
Updated Parameters for Listeria monocytogenes Dose–Response Model Considering Pathogen Virulence and Age and Sex of Consumer
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dose–Response Model
2.2. Inference Process
- A classification of L. monocytogenes strains as a function of their virulence;
- Estimates of the inter-individual variability in susceptibility within each sub-population and estimates of the inter-strain variability in virulence within each class of virulence;
- Estimates of the exposure of the considered sub-populations to the various classes of virulence;
- Estimates of the corresponding number of cases of invasive listeriosis for each combination of sub-populations/class of virulence.
2.2.1. Classification of Strain Virulence
2.2.2. Estimate of the Standard Deviation of log10(r) within a Sub-Population/Class of Virulence
2.2.3. Proportion of Various Classes of Strain Virulence in EU RTE and Clinical Cases
2.2.4. Exposure of the EU Population to Various Classes of Virulence
2.2.5. Prevalence of Invasive Listeriosis in the EU Population according to Classes of Virulence
2.3. Scaling of the Model to Epidemiological Data
- Estimates of the number of invasive human listeriosis cases in the EU from 2008 to 2015 within 14 sub-populations for 3 different classes of virulence (42 categories);
- An estimate of the prevalence of contaminated RTE and the ecdf of the exposure to L. monocytogenes from contaminated servings of RTE for each of these 42 categories;
- Estimates of the standard deviation of the lognormal (base 10) distribution of r for 3 classes of virulence, considered as similar in each sub-population for a given class of virulence.
2.4. Implementation and Diffusion
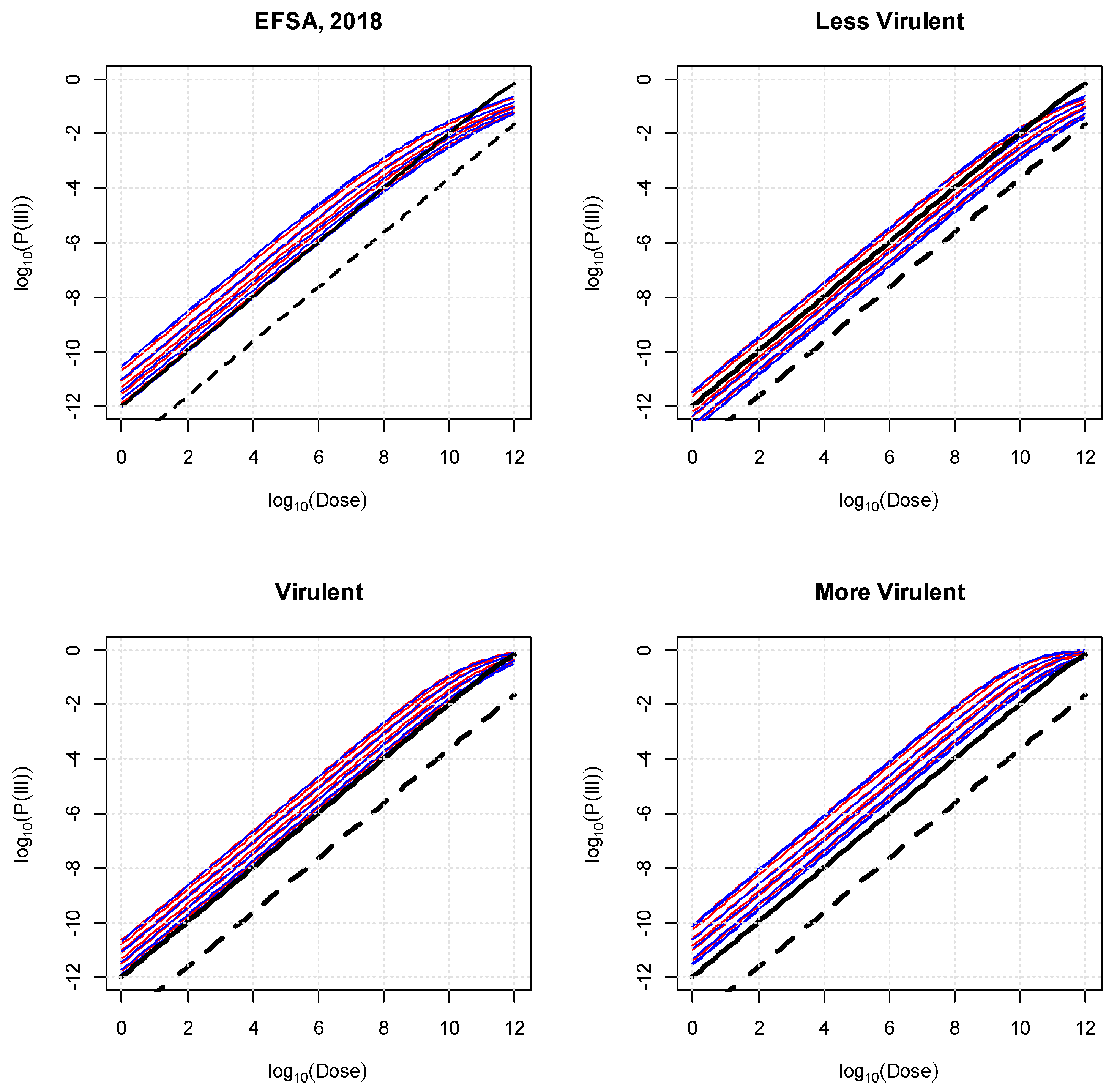
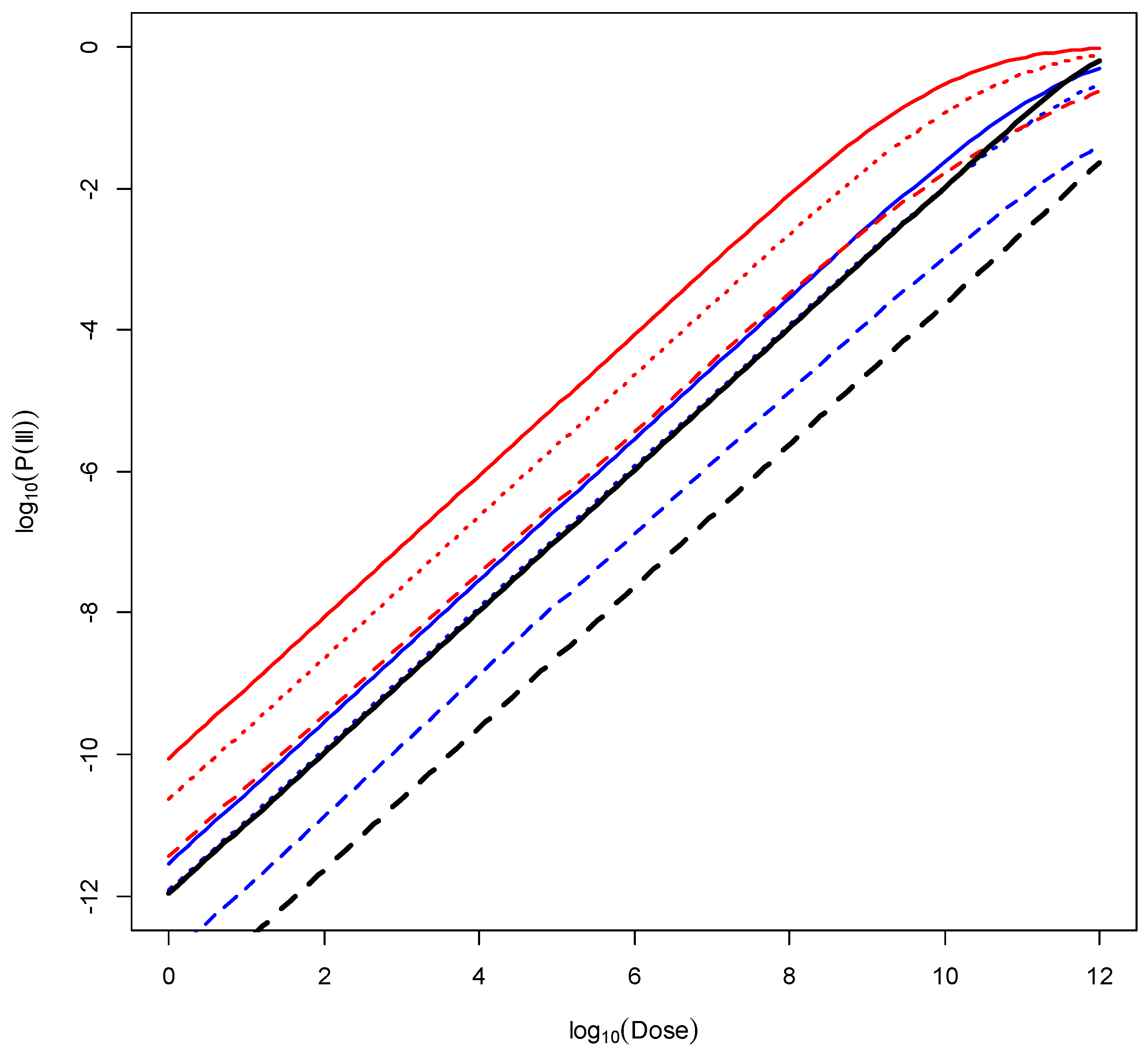
3. Results
- (i)
- Susceptible FAO/WHO population versus the most susceptible population (females > 75 years old) yields an RR of 22.0;
- (ii)
- General FAO/WHO population versus least susceptible population (males between 15 and 24 years old) yields an RR of 50.1.
- (iii)
- The RR between the least susceptible group (males between 15 and 24 years old) exposed to the “less virulent” class of strains versus the most susceptible group (females > 75 years old) exposed to the most virulent class of strains is 655.1.
- (iv)
- Taking class of strain virulence and age–sex sub-populations into account results in a wider range of possible probability of illness at a specific dose (RR = 655) compared with the range obtained from the previous FAO and WHO model (RR = 45).
4. Discussion
- Pouillot et al. [8] modeled the r-value as a log10-normal distribution. The host’s underlying health conditions, such as pregnancy, cancer, heart disease, etc., were incorporated using RRs for various sub-populations and a proportion of each sub-population estimated from the French population [13]. Exposure to L. monocytogenes was estimated from US data [19]. No growth from retail to consumption was considered. For the MPD, an important parameter in L. monocytogenes risk assessment [21], a value of 6.1 log10 cfu/g was used;
- Fritsch et al. [10] estimated r-values for 26 strains of L. monocytogenes and then grouped these strains into three classes of virulence according to the similarity of the r-value, namely, “hypovirulent” (called here “less virulent”), “medium virulent” (called here “virulent”), and “hypervirulent” (called here “more virulent”);
- The EFSA [9] estimated r-values for 14 age–sex combinations. Age and sex were used as surrogates for underlying health conditions because the demographic stratification into the 14 sub-populations was readily available across the EU, unlike actual information on underlying health conditions. The EFSA considered a bacterial growth from retail to consumption and MPDs ranging from 6.23 to 7.53 log10 cfu/g according to the considered RTE [9].
- In 2022, an FAO and WHO expert group [11] considered that the most appropriate DR model approaches would include variability in sub-population susceptibility and variability in strain virulence. The availability of an exposure model to L. monocytogenes in the EU [9] and of CC data from representative samples of food and clinical cases in the same population [16] made this derivation possible.
- The novelty of the current work is to conflate these three works to develop a DR model for invasive listeriosis that could consider differences in strain virulence class as well as better account for host susceptibility through the inclusion of surrogate information—age and sex. Consequently, this new model allows for more specific estimation of the risk of invasive listeriosis at the cost of two additional pieces of information, namely, population demographics and proportional contribution of strain virulence classes in food products of interest.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar] [CrossRef]
- FAO; WHO. Microbiological Risk Assessment Guidance for Food; Microbiological Risk Assessment Series, No 36; Food and Agriculture Organization of the United Nations: Roma, Italy; World Health Organization: Geneva, Switzerland, 2021; p. 288. [Google Scholar]
- Buchanan, R.L.; Smith, J.L.; Long, W. Microbial Risk Assessment: Dose-Response Relations and Risk Characterization. Int. J. Food Microbiol. 2000, 58, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Hoelzer, K.; Chen, Y.; Dennis, S.; Evans, P.; Pouillot, R.; Silk, B.J.; Walls, I. New Data, Strategies, and Insights for Listeria monocytogenes Dose-Response Models: Summary of an Interagency Workshop, 2011. Risk Anal. 2013, 33, 1568–1581. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Risk Assessment of Listeria monocytogenes in Ready to Eat Foods; Technical Report; Microbiological Risk Assessment Series, No 5; Food and Agriculture Organization of the United Nations: Roma, Italy; World Health Organization: Geneva, Switzerland, 2004; p. 269. [Google Scholar]
- Haas, C.N.; Rose, J.B.; Gerba, C.P. Quantitative Microbial Risk Assessment; Wiley: New York, NY, USA, 1999. [Google Scholar]
- Buchanan, R.L.; Damert, W.G.; Whiting, R.C.; van Schothorst, M. Use of Epidemiologic and Food Survey Data to Estimate a Purposefully Conservative Dose-Response Relationship for Listeria monocytogenes Levels and Incidence of Listeriosis. J. Food Prot. 1997, 60, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Pouillot, R.; Hoelzer, K.; Chen, Y.; Dennis, S.B. Listeria monocytogenes Dose Response Revisited—Incorporating Adjustments for Variability in Strain Virulence and Host Susceptibility. Risk Anal. 2015, 35, 90–108. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Listeria monocytogenes Contamination of Ready-to-Eat Foods and the Risk for Human Health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Fritsch, L.; Guillier, L.; Augustin, J.-C. Next Generation Quantitative Microbiological Risk Assessment: Refinement of the Cold Smoked Salmon-Related Listeriosis Risk Model by Integrating Genomic Data. Microb. Risk Anal. 2018, 10, 20–27. [Google Scholar] [CrossRef]
- FAO; WHO. Listeria monocytogenes in Ready-to-Eat (RTE) Foods: Attribution, Characterization and Monitoring—Meeting Report; Microbiological Risk Assessment Series, No 38; Food and Agriculture Organization of the United Nations: Roma, Italy; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Haas, C.N. Conditional Dose-Response Relationships for Microorganisms: Development and Application. Risk Anal. 2002, 22, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Goulet, V.; Hebert, M.; Hedberg, C.; Laurent, E.; Vaillant, V.; De Valk, H.; Desenclos, J.C. Incidence of Listeriosis and Related Mortality among Groups at Risk of Acquiring Listeriosis. Clin. Infect. Dis. 2012, 54, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Roche, S.M.; Velge, P.; Bottreau, E.; Durier, C.; Marquet-van der Mee, N.; Pardon, P. Assessment of the Virulence of Listeria monocytogenes: Agreement between a Plaque-Forming Assay with HT-29 Cells and Infection of Immunocompetent Mice. Int. J. Food Microbiol. 2001, 68, 33–44. [Google Scholar] [CrossRef] [PubMed]
- FDA; FSIS. Quantitative Assessment of Relative Risk to Public Health from Foodborne Listeria monocytogenes among Selected Categories of Ready-to-Eat Foods; Food and Drug Administration, United States: College Park, MD, USA; Department of Agriculture: Washington, DC, USA; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2003; p. 541.
- Møller Nielsen, E.; Björkman, J.T.; Kiil, K.; Grant, K.; Dallman, T.; Painset, A.; Amar, C.; Roussel, S.; Guillier, L.; Félix, B.; et al. Closing Gaps for Performing a Risk Assessment on Listeria monocytogenes in Ready-to-eat (RTE) Foods: Activity 3, the Comparison of Isolates from Different Compartments along the Food Chain, and from Humans Using Whole Genome Sequencing (WGS) Analysis. EFSA Support. Publ. 2017, 14, 1151E. [Google Scholar] [CrossRef]
- European Food Safety Authority. Analysis of the Baseline Survey on the Prevalence of Listeria monocytogenes in Certain Ready-to-Eat Foods in the EU, 2010–2011 Part A: Listeria Monocytogenes Prevalence Estimates. EFSA J. 2013, 11, 3241. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, F.; Carrasco, E.; Bover-Cid, S.; Jofré, A.; Valero, A. Closing Gaps for Performing a Risk Assessment on Listeria monocytogenes in Ready-to-Eat (RTE) Foods: Activity 2, a Quantitative Risk Characterization on L. monocytogenes in RTE Foods; Starting from the Retail Stage. EFSA Support. Publ. 2017, 14, 1252E. [Google Scholar] [CrossRef]
- Gombas, D.E.; Chen, Y.; Clavero, R.S.; Scott, V.N. Survey of Listeria monocytogenes in Ready-to-Eat Foods. J. Food Prot. 2003, 66, 559–569. [Google Scholar] [CrossRef]
- R Development Core Team, R. A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 1 February 2024).
- Pouillot, R.; Lubran, M.B. Predictive Microbiology Models vs. Modeling Microbial Growth within Listeria monocytogenes Risk Assessment: What Parameters Matter and Why. Food Microbiol. 2011, 28, 720–726. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Guidance of the Scientific Committee on Transparency in the Scientific Aspects of Risk Assessments Carried out by EFSA. Part 2: General Principles. EFSA J. 2009, 7, 1051. [Google Scholar] [CrossRef]
- Pouillot, R.; Goulet, V.; Delignette-Muller, M.L.; Mahe, A.; Cornu, M. Quantitative Risk Assessment of Listeria monocytogenes in French Cold-Smoked Salmon: II. Risk Characterization. Risk Anal. 2009, 29, 806–819. [Google Scholar] [CrossRef]
- Liao, J.; Guo, X.; Li, S.; Anupoju, S.M.B.; Cheng, R.A.; Weller, D.L.; Sullivan, G.; Zhang, H.; Deng, X.; Wiedmann, M. Comparative Genomics Unveils Extensive Genomic Variation between Populations of Listeria Species in Natural and Food-Associated Environments. ISME Commun. 2023, 3, 85. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Chen, W.; Wang, J.; Xu, B.; Liu, H.; Dong, Q.; Zhang, X. 10-Year Molecular Surveillance of Listeria Monocytogenes Using Whole-Genome Sequencing in Shanghai, China, 2009–2019. Front. Microbiol. 2020, 11, 551020. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Less Virulent Strains | Virulent Strains | More Virulent Strains |
|---|---|---|
| CC121, CC204, CC31, CC9, CC193, CC19, ST214 | CC14, CC155, CC177, CC18, CC20, CC21, CC26, CC3, CC37, CC379, C388, CC398, CC5, CC59, CC8, CC403 and all others | CC1, CC101, CC2, CC220, CC224, CC4, CC451, CC54, CC6, CC7, CC87 |
| Less Virulent | Virulent | More Virulent | Others/Unknown | n | |
|---|---|---|---|---|---|
| RTE Seafood | 149 (51.4%) | 102 (35.2%) | 36 (12.4%) | 3 (1.0%) | 290 |
| RTE Meats | 104 (59.1%) | 35 (19.9%) | 35 (19.9%) | 2 (1.1%) | 176 |
| RTE cheese and dairy | 11 (12.4%) | 42 (47.2%) | 29 (32.6%) | 7 (7.9%) | 89 |
| Sporadic | 22 (8.4%) | 76 (29.0%) | 156 (59.5%) | 8 (3.1%) | 262 |
| Population | Less Virulent (σ = 1.247) | Virulent (σ = 0.836) | More Virulent (σ = 0.756) |
|---|---|---|---|
| Female 1–4 yo | −14.166 (4.22 × 10−13) | −12.296 (3.22 × 10−12) | −11.671 (9.71 × 10−12) |
| Male 1–4 yo | −14.124 (4.65 × 10−13) | −12.256 (3.54 × 10−12) | −11.625 (1.08 × 10−11) |
| Female 5–14 yo | −14.516 (1.89 × 10−13) | −12.582 (1.67 × 10−12) | −12.046 (4.09 × 10−12) |
| Male 5–14 yo | −14.633 (1.44 × 10−13) | −12.690 (1.30 × 10−12) | −12.165 (3.11 × 10−12) |
| Female 15–24 yo | −14.002 (6.45 × 10−13) | −12.123 (4.80 × 10−12) | −11.521 (1.37 × 10−11) |
| Male 15–24 yo | −14.668 (1.33 × 10−13) | −12.730 (1.19 × 10−12) | −12.200 (2.87 × 10−12) |
| Female 25–44 yo | −13.708 (1.21 × 10−12) | −11.815 (9.76 × 10−12) | −11.239 (2.63 × 10−11) |
| Male 25–44 yo | −14.444 (2.22 × 10−13) | −12.522 (1.92 × 10−12) | −11.973 (4.84 × 10−12) |
| Female 45–64 yo | −13.755 (1.09 × 10−12) | −11.890 (8.21 × 10−12) | −11.272 (2.43 × 10−11) |
| Male 45–64 yo | −13.753 (1.09 × 10−12) | −11.869 (8.61 × 10−12) | −11.274 (2.42 × 10−11) |
| Female 65–74 yo | −13.418 (2.36 × 10−12) | −11.594 (1.62 × 10−11) | −10.916 (5.52 × 10−11) |
| Male 65–74 yo | −13.283 (3.23 × 10−12) | −11.447 (2.28 × 10−11) | −10.785 (7.45 × 10−11) |
| Female >75 yo | −13.234 (3.61 × 10−12) | −11.437 (2.33 × 10−11) | −10.718 (8.70 × 10−11) |
| Male >75 yo | −13.255 (3.44 × 10−12) | −11.468 (2.17 × 10−11) | −10.734 (8.38 × 10−11) |
| Model | Population | Virulence | Dose (Bacteria) | Relative Risk | |||
|---|---|---|---|---|---|---|---|
| 102 | 103 | 106 | 109 | ||||
| FAO and WHO [5] | Healthy Population | All strains | 2.37 × 10−12 | 2.37 × 10−11 | 2.37 × 10−08 | 2.37 × 10−05 | Reference |
| Increased susceptibility | 1.06 × 10−10 | 1.06 × 10−09 | 1.06 × 10−06 | 1.06 × 10−03 | 44.7 | ||
| Pouillot et al. [8] | Less than 65 yo, no underlying condition | All strains | 8.15 × 10−10 | 8.14 × 10−09 | 7.33 × 10−06 | 2.74 × 10−03 | 343.5 |
| More than 65 yo, no underlying condition | 1.55 × 10−08 | 1.53 × 10−07 | 1.10 × 10−04 | 1.94 × 10−02 | 6473.8 | ||
| Pregnancy | 2.06 × 10−07 | 1.98 × 10−06 | 9.73 × 10−04 | 7.70 × 10−02 | 83,675.8 | ||
| Non-hematological cancer | 8.09 × 10−08 | 7.89 × 10−07 | 4.54 × 10−04 | 4.85 × 10−02 | 33,270.7 | ||
| Hematological Cancer | 9.66 × 10−07 | 8.93 × 10−06 | 3.18 × 10−03 | 1.50 × 10-01 | 376,694.8 | ||
| Renal or Liver Failure | 2.84 × 10−07 | 2.71 × 10−06 | 1.25 × 10−03 | 8.93 × 10−02 | 114,387.0 | ||
| Solid Organ Transplant | 3.18 × 10−07 | 3.03 × 10−06 | 1.37 × 10−03 | 9.40 × 10−02 | 127,857.9 | ||
| Inflammatory diseases | 8.66 × 10−08 | 8.44 × 10−07 | 4.81 × 10−04 | 5.03 × 10−02 | 35,604.0 | ||
| HIV/AIDS | 6.73 × 10−08 | 6.58 × 10−07 | 3.90 × 10−04 | 4.41 × 10−02 | 27,763.9 | ||
| Diabetes | 7.78 × 10−09 | 7.72 × 10−08 | 5.96 × 10−05 | 1.27 × 10−02 | 3259.4 | ||
| Heart Diseases | 5.26 × 10−09 | 5.23 × 10−08 | 4.18 × 10−05 | 9.90 × 10−03 | 2207.8 | ||
| EFSA [9] | Female 1–4 yo | All strains | 2.80 × 10−10 | 2.80 × 10−09 | 2.63 × 10−06 | 1.22 × 10−03 | 118.1 |
| Male 1–4 yo | 3.58 × 10−10 | 3.58 × 10−09 | 3.33 × 10−06 | 1.48 × 10−03 | 151.1 | ||
| Female 5–14 yo | 1.27 × 10−10 | 1.27 × 10−09 | 1.22 × 10−06 | 6.54 × 10−04 | 53.8 | ||
| Male 5–14 yo | 1.04 × 10−10 | 1.04 × 10−09 | 9.99 × 10−07 | 5.54 × 10−04 | 43.8 | ||
| Female 15–24 yo | 4.97 × 10−10 | 4.97 × 10−09 | 4.57 × 10−06 | 1.89 × 10−03 | 209.5 | ||
| Male 15–24 yo | 9.67 × 10−11 | 9.67 × 10−10 | 9.32 × 10−07 | 5.23 × 10−04 | 40.8 | ||
| Female 25–44 yo | 9.92 × 10−10 | 9.90 × 10−09 | 8.83 × 10−06 | 3.15 × 10−03 | 417.7 | ||
| Male 25–44 yo | 1.81 × 10−10 | 1.81 × 10−09 | 1.72 × 10−06 | 8.66 × 10−04 | 76.3 | ||
| Female 45–64 yo | 8.72 × 10−10 | 8.70 × 10−09 | 7.81 × 10−06 | 2.87 × 10−03 | 367.2 | ||
| Male 45–64 yo | 9.47 × 10−10 | 9.45 × 10−09 | 8.45 × 10−06 | 3.05 × 10−03 | 398.9 | ||
| Female 65–74 yo | 2.09 × 10−09 | 2.08 × 10−08 | 1.78 × 10−05 | 5.33 × 10−03 | 877.5 | ||
| Male 65–74 yo | 2.89 × 10−09 | 2.88 × 10−08 | 2.41 × 10−05 | 6.66 × 10−03 | 1215.8 | ||
| Female > 75 yo | 3.06 × 10−09 | 3.04 × 10−08 | 2.54 × 10−05 | 6.92 × 10−03 | 1284.7 | ||
| Male > 75 yo | 3.06 × 10−09 | 3.04 × 10−08 | 2.54 × 10−05 | 6.92 × 10−03 | 1284.7 | ||
| This Study | Female 1–4 yo | Less virulent | 4.22 × 10−11 | 4.22 × 10−10 | 4.22 × 10−07 | 3.77 × 10−04 | 17.8 |
| Male 1–4 yo | 4.65 × 10−11 | 4.65 × 10−10 | 4.65 × 10−07 | 4.12 × 10−04 | 19.6 | ||
| Female 5–14 yo | 1.89 × 10−11 | 1.89 × 10−10 | 1.88 × 10−07 | 1.75 × 10−04 | 8.0 | ||
| Male 5–14 yo | 1.44 × 10−11 | 1.44 × 10−10 | 1.44 × 10−07 | 1.35 × 10−04 | 6.1 | ||
| Female 15–24 yo | 6.15 × 10−11 | 6.15 × 10−10 | 6.14 × 10−07 | 5.35 × 10−04 | 26.0 | ||
| Male 15–24 yo | 1.33 × 10−11 | 1.33 × 10−10 | 1.33 × 10−07 | 1.25 × 10−04 | 5.6 | ||
| Female 25–44 yo | 1.21 × 10−10 | 1.21 × 10−09 | 1.21 × 10−06 | 9.96 × 10−04 | 51.2 | ||
| Male 25–44 yo | 2.22 × 10−11 | 2.22 × 10−10 | 2.22 × 10−07 | 2.05 × 10−04 | 9.4 | ||
| Female 45–64 yo | 1.09 × 10−10 | 1.09 × 10−09 | 1.09 × 10−06 | 9.02 × 10−04 | 45.9 | ||
| Male 45–64 yo | 1.09 × 10−10 | 1.09 × 10−09 | 1.09 × 10−06 | 9.05 × 10−04 | 46.0 | ||
| Female 65–74 yo | 2.36 × 10−10 | 2.36 × 10−09 | 2.36 × 10−06 | 1.80 × 10−03 | 99.7 | ||
| Male 65–74 yo | 3.23 × 10−10 | 3.23 × 10−09 | 3.21 × 10−06 | 2.37 × 10−03 | 136.1 | ||
| Female > 75 yo | 3.61 × 10−10 | 3.61 × 10−09 | 3.59 × 10−06 | 2.61 × 10−03 | 152.3 | ||
| Male > 75 yo | 3.44 × 10−10 | 3.44 × 10−09 | 3.42 × 10−06 | 2.50 × 10−03 | 144.9 | ||
| Female 1–4 yo | Virulent | 3.22 × 10−10 | 3.22 × 10−09 | 3.22 × 10−06 | 3.08 × 10−03 | 136.0 | |
| Male 1–4 yo | 3.54 × 10−10 | 3.54 × 10−09 | 3.54 × 10−06 | 3.37 × 10−03 | 149.2 | ||
| Female 5–14 yo | 1.67 × 10−10 | 1.67 × 10−09 | 1.67 × 10−06 | 1.63 × 10−03 | 70.4 | ||
| Male 5–14 yo | 1.30 × 10−10 | 1.30 × 10−09 | 1.30 × 10−06 | 1.27 × 10−03 | 54.9 | ||
| Female 15–24 yo | 4.80 × 10−10 | 4.80 × 10−09 | 4.80 × 10−06 | 4.52 × 10−03 | 202.6 | ||
| Male 15–24 yo | 1.19 × 10−10 | 1.19 × 10−09 | 1.19 × 10−06 | 1.17 × 10−03 | 50.1 | ||
| Female 25–44 yo | 9.76 × 10−10 | 9.76 × 10−09 | 9.76 × 10−06 | 8.80 × 10−03 | 412.0 | ||
| Male 25–44 yo | 1.92 × 10−10 | 1.92 × 10−09 | 1.92 × 10−06 | 1.86 × 10−03 | 80.9 | ||
| Female 45–64 yo | 8.21 × 10−10 | 8.21 × 10−09 | 8.20 × 10−06 | 7.48 × 10−03 | 346.2 | ||
| Male 45–64 yo | 8.61 × 10−10 | 8.61 × 10−09 | 8.61 × 10−06 | 7.83 × 10−03 | 363.3 | ||
| Female 65–74 yo | 1.62 × 10−09 | 1.62 × 10−08 | 1.62 × 10−05 | 1.40 × 10−02 | 685.3 | ||
| Male 65–74 yo | 2.28 × 10−09 | 2.28 × 10−08 | 2.28 × 10−05 | 1.89 × 10−02 | 960.5 | ||
| Female > 75 yo | 2.33 × 10−09 | 2.33 × 10−08 | 2.33 × 10−05 | 1.93 × 10−02 | 982.4 | ||
| Male > 75 yo | 2.17 × 10−09 | 2.17 × 10−08 | 2.17 × 10−05 | 1.81 × 10−02 | 916.1 | ||
| Female 1–4 yo | More Virulent | 9.71 × 10−10 | 9.71 × 10−09 | 9.71 × 10−06 | 9.06 × 10−03 | 409.6 | |
| Male 1–4 yo | 1.08 × 10−09 | 1.08 × 10−08 | 1.08 × 10−05 | 9.99 × 10−03 | 454.5 | ||
| Female 5–14 yo | 4.09 × 10−10 | 4.09 × 10−09 | 4.09 × 10−06 | 3.95 × 10−03 | 172.4 | ||
| Male 5–14 yo | 3.11 × 10−10 | 3.11 × 10−09 | 3.11 × 10−06 | 3.03 × 10−03 | 131.2 | ||
| Female 15–24 yo | 1.37 × 10−09 | 1.37 × 10−08 | 1.37 × 10−05 | 1.25 × 10−02 | 578.3 | ||
| Male 15–24 yo | 2.87 × 10−10 | 2.87 × 10−09 | 2.87 × 10−06 | 2.80 × 10−03 | 121.2 | ||
| Female 25–44 yo | 2.63 × 10−09 | 2.63 × 10−08 | 2.62 × 10−05 | 2.27 × 10−02 | 1107.6 | ||
| Male 25–44 yo | 4.84 × 10−10 | 4.84 × 10−09 | 4.84 × 10−06 | 4.65 × 10−03 | 204.0 | ||
| Female 45–64 yo | 2.43 × 10−09 | 2.43 × 10−08 | 2.43 × 10−05 | 2.12 × 10−02 | 1026.1 | ||
| Male 45–64 yo | 2.42 × 10−09 | 2.42 × 10−08 | 2.42 × 10−05 | 2.10 × 10−02 | 1020.0 | ||
| Female 65–74 yo | 5.52 × 10−09 | 5.52 × 10−08 | 5.52 × 10−05 | 4.33 × 10−02 | 2330.5 | ||
| Male 65–74 yo | 7.45 × 10−09 | 7.45 × 10−08 | 7.45 × 10−05 | 5.55 × 10−02 | 3145.5 | ||
| Female > 75 yo | 8.70 × 10−09 | 8.70 × 10−08 | 8.69 × 10−05 | 6.28 × 10−02 | 3669.7 | ||
| Male > 75 yo | 8.38 × 10−09 | 8.38 × 10−08 | 8.37 × 10−05 | 6.10 × 10−02 | 3536.8 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pouillot, R.; Kiermeier, A.; Guillier, L.; Cadavez, V.; Sanaa, M. Updated Parameters for Listeria monocytogenes Dose–Response Model Considering Pathogen Virulence and Age and Sex of Consumer. Foods 2024, 13, 751. https://doi.org/10.3390/foods13050751
Pouillot R, Kiermeier A, Guillier L, Cadavez V, Sanaa M. Updated Parameters for Listeria monocytogenes Dose–Response Model Considering Pathogen Virulence and Age and Sex of Consumer. Foods. 2024; 13(5):751. https://doi.org/10.3390/foods13050751
Chicago/Turabian StylePouillot, Régis, Andreas Kiermeier, Laurent Guillier, Vasco Cadavez, and Moez Sanaa. 2024. "Updated Parameters for Listeria monocytogenes Dose–Response Model Considering Pathogen Virulence and Age and Sex of Consumer" Foods 13, no. 5: 751. https://doi.org/10.3390/foods13050751