.; He, Y.; Zhang, W.; Wang, J.; Shu, Z.; Xiao, A. Effects of high-sugar diet on development and antioxidant capacity of Drosophila melano-gaster and its mechanism. Sci. Technol. Food Ind. 2021, 42, 377–384. [Google Scholar]
- Fang, Z.; Xiao, B.; Jiang, W.; Hao, X.; Tan, J.; Lu, A.; Li, J.; Wang, W.; Wang, G.; Zhang, Y. The antioxidant capacity evaluation of polysaccharide hydrolyzates from pumpkin using Caenorhabditis elegans model. J. Food Biochem. 2020, 45, e13275. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zou, Y.; Fang, Q.; Feng, R.; Zhang, J.; Zhou, W.; Wei, Q. Polysaccharides from Brasenia schreberi with Great Antioxidant Ability and the Potential Application in Yogurt. Molecules 2023, 29, 150. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tang, M.; Zhao, X.-R.; Feng, S.-L.; Liu, L.; Zhou, L.-J.; Cao, X.-H.; Huang, Y.; Yang, H.-Y.; Ding, C.-B. Antioxidant potential evaluation of polysaccharides from Camellia oleifera Abel in vitro and in vivo. Int. J. Biol. Macromol. 2023, 248, 125726. [Google Scholar] [CrossRef]
- Farias-Pereira, R.; Savarese, J.; Yue, Y.; Lee, S.-H.; Park, Y. Fat-lowering effects of isorhamnetin are via NHR-49-dependent pathway in Caenorhabditis elegans. Curr. Res. Food Sci. 2019, 2, 70–76. [Google Scholar] [CrossRef]
- Wang, K.; Chen, S.; Zhang, C.; Huang, J.; Wu, J.; Zhou, H.; Jin, L.; Qian, X.; Jin, J.; Lyu, J. Enhanced ROS production leads to excessive fat accumulation through DAF-16 in Caenorhabditis elegans. Exp. Gerontol. 2018, 112, 20–29. [Google Scholar] [CrossRef]
- Barathikannan, K.; Tyagi, A.; Shan, L.; Kim, N.-H.; Lee, D.-S.; Park, J.-S.; Chelliah, R.; Oh, D.-H. Antiobesity and Antioxidative Effect of Fermented Brown Rice Using In Vitro with In Vivo Caenorhabditis elegans Model. Life 2023, 13, 374. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, Y.; Yang, Z.; Liu, X.; Fan, F. Study on the processing technology and antioxidant properties of Dendrobium officinale and apple compound beverages. Storage Process 2021, 21, 93–98. [Google Scholar]
- Liu, D.; Yin, X.; Yi, L.; Li, Y. Research on the technology of dendrobium officinale milk functional jelly. Food Ferment. Ind. 2014, 50, 99–102. [Google Scholar]
- Luo, M.; Xie, W. Development of Dendrobium officinale leaf health-care tea bag. Food Ind. 2021, 42, 38–43. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Primer (5′-3′) |
|---|---|---|
| Sbp-1 | F | TGGCGTTCCAATCTATTCGC |
| R | CTTGTGGGTTGGCTCCGTT | |
| Fat-5 | F | TCGGAGAAGGAGGTCACAAC |
| R | TCCCGTTCAGTTTCACAGCC | |
| Fat-6 | F | GCGCTGCTCACTATTTCGGA |
| R | TGGAAGTTGTGACCTCCCTC | |
| Fat-7 | F | TGTTTCACACTTCACGCCAC |
| R | CCTCCTTCACCAACGGCTAC | |
| Nhr-49 | F | ATCAGATGCCAGATGACGCA |
| R | TGCTGTAAAGAGACCGGAGC | |
| acs-2 | F | GCGGAGCACATTAAGAAGGC |
| R | TGACAGTTCCGAGACCCAAC | |
| Daf-16 | F | TACCGGGTGCCTATGGAAAC |
| R | AGAGCCGATGAAGAAGCGAC |
| Groups | Average Lifespan (d) | Increase (%) | Maximum Lifespan (d) |
|---|---|---|---|
| MC | 17.38 ± 0.85 # | - | 28.47 ± 1.01 # |
| NC | 19.12 ± 0.55 | - | 30.61 ± 1.22 |
| DOP | 21.10 ± 0.50 ** | 10.32 | 30.00 ± 0.77 |
| DOP5 | 23.66 ± 1.36 ** | 23.73 | 34.65 ± 0.92 ** |
| DOP15 | 22.02 ± 0.58 ** | 15.19 | 33.90 ± 0.73 ** |
| DOP25 | 20.09 ± 0.25 ** | 5.06 | 31.36 ± 1.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, X.; Wang, H.; Guan, C.; Chen, Q.; Cui, X.; Zhang, X. Impact of Molecular Weight Variations in Dendrobium officinale Polysaccharides on Antioxidant Activity and Anti-Obesity in Caenorhabditis elegans. Foods 2024, 13, 1040. https://doi.org/10.3390/foods13071040
Pang X, Wang H, Guan C, Chen Q, Cui X, Zhang X. Impact of Molecular Weight Variations in Dendrobium officinale Polysaccharides on Antioxidant Activity and Anti-Obesity in Caenorhabditis elegans. Foods. 2024; 13(7):1040. https://doi.org/10.3390/foods13071040
Chicago/Turabian StylePang, Xiao, Heqin Wang, Chunbo Guan, Qiufeng Chen, Xinwen Cui, and Xiuqing Zhang. 2024. "Impact of Molecular Weight Variations in Dendrobium officinale Polysaccharides on Antioxidant Activity and Anti-Obesity in Caenorhabditis elegans" Foods 13, no. 7: 1040. https://doi.org/10.3390/foods13071040