
Immunomodulatory Effects of Sodium Hyaluronate Health Drink on Immunosuppressed Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Preparation of Sodium Hyaluronate Health Drink
2.3. Immunological Animal Experimental Design
2.4. Immunological Test Method
2.5. Animal Experimental Design for Intestinal Flora
2.6. 16S rRNA Sequencing Method
2.7. Statistical Analysis
3. Results
3.1. Body Weight and Organ Index of Mice
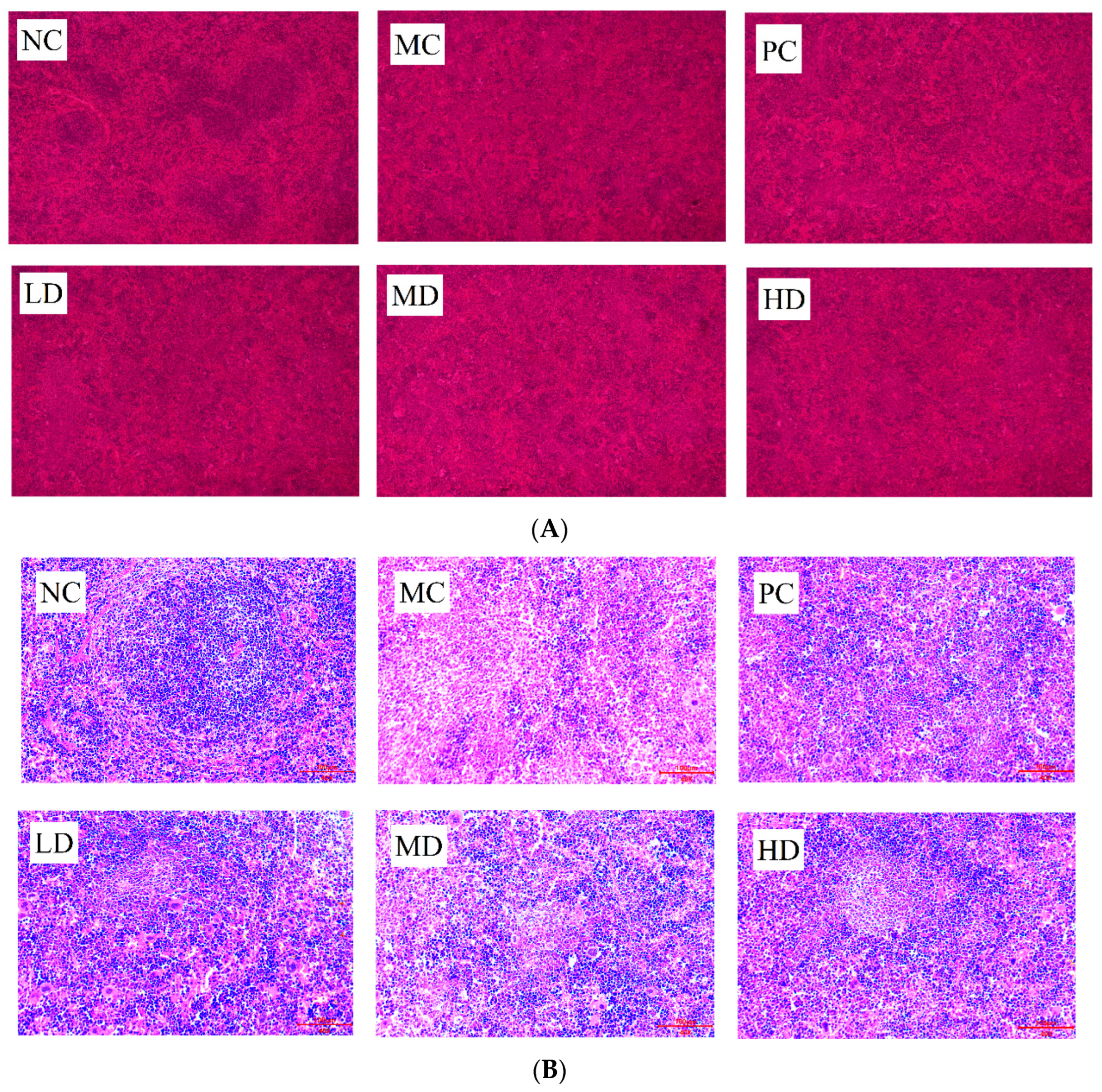
3.2. Spleen Histomorphology of Mice
3.3. Immunology Experiments
3.4. Serum Levels of Immune Factors
3.5. Dilution Curve and Alpha and Beta Diversity
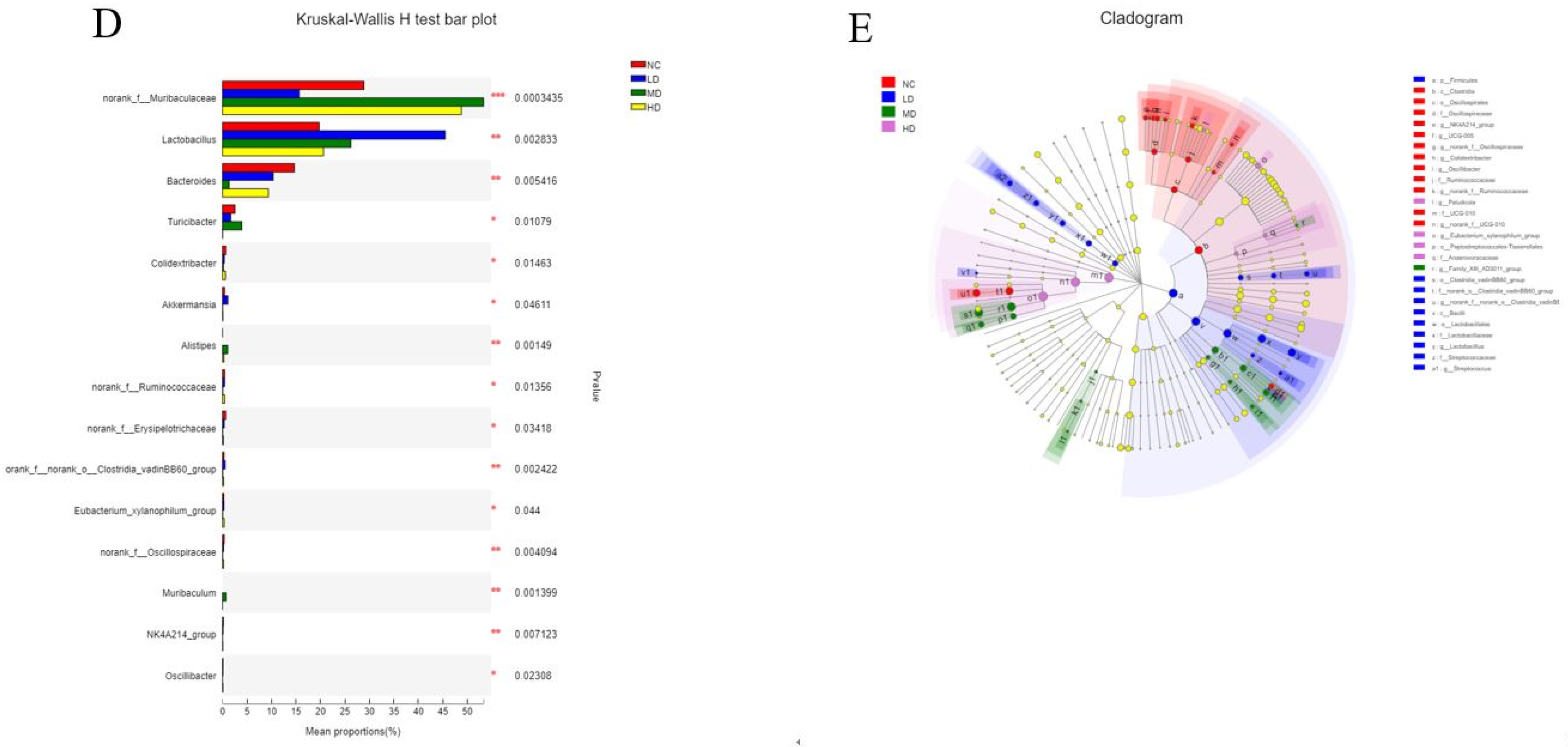
3.6. Species Composition Analysis
3.7. Species Variation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Gong, Y.; Wu, J.; Li, S.T. Immuno-enhancement effects of Lycium ruthenicum Murr. polysaccharide on cyclophosphamide-induced immunosuppression in mice. Int. J. Clin. Exp. Med. 2015, 8, 20631–20637. [Google Scholar] [PubMed]
- Chen, X.; Nie, W.; Fan, S. A polysaccharide from Sargassum fusiforme protects against immunosuppression in cyclophosphamide-treated mice. Carbohydr. Polym. Sci. Technol. Asp. Ind. Important Polysacch. 2012, 90, 1114–1119. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Duan, S.; Li, Y.; Pan, X.; Han, L. Polysaccharides in natural products that repair the damage to intestinal mucosa caused by cyclophosphamide and their mechanisms: A review. Carbohydr. Polym. 2021, 261, 117876. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yan, Y.; Zhou, W.; Chen, D.; Cao, Y. Effects of polysaccharides from bee collected pollen of Chinese wolfberry on immune response and gut microbiota composition in cyclophosphamide-treated mice. J. Funct. Foods 2020, 72, 104057. [Google Scholar] [CrossRef]
- Bai, Y.; Huang, F.; Zhang, R.; Dong, L.; Zhang, M. Longan pulp polysaccharides relieve intestinal injury in vivo and in vitro by promoting tight junction expression. Carbohydr. Polym. 2019, 229, 115475. [Google Scholar] [CrossRef]
- Xie, A.; Wan, H.; Feng, L.; Yang, B.; Wan, Y. Protective Effect of Anoectochilus formosanus Polysaccharide against Cyclophosphamide-Induced Immunosuppression in BALB/c Mice. Foods 2023, 12, 1910. [Google Scholar] [CrossRef]
- Wang, W.; Xu, C.; Liu, Z.; Gu, L.; Ma, J.; Hou, J.; Jiang, Z. Physicochemical properties and bioactivity of polysaccharides from Isaria cicadae Miquel with different extraction processes: Effects on gut microbiota and immune response in mice. Food Funct. 2022, 13, 9268–9284. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, J.; Zhao, Q.; Wen, A.; Li, L.; Zhang, Y. The regulating effect of Tibet Opuntia ficusindica (Linn.) Mill. polysaccharides on the intestinal flora of cyclophosphamide-induced immunocompromised mice. Int. J. Biol. Macromol. 2022, 207, 570–579. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, G.; Wen, Y.; Liu, S.; Li, W. Intestinal microbiota are involved in the immunomodulatory activities of longan polysaccharide. Mol. Nutr. Food Res. 2017, 61, 1700466. [Google Scholar] [CrossRef]
- Liu, J.P.; Wang, J.; Zhou, S.X.; Huang, D.C.; Qi, G.H.; Chen, G.T. Ginger polysaccharides enhance intestinal immunity by modulating gut microbiota in cyclophosphamide-induced immunosuppressed mice. Int. J. Biol. Macromol. 2022, 223, 1308–1319. [Google Scholar] [CrossRef]
- Ying, M.; Yu, Q.; Zheng, B.; Wang, H.; Wang, J.; Chen, S.; Nie, S.; Xie, M. Cultured Cordyceps sinensis polysaccharides modulate intestinal mucosal immunity and gut microbiota in cyclophosphamide-treated mice. Carbohydr. Polym. 2020, 235, 115957. [Google Scholar] [CrossRef]
- Zhu, Z.; Luo, Y.; Lin, L.; Gao, T.; Yang, Q.; Fan, Y.; Wang, S.; Fu, C.; Liao, W. Modulating Effects of Turmeric Polysaccharides on Immune Response and Gut Microbiota in Cyclophosphamide-Treated Mice. J. Agric. Food Chem. 2024, 72, 3469–3482. [Google Scholar] [CrossRef]
- Meyer, K.; Palmer, J.W. The Polysaccharide of the Vitreous Humor. J. Biol. Chem. 1934, 107, 629–634. [Google Scholar] [CrossRef]
- Tokita, Y.; Sakashita, H.; Okamoto, A.; Kubota, K. Kinetic study of a radical scavenging effect of hyaluronic acid. Polym. Int. 2010, 38, 161–164. [Google Scholar] [CrossRef]
- Sara, S.; Maryam, B. Daily consumption of the collagen supplement Pure Gold Collagen reduces visible signs of aging. Clin. Interv. Aging 2014, 9, 1747–1758. [Google Scholar]
- Brandt, K.D.; Block, J.A.; Michalski, J.P.; Moreland, L.W.; Caldwell, J.R.; Lavin, P.T. Efficacy and safety of intraarticular sodium hyaluronate in knee osteoarthritis. ORTHOVISC Study Group. Clin. Orthop. Relat. Res. 2001, 385, 130. [Google Scholar] [CrossRef]
- Cole, B.J.; Karas, V.; Hussey, K.; Pilz, K.; Fortier, L.A. Hyaluronic Acid Versus Platelet-Rich Plasma: A Prospective, Double-Blind Randomized Controlled Trial Comparing Clinical Outcomes and Effects on Intra-articular Biology for the Treatment of Knee Osteoarthritis. Am. J. Sports Med. 2016, 45, 339–346. [Google Scholar] [CrossRef]
- Huang, G.; Huang, H. Application of hyaluronic acid as carriers in drug delivery. Drug Deliv. 2018, 25, 766–772. [Google Scholar] [CrossRef]
- Park, B.G.; Park, Y.S.; Park, J.W.; Shin, E.; Shin, W.S. Anti-obesity potential of enzymatic fragments of hyaluronan on high-fat diet-induced obesity in C57BL/6 mice. Biochem. Biophys. Res. Commun. 2016, 473, 290–295. [Google Scholar] [CrossRef]
- Petricevic, B.; Wessner, B.; Sachet, M.; Vrbanec, D.; Bergmann, M. CL097, a TLR7/8 ligand, inhibits TLR-4—Dependent activation of IRAK-M and BCL-3 expression. Shock 2009, 32, 484–490. [Google Scholar] [CrossRef]
- Miyazaki, M.; Yuba, E.; Hayashi, H.; Harada, A.; Kono, K. Development of pH-Responsive Hyaluronic Acid-Based Antigen Carriers for Induction of Antigen-Specific Cellular Immune Responses. ACS Biomater. Sci. Eng. 2019, 5, 5790–5797. [Google Scholar] [CrossRef] [PubMed]
- Chadwick, W.; Magnus, T.; Martin, B.; Keselman, A.; Mattson, M.P.; Maudsley, S. Targeting TNF-α receptors for neurotherapeutics. Trends Neurosci. 2008, 31, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Charles, P.; Elliott, M.J.; Davis, D.; Potter, A.; Kalden, J.R.; Antoni, C.; Breedveld, F.C.; Smolen, J.S.; Eberl, G.; de Woody, K.; et al. Regulation of Cytokines, Cytokine Inhibitors, and Acute-Phase Proteins Following Anti-TNF-α Therapy in Rheumatoid Arthritis. J. Immunol. 1999, 163, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Ritchlin, C.T.; Haas-Smith, S.A.; Li, P.; Hicks, D.G.; Schwarz, E.M. Mechanisms of TNF-alpha- and RANKL-mediated osteoclastogenesis and bone resorption in psoriatic arthritis. J. Clin. Investig. 2003, 111, 821–831. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.A.; Tam, C.S.; Stanhope, K.L.; Wolfe, B.M.; Ali, M.R.; Majella, O.K.; O’Keeffe, M.; St-Onge, M.-P.; Ravussin, E.; Havel, P.J. Low Circulating Adropin Concentrations with Obesity and Aging Correlate with Risk Factors for Metabolic Disease and Increase after Gastric Bypass Surgery in Humans. J. Clin. Endocrinol. Metab. 2012, 97, 3783–3791. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila gen. Nov., sp. Nov., a human intestinal mucin-degrading bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef]
- Zi-Feng, S.U.; Zhi-Ming, D.; Jian-Fa, Y. Research Progress on Immune Mechanism of Polysaccharide. J. Yunnan Agric. Univ. 2006, 21, 205–209. [Google Scholar]
- Wang, J.; Tong, X.; Li, P.; Cao, H.; Su, W. Immuno-enhancement effects of Shenqi Fuzheng Injection on cyclophosphamide-induced immunosuppression in Balb/c mice. J. Ethnopharmacol. 2012, 139, 788–795. [Google Scholar]
- Zhao, Y.L.; Wang, J.B.; Shan, L.M.; Jin, C.; Ma, L.; Xiao, X.H. Effect of Radix isatidis polysaccharides on immunological function and expression of immune related cytokines in mice. Chin. J. Integr. Med. 2008, 14, 207–211. [Google Scholar] [CrossRef]
- Yan, H.; Lu, J.; Wang, Y.; Gu, W.; Yang, X.; Yu, J. Intake of total saponins and polysaccharides from Polygonatum kingianum affects the gut microbiota in diabetic rats. Phytomedicine 2017, 26, 45. [Google Scholar] [CrossRef]
- Fang, Q.; Hu, J.; Nie, Q.; Nie, S. Effects of polysaccharides on glycometabolism based on gut microbiota alteration. Trends Food Sci. Technol. 2019, 92, 65–70. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Marín, M.L.; Martín, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernández, L.; Rodríguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Solís, G.; Reyes-Gavilan CG, D.L.; Fernández, N.; Margolles, A.; Gueimonde, M. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe 2010, 16, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Salzman, N.H. The potter’s wheel: The host’s role in sculpting its microbiota. Cell. Mol. Life Sci. 2011, 68, 3675–3685. [Google Scholar] [CrossRef]
- Koboziev, I.; Reinoso Webb, C.; Furr, K.L.; Grisham, M.B. Role of the enteric microbiota in intestinal homeostasis and inflammation. Free Radic. Biol. Med. 2014, 68, 122–133. [Google Scholar] [CrossRef]
- Oe, M.; Mitsugi, K.; Odanaka, W.; Yoshida, H.; Matsuoka, R.; Seino, S.; Kanemitsu, T.; Masuda, Y. Dietary hyaluronic acid migrates into the skin of rats. Sci. World J. 2014, 2014, 378024. [Google Scholar] [CrossRef]
- Cobo, F.; Foronda, C.; Pérez-Carrasco, V.; Martin-Hita, L.; Navarro-Marí, J.M. First description of abdominal infection due to Alistipes onderdonkii. Anaerobe 2020, 66, 102283. [Google Scholar] [CrossRef]
- Yuan, X.; Chen, R.; Zhang, Y.; Lin, X.; Mccormick, K.L. Gut Microbiota of Chinese Obese Children and Adolescents with and Without Insulin Resistance. Front. Endocrinol. 2021, 12, 636272. [Google Scholar] [CrossRef]
- Derrien, M.; Van Baarlen, P.; Hooiveld, G.; Norin, E.; Müller, M.; De Vos, W.M. Modulation of Mucosal Immune Response, Tolerance, and Proliferation in Mice Colonized by the Mucin-Degrader Akkermansia muciniphila. Front. Microbiol. 2011, 2, 166. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Spleen Index (mg/g) | Thymus Index (mg/g) |
|---|---|---|
| NC | 4.01 ± 0.03 | 2.54 ± 0.01 * |
| MC | 3.85 ± 0.04 | 1.67 ± 0.02 |
| PC | 4.39 ± 0.06 | 1.93 ± 0.01 |
| LD | 4.08 ± 0.02 | 1.91 ± 0.01 |
| MD | 4.40 ± 0.04 | 2.07 ± 0.03 * |
| HD | 4.25 ± 0.03 | 2.06 ± 0.02 * |
| Sample | Richness | Diversity | Coverage | ||
|---|---|---|---|---|---|
| Ace | Chao | Shannon | Simpson | ||
| NC | 252.59 ± 17.55 | 255.59 ± 23.59 | 3.19 ± 0.31 | 0.09 ± 0.02 | 0.9992 |
| LD | 213.25 ± 20.09 ** | 217.33 ± 24.75 * | 2.52 ± 0.13 ** | 0.21 ± 0.05 ** | 0.9993 |
| MD | 227.87 ± 25.23 | 230.46 ± 30.10 | 2.67 ± 0.11 ** | 0.14 ± 0.02 * | 0.9990 |
| HD | 207.86 ± 18.13 ** | 206.81 ± 17.24 ** | 2.60 ± 0.41 ** | 0.16 ± 0.05 ** | 0.9992 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Yao, X.; Liu, Y.; Peng, B. Immunomodulatory Effects of Sodium Hyaluronate Health Drink on Immunosuppressed Mice. Foods 2024, 13, 842. https://doi.org/10.3390/foods13060842
Peng X, Yao X, Liu Y, Peng B. Immunomodulatory Effects of Sodium Hyaluronate Health Drink on Immunosuppressed Mice. Foods. 2024; 13(6):842. https://doi.org/10.3390/foods13060842
Chicago/Turabian StylePeng, Xiaozhen, Xiaoqiao Yao, Ya Liu, and Bangzhu Peng. 2024. "Immunomodulatory Effects of Sodium Hyaluronate Health Drink on Immunosuppressed Mice" Foods 13, no. 6: 842. https://doi.org/10.3390/foods13060842