
Evaluation of the Potential Protective Effect of Ellagic Acid against Heavy Metal (Cadmium, Mercury, and Lead) Toxicity in SH-SY5Y Neuroblastoma Cells




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Cd, Hg, Pb, and EA on Cell Viability
2.2. Effect of EA on Cd-, Hg-, and Pb-Induced Cytotoxicity
2.3. Effect of EA on Cd2+-, Hg2+-, and Pb2+-Induced LDH Release
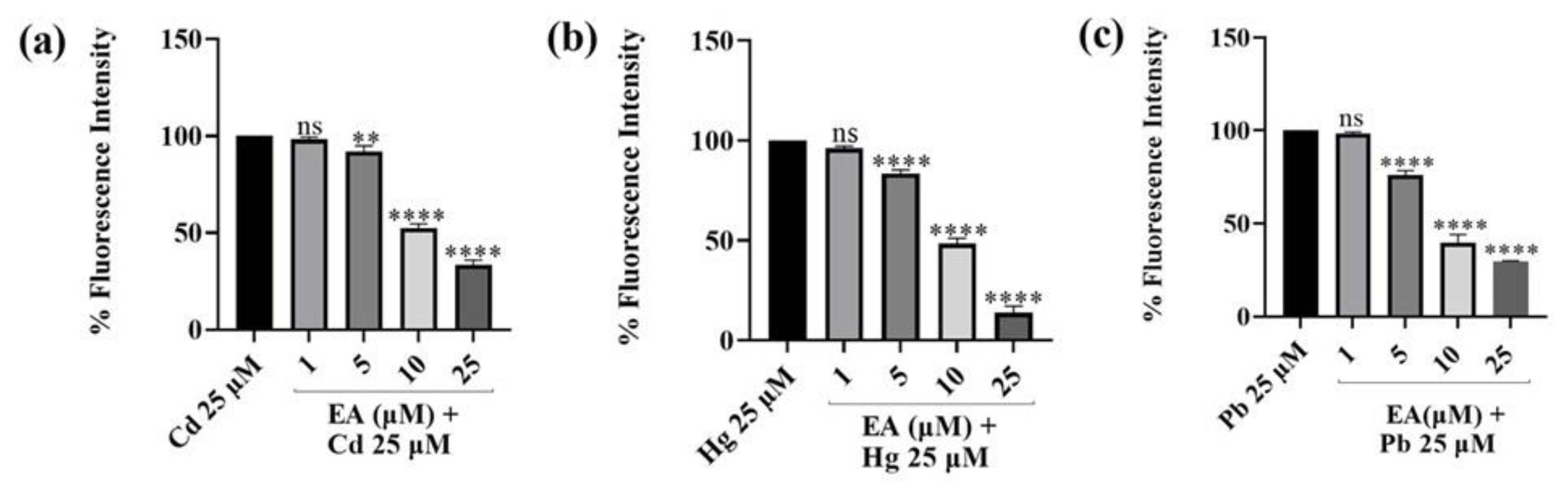
2.4. ROS Scavenging Effects of EA against Cd2+-, Hg2+-, Pb2+-, and H2O2-Induced Oxidative Stress
3. Discussion
3.1. Effect of Cd, Hg, Pb, and EA on Cell Viability
3.2. Effect of EA on Cd, Hg and Pb-Induced Cytotoxicity
3.3. Effect of EA on Cd2+-, Hg2+-, and Pb2+-Induced LDH Release
3.4. ROS Scavenging Effects of EA against Cd2+-, Hg2+-, Pb2+-, and H2O2-Induced Oxidative Stress
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Culture Cells
4.3. Determination of Cell Viability
4.4. Lactate Dehydrogenase (LDH) Release
4.5. Measurement of Intracellular ROS Production
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raz, N.; Daugherty, A.M. Pathways to brain aging and their modifiers: Free-radical-induced energetic and neural decline in senescence (FRIENDS) model—A mini-review. Gerontology 2017, 64, 49–57. [Google Scholar] [CrossRef]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Ijomone, O.M.; Ifenatuoha, C.W.; Aluko, O.M.; Ijomone, O.K.; Aschner, M. The aging brain: Impact of heavy metal neurotoxicity. Crit. Rev. Toxicol. 2020, 50, 801–814. [Google Scholar] [CrossRef]
- Karri, V.; Schuhmacher, M.; Kumar, V. Heavy metals (Pb, Cd, As and MeHg) as risk factors for cognitive dysfunction: A general review of metal mixture mechanism in brain. Environ. Toxicol. Pharmacol. 2016, 48, 203–213. [Google Scholar] [CrossRef]
- Belyaaeva, E.A.; Korotkov, S.M. Mechanism of primary Cd2+-induced rat liver mitochondria dysfunction: Discrete modes of Cd2+ action on calcium and thiol-dependent domains. Toxicol. Appl. Pharmacol. 2003, 192, 56–68. [Google Scholar] [CrossRef]
- He, X.; Qi, Z.; Hou, H.; Gao, J.; Zhang, X.X. Effects of chronic cadmium exposure at food limitation-relevant levels on energy metabolism in mice. J. Hazard. Mater. 2020, 388, 121791. [Google Scholar] [CrossRef] [PubMed]
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The effects of cadmium toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Peana, M.; Pelucelli, A.; Chasapis, C.T.; Perlepes, S.P.; Bekiari, V.; Medici, S.; Zoroddu, M.A. Biological Effects of Human Exposure to Environmental Cadmium. Biomolecules 2022, 13, 36. [Google Scholar] [CrossRef]
- Karthikeyan, J.; Bavani, G. Effect of cadmium on lactate dehydrogenase isoenzyme, succinate dehydrogenase and NA+-K+-ATPase in liver tissue of rat. J. Environ. Biol. 2009, 30, 895–898. [Google Scholar]
- Mazmanian, K.; Dudev, T.; Lim, C. How First Shell–Second Shell Interactions and Metal Substitution Modulate Protein Function. Inorg. Chem. 2018, 57, 14052–14061. [Google Scholar] [CrossRef]
- Branca, J.J.V.; Morucci, G.; Pacini, A. Cadmium-induced neurotoxicity: Still much ado. Neural. Regen. Res. 2018, 13, 1879–1882. [Google Scholar]
- Waalkes, M.P. Cadmium carcinogenesis. Mutat. Res. 2003, 533, 107–120. [Google Scholar] [CrossRef]
- Marrelli, M.; Argentieri, M.P.; Alexa, E.; Meleleo, D.; Statti, G.; Avato, P.; Conforti, F.; Mallamaci, R. Antioxidant activity and protective effect of the outer scales hydroalcoholic extract of Allium cepa L. var. Tropea on toxicity damage induced by Cadmium in Caco-2 cells. Food Chem. Toxicol. 2022, 170, 113495. [Google Scholar] [CrossRef]
- Cariccio, V.L.; Samà, A.; Bramanti, P.; Mazzon, E. Mercury involvement in neuronal damage and in neurodegenerative diseases. Biol. Trace Elem. Res. 2019, 187, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Carocci, A.; Rovito, N.; Sinicropi, M.S.; Genchi, G. Mercury toxicity and neurodegenerative effects. In Reviews of Environmental Contamination and Toxicology; Springer: Cham, Switzerland, 2014; pp. 1–18. [Google Scholar]
- Kehrer, J.P.; Klotz, L.O. Free radicals and related reactive species as mediators of tissue injury and disease: Implications for health. Crit. Rev. Toxicol. 2015, 45, 765–798. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Viso, P.; Domene, A.; Vélez, D.; Devesa, V.; Monedero, V.; Zúñiga, M. Oral exposure to inorganic mercury or methylmercury elicits distinct pro-inflammatory and pro-oxidant intestinal responses in a mouse model system. Food Chem. Toxicol. 2023, 177, 113801. [Google Scholar] [CrossRef] [PubMed]
- Mahboob, M.; Shireen, K.F.; Atkinson, A.; Khan, A.T. Lipid peroxidation and antioxidant enzyme activity in different organs of mice exposed to low level of mercury. J. Environ. Sci. Health Part B 2001, 36, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Aschner, M.; Syversen, T.; Souza, D.O.; Rocha, J.B.T.D.; Farina, M. Involvement of glutamate and reactive oxygen species in methylmercury neurotoxicity. Braz. J. Med. Biol. Res. 2007, 40, 285–291. [Google Scholar] [CrossRef]
- Ara, A.; Usmani, J.A. Lead toxicity: A review. Interdisci. Toxicol. 2015, 8, 55–64. [Google Scholar]
- Glass, T.A.; Bandeen-Roche, K.; McAtee, M.; Bolla, K.; Todd, A.C.; Schwartz, B.S. Neighborhood psychosocial hazards and the association of cumulative lead dose with cognitive function in older adults. Am. J. Epidemiol. 2009, 169, 683–692. [Google Scholar] [CrossRef]
- Pottier, G.; Viau, M.; Ricoul, M.; Shim, G.; Bellamy, M.; Cuceu, C.; Hempel, W.M.; Sabatier, L. Lead exposure induces telomere instability in human cells. PLoS ONE 2013, 8, e67501. [Google Scholar] [CrossRef]
- Carocci, A.; Catalano, A.; Lauria, G.; Sinicropi, M.S.; Genchi, G. Lead toxicity, antioxidant defense and environment. Rev. Environ. Contam. Toxicol. 2016, 235, 45–67. [Google Scholar]
- Virgolini, M.B.; Aschner, M. Chapter five—Molecular mechanisms of lead neurotoxicity. In Advances in neurotoxicology; Aschner, M., Costa, L.G., Eds.; Neurotoxicity of Metals: Old Issues and New Developments; Academic Press: Cambridge, MA, USA, 2021; Volume 5, pp. 159–213. [Google Scholar]
- Flora, S.J.S.; Shrivastava, R.; Mittal, M. Chemistry and pharmacological properties of some natural and synthetic antioxidants for heavy metal toxicity. Curr. Med. Chem. 2013, 20, 4540–4574. [Google Scholar] [CrossRef]
- Galano, A.; Francisco Marquez, M.; Pérez-González, A. Ellagic acid: An unusually versatile protector against oxidative stress. Chem. Res. Toxicol. 2014, 27, 904–918. [Google Scholar] [CrossRef]
- Harper, P. A Review of the Dietary Intake, Bioavailability and Health Benefits of Ellagic Acid (EA) with a Primary Focus on Its Anti-Cancer Properties. Cureus 2023, 15, e43156. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Lysiuk, R.; Oliinyk, P.; De Masi, L.; Bontempo, P.; et al. Ellagic acid: A review on its natural sources, chemical stability, and therapeutic potential. Oxidative Med. Cell. Longev. 2022, 2022, 3848084. [Google Scholar] [CrossRef] [PubMed]
- Durgun, C.; Aşir, F. Effect of ellagic acid on damage caused by hepatic ischemia reperfusion in rats. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 8209–8215. [Google Scholar] [PubMed]
- Fan, G.; Xu, Z.; Tang, J.; Liu, L.; Dai, R. Effect of ellagic acid on wound healing of chronic skin ulceration in diabetic mice and macrophage phenotype transformation. J. Biomater. Tissue Eng. 2019, 9, 1108–1113. [Google Scholar] [CrossRef]
- Mohammadinejad, A.; Mohajeri, T.; Aleyaghoob, G.; Heidarian, F.; Kazemi Oskuee, R. Ellagic acid as a potent anticancer drug: A comprehensive review on in vitro, in vivo, in silico, and drug delivery studies. Biotechnol. Appl. Biochem. 2022, 69, 2323–2356. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Ellagic acid in suppressing in vivo and in vitro oxidative stresses. Mol. Cell. Biochem. 2018, 448, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, K.I.; Khopde, S.M.; Kumar, S.S.; Mohan, H. Free radical studies of ellagic acid, a natural phenolic antioxidant. J. Agric. Food Chem. 2002, 50, 2200–2206. [Google Scholar] [CrossRef]
- Naraki, K.; Rahbardar, M.G.; Ajiboye, B.O.; Hosseinzadeh, H. The effect of ellagic acid on the metabolic syndrome: A review article. Heliyon 2023, 9, e21844. [Google Scholar] [CrossRef]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative stress, antioxidant capabilities, and bioavailability: Ellagic acid or urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Muthukumaran, S.; Tranchant, C.; Shi, J.; Ye, X.; Xue, S.J. Ellagic acid in strawberry (Fragaria spp.): Biological, technological, stability, and human health aspects. Food Qual. Saf. 2017, 1, 227–252. [Google Scholar] [CrossRef]
- Noori, M.; Jafari, B.; Hekmatdoost, A. Pomegranate juice prevents development of non-alcoholic fatty liver disease in rats by attenuating oxidative stress and inflammation. J. Sci. Food Agric. 2016, 97, 2327–2332. [Google Scholar] [CrossRef] [PubMed]
- Aslan, A.; Gok, O.; Beyaz, S.; Ağca, C.A.; Erman, O.; Zerek, A. Ellagic acid prevents kidney injury and oxidative damage via regulation of Nrf-2/NF-κB signaling in carbon tetrachloride induced rats. Mol. Bio. Rep. 2020, 47, 7959–7970. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.Z.; Zhu, G.F.; Zheng, C.Q.; Li, J.J.; Sheng, S.; Li, D.D.; Zhang, F. Ellagic acid protects dopamine neurons from rotenone-induced neurotoxicity via activation of Nrf2 signalling. J. Cell. Mol. Med. 2020, 24, 9446–9456. [Google Scholar] [CrossRef]
- Goudarzi, M.; Mombeini, M.A.; Fatemi, I.; Aminzadeh, A.; Kalantari, H.; Nesari, A.; Najafzadehvarzi, H.; Mehrzadi, S. Neuroprotective effects of Ellagic acid against acrylamide-induced neurotoxicity in rats. Neurol. Res. 2019, 41, 419–428. [Google Scholar] [CrossRef]
- Javaid, N.; Shah, M.A.; Rasul, A.; Chauhdary, Z.; Saleem, U.; Khan, H.; Ahmed, N.; Uddin, M.S.; Mathew, B.; Behl, T.; et al. Neuroprotective effects of Ellagic acid in Alzheimer’s disease: Focus on underlying molecular mechanisms of therapeutic potential. Curr. Pharm. Des. 2021, 27, 3591–3601. [Google Scholar] [CrossRef]
- Jamshidi, Z.; Roohbakhsh, A.; Karimi, G. An overview on the protective effects of ellagic acid against heavy metals, drugs, and chemicals. Food Sci. Nutr. 2023, 11, 7469–7484. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Kulkarni, V.H.; Chakraborty, M.; Habbu, P.V.; Ray, A. Ellagic acid restored lead-induced nephrotoxicity by anti-inflammatory, anti-apoptotic and free radical scavenging activities. Heliyon 2021, 7, e05921. [Google Scholar] [CrossRef] [PubMed]
- Meena, V.K.; Kumar, V.; Karalia, S.; Garima; Sundd, M. Ellagic Acid Modulates Uninduced as well as Mutation and Metal-Induced Aggregation of α-Synuclein: Implications for Parkinson’s Disease. ACS Chem. Neurosci. 2021, 12, 3598–3614. [Google Scholar] [CrossRef] [PubMed]
- Sudo, K.; Van Dao, C.; Miyamoto, A.; Shiraishi, M. Comparative analysis of in vitro neurotoxicity of methylmercury, mercury, cadmium, and hydrogen peroxide on SH-SY5Y cells. J. Vet. Med. Sci. 2019, 81, 828–837. [Google Scholar] [CrossRef]
- Anyanwu, B.O.; Ezejiofor, A.N.; Igweze, Z.N.; Orisakwe, O.E. Heavy metal mixture exposure and effects in developing nations: An update. Toxics 2018, 6, 65. [Google Scholar] [CrossRef]
- Meleleo, D.; Sblano, C.; Storelli, M.M.; Mallamaci, R. Evidence of cadmium and mercury involvement in the Aβ42 aggregation process. Biophys. Chem. 2020, 266, 106453. [Google Scholar] [CrossRef]
- Carmona, A.; Roudeau, S.; Ortega, R. Molecular Mechanisms of Environmental Metal Neurotoxicity: A Focus on the Interactions of Metals with Synapse Structure and Function. Toxics 2021, 9, 198. [Google Scholar] [CrossRef]
- Yousef, A.O.S.; Fahad, A.A.; Abdel Moneim, A.E.; Metwally, D.M.; El-Khadragy, M.F.; Kassab, R.B. The neuroprotective role of coenzyme Q10 against lead acetate-induced neurotoxicity is mediated by antioxidant, anti-inflammatory and anti-apoptotic activities. Int. J. Environ. Res. Public Health. 2019, 16, 2895. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, A.K.; Kumar, R.; Jamieson, S.; Pandey, A.K.; Bishayee, A. Neuroprotective potential of ellagic acid: A critical review. Adv. Nutr. 2021, 12, 1211–1238. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Núñez-Sánchez, M.Á.; Tomás-Barberán, F.A.; Espín, J.C. Neuroprotective Effects of Bioavailable Polyphenol-Derived Metabolites against Oxidative Stress-Induced Cytotoxicity in Human Neuroblastoma SH-SY5Y Cells. J. Agric. Food Chem. 2017, 65, 752–758. [Google Scholar] [CrossRef]
- Farbood, Y.; Sarkaki, A.; Dolatshahi, M.; Taqhi Mansouri, S.M.; Khodadadi, A. Ellagic acid protects the brain from 6-hydroxydopamine-induced neuroinflammation in a rat model of Parkinson’s disease. Basic Clin. Neurosci. 2015, 6, 83–89. [Google Scholar]
- Mannino, F.; Pallio, G.; Imbesi, C.; Scarfone, A.; Puzzolo, D.; Micali, A.; Freni, J.; Squadrito, F.; Bitto, A.; Minutoli, L.; et al. Beta-Caryophyllene, a Plant-Derived CB2 Receptor Agonist, Protects SH-SY5Y Cells from Cadmium-Induced Toxicity. Int. J. Mol. Sci. 2023, 24, 15487. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Huang, S. Cadmium activates the mitogen-activated protein kinase (MAPK) pathway via induction of reactive oxygen species and inhibition of protein phosphatases 2A and 5. Free Radic. Biol. Med. 2008, 45, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Suresh, C.; Dennis, A.O.; Heinz, J.; Vemuri, M.C.; Chetty, C.S. Melatonin protection against lead-induced changes in human neuroblastoma cell cultures. Int. J. Toxicol. 2006, 25, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Toimela, T.; Mäenpää, H.; Mannerström, M.; Tähti, H. Development of an in vitro blood–brain barrier model—Cytotoxicity of mercury and aluminum. Toxicol. Appl. Pharmacol. 2004, 195, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Méry, B.; Guy, J.B.; Vallard, A.; Espenel, S.; Ardail, D.; Rodriguez-Lafrasse, C.; Rancoule, C.; Magné, N. In Vitro Cell Death Determination for Drug Discovery: A Landscape Review of Real Issues. J. Cell Death 2017, 10, 1179670717691251. [Google Scholar] [CrossRef] [PubMed]
- Dalvi, L.T.; Moreira, D.C.; Andrade, R., Jr.; Ginani, J.; Alonso, A.; Hermes-Lima, M. Ellagic acid inhibits iron-mediated free radical formation. Spectrochim. Acta Part A 2017, 173, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Barbarossa, A.; Ceramella, J.; Carocci, A.; Iacopetta, D.; Rosato, A.; Limongelli, F.; Carrieri, A.; Bonofiglio, D.; Sinicropi, M.S. Benzothiazole-Phthalimide Hybrids as Anti-Breast Cancer and Antimicrobial Agents. Antibiotics 2023, 12, 1651. [Google Scholar] [CrossRef]
- Kaja, S.; Payne, A.J.; Naumchuk, Y.; Koulen, P. Quantification of lactate dehydrogenase for cell viability testing using cell lines and primary cultured astrocytes. Curr. Protoc. Toxicol. 2017, 72, 2–26. [Google Scholar] [CrossRef]
- Sun, A.; Chen, Y.; James-Kracke, M.; Wixom, P.; Cheng, Y. Ethanol-induced cell death by lipid peroxidation in PC12 cells. Neurochem. Res. 1997, 22, 1187–1192. [Google Scholar] [CrossRef]
- Carocci, A.; Catalano, A.; Bruno, C.; Lovece, A.; Roselli, M.G.; Cavalluzzi, M.M.; De Santis, F.; De Palma, A.; Rusciano, M.R.; Illario, M.; et al. N-(Phenoxyalkyl)amides as MT1 and MT2 ligands: Antioxidant properties and inhibition of Ca2+/CaM-dependent kinase II. Bioorg. Med. Chem. 2013, 21, 847–851. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallamaci, R.; Barbarossa, A.; Carocci, A.; Meleleo, D. Evaluation of the Potential Protective Effect of Ellagic Acid against Heavy Metal (Cadmium, Mercury, and Lead) Toxicity in SH-SY5Y Neuroblastoma Cells. Foods 2024, 13, 419. https://doi.org/10.3390/foods13030419
Mallamaci R, Barbarossa A, Carocci A, Meleleo D. Evaluation of the Potential Protective Effect of Ellagic Acid against Heavy Metal (Cadmium, Mercury, and Lead) Toxicity in SH-SY5Y Neuroblastoma Cells. Foods. 2024; 13(3):419. https://doi.org/10.3390/foods13030419
Chicago/Turabian StyleMallamaci, Rosanna, Alexia Barbarossa, Alessia Carocci, and Daniela Meleleo. 2024. "Evaluation of the Potential Protective Effect of Ellagic Acid against Heavy Metal (Cadmium, Mercury, and Lead) Toxicity in SH-SY5Y Neuroblastoma Cells" Foods 13, no. 3: 419. https://doi.org/10.3390/foods13030419