
A Long-Term Study on Chemical Compounds and Their Location in Sweet Basil Leaves from Organic and Conventional Producers

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Equipment
2.3. Plant Material
2.4. Chemical Analysis
2.4.1. Dry Matter Content
2.4.2. Hydrogen Peroxide Level Determination
2.4.3. Protein Extraction and Enzyme Activity Measurements
2.4.4. Total Polyphenol Content
2.4.5. Total Phenolic Acid Content
2.4.6. Total Flavonoid Content
2.4.7. Individual Phenolic Identification and Quantification
2.4.8. Total Chlorophyll and Carotenoid Contents
2.4.9. Individual Carotenoids and Chlorophylls Identification and Quantification
2.4.10. Antioxidant Activity: ABTS, DPPH, and FRAP
2.4.11. Microscopic Analyses
2.5. Statistical Analysis
3. Results and Discussion
3.1. Dry Matter in Basil
3.2. Enzymatic Status of Plants
3.3. Phenolic Compounds in Basil
3.4. Carotenoid Compounds in Plants
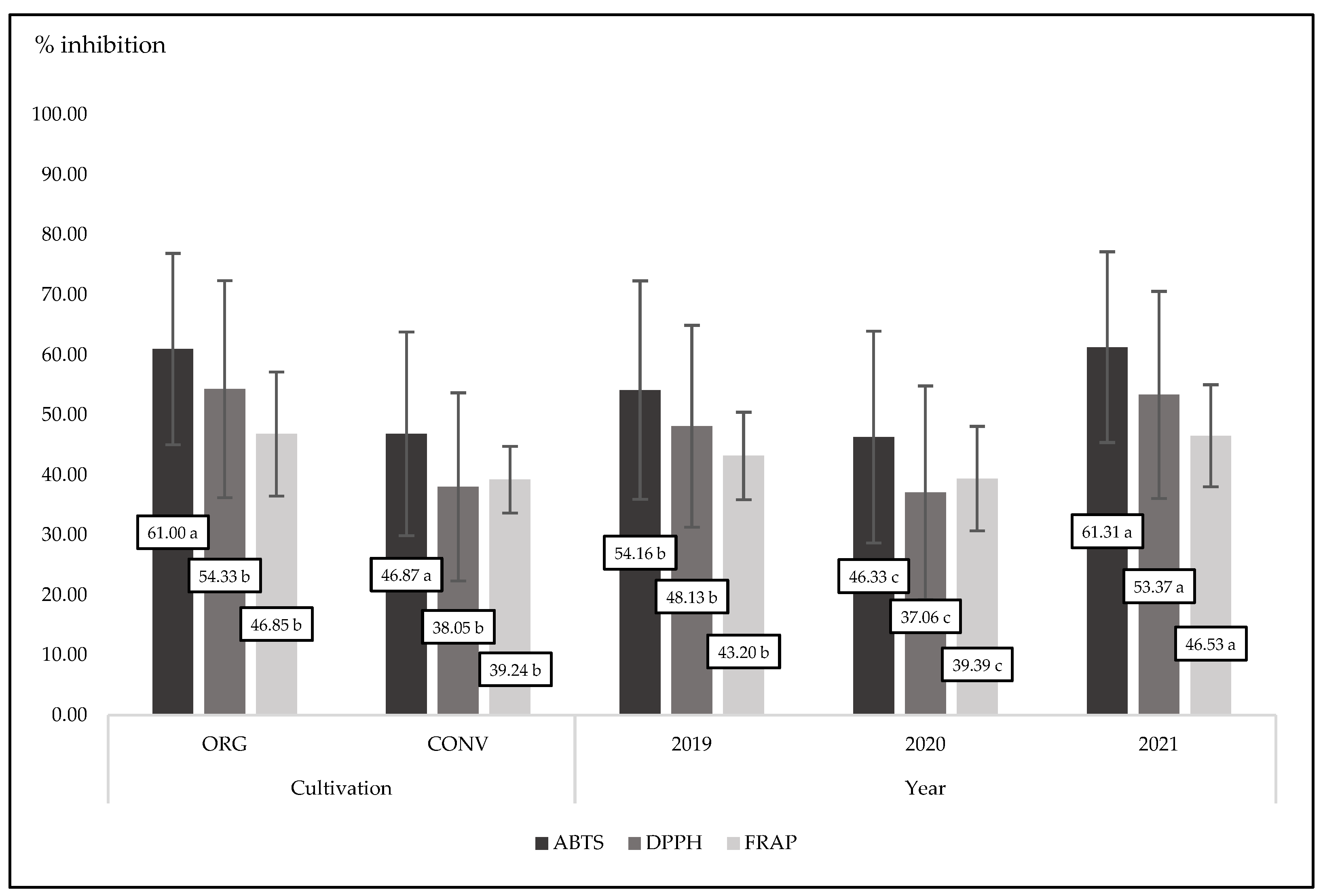
3.5. Antioxidant Status in Plants
3.6. Principal Component Analysis
3.7. Fluorescent Identification of Bioactive Compounds in Plants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Chemical components and pharmacological benefits of Basil (Ocimum basilicum): A review. Int. J. Food Prop. 2020, 23, 1961–1970. [Google Scholar] [CrossRef]
- Dudai, N.; Nitzan, N.; Gonda, I. Ocimum basilicum L. (Basil). In Medicinal, Aromatic and Stimulant Plants; Novak, J., Blüthner, W.-D., Eds.; Springer: Cham, Switzerland, 2020; Chapter 12. [Google Scholar]
- Teofilović, B.; Grujić-Letić, N.; Karadžić, M.; Kovačević, S.; Podunavac-Kuzmanović, S.; Gligorić, E.; Gadžurić, S. Analysis of functional ingredients and composition of Ocimum basilicum. S. Afr. J. Bot. 2021, 141, 227–234. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Ashkani, S.; Baghdadi, A.; Pazoki, A.; Jaafar, H.Z.E.; Rahmat, A. Improvement in flavonoids and phenolic acids production and pharmaceutical quality of sweet basil (Ocimum basilicum L.) by ultraviolet-B irradiation. Molecules 2016, 21, 1203. [Google Scholar] [CrossRef] [PubMed]
- Zeljković, S.Ć.; Komzáková, K.; Šišková, J.; Karalija, E.; Smékalová, K.; Tarkowski, P. Phytochemical variability of selected basil genotypes. Ind. Crops Prod. 2020, 157, 112910. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Brede, C. Flavonoids and other phenolics in herbs commonly used in Norwegian commercial kitchens. Food Chem. 2019, 309, 125678. [Google Scholar] [CrossRef] [PubMed]
- Balanescu, F.; Mihaila, M.D.I.; Cârâc, G.; Furdui, B.; Vînătoru, C.; Avramescu, S.M.; Lisa, E.L.; Cudalbeanu, M.; Dinica, R.M. Flavonoid profiles of two new approved romanian Ocimum hybrids. Molecules 2020, 25, 4573. [Google Scholar] [CrossRef] [PubMed]
- Stetsenko, L.A.; Pashkovsky, P.P.; Rvoloshin, A.; Kreslavski, V.D.; Kuznetsov, V.V.; Allakhverdiev, S.I. Role of anthocyanin and carotenoids in the adaptation of the photosynthetic apparatus of purple- and green-leaved cultivars of sweet basil (Ocimum basilicum) to high-intensity light. Photosynthetica 2020, 58, 890–901. [Google Scholar] [CrossRef]
- Da Silva, W.M.F.; Kringel, D.H.; de Souza, E.J.D.; da Rosa Zavareze, E.; Dias, A.R.G. Basil essential oil: Methods of extraction, chemical composition, biological activities, and food applications. Food Bioproc. Tech. 2021, 15, 1–27. [Google Scholar] [CrossRef]
- Avetisyan, A.; Markosian, A.; Petrosyan, M.; Sahakyan, N.; Babayan, A.; Aloyan, S.; Trchounian, A. Chemical composition and some biological activities of the essential oils from basil Ocimum different cultivars. BMC Complement. Altern. Med. 2017, 17, 60. [Google Scholar] [CrossRef]
- Stanojevic, L.P.; Marjanovic-Balaban, Z.R.; Kalaba, V.D.; Stanojevic, J.S.; Cvetkovic, D.J.; Cakic, M.D. Chemical composition, antioxidant and antimicrobial activity of basil (Ocimum basilicum L.) essential oil. J. Essent. Oil Bear. Plants 2017, 20, 1557–1569. [Google Scholar] [CrossRef]
- Ahmed, A.F.; Attia, F.A.K.; Liu, Z.; Li, C.; Wei, J.; Kang, W. Antioxidant activity and total phenolic content of essential oils and extracts of sweet basil (Ocimum basilicum L.) plants. Food Sci. Hum. Wellness 2019, 8, 299–305. [Google Scholar] [CrossRef]
- Jermini, M.; Dubois, J.; Rodondi, P.Y.; Zaman, K.; Buclin, T.; Csajka, C.; Orcurto, A.; Rothuizen, L.E. Complementary medicine use during cancer treatment and potential herb-drug interactions from a cross-sectional study in an academic centre. Sci. Rep. 2019, 9, 5078. [Google Scholar] [CrossRef] [PubMed]
- Majdi, C.; Pereira, C.; Dias, M.I.; Calhelha, R.C.; Alves, M.J.; Frih, B.; Charrouf, Z.; Barros, L.; Amaral, J.S.; Ferreira, I.C.F.R. Phytochemical characterization and bioactive properties of cinnamon basil (Ocimum basilicum cv. ‘Cinnamon’) and Lemon Basil (Ocimum × citriodorum). Antioxidants 2020, 9, 369. [Google Scholar] [CrossRef]
- Mahmoud, G.I. Biological effects, antioxidant and anticancer activities of marigold and basil essential oils. J. Med. Plants Res. 2013, 7, 561–572. [Google Scholar]
- Elansary, H.O.; Mahmoud, E.A. In vitro antioxidant and antiproliferative activities of six international basil cultivars. Nat. Prod. Res. 2015, 29, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Matyjaszczyk, E. Plant protection means used in organic farming throughout the European Union. Pest Manag. Sci. 2017, 74, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Pratyusha, S. Phenolic Compounds in the Plant Development and Defense: AN overview. In Plant Stress Physiology—Perspectives in Agriculture; Nahar, K., Hasanuzzaman, M., Eds.; IntechOpen: London, UK, 2022; Chapter 7; ISSN 2631-8261. [Google Scholar]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant secondary metabolite biosynthesis and transcriptional regulation in response to biotic and abiotic stress conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Khare, S.; Singh, N.B.; Singh, A.; Hussain, I.; Niharika, K.; Yadav, V.; Yadov, R.K.; Amist, N. Plant secondary metabolites synthesis and their regulations under biotic and abiotic constraints. J. Plant Biol. 2020, 63, 203–216. [Google Scholar] [CrossRef]
- Balmer, D.; Flors, V.; Glauser, G.; Mauch-Mani, B. Metabolomics of cereals under biotic stress: Current knowledge and techniques. Front. Plant Sci. 2013, 4, 82. [Google Scholar] [CrossRef]
- Matłok, N.; Stępień, A.E.; Gorzelany, J.; Wojnarowska-Nowak, R.; Balawejder, M. Effects of organic and mineral fertilization on yield and selected quality parameters for dried herbs of two varieties of oregano (Origanum vulgare L.). Appl. Sci. 2020, 16, 5503. [Google Scholar] [CrossRef]
- Stracke, B.A.; Eitel, J.; Watzl, B.; Mäder, P.; Rüfer, C.E. Influence of the production method on phytochemical concentrations in whole wheat (Triticum aestivum L.): A comparative study. J. Agric. Food Chem. 2009, 57, 10116–10121. [Google Scholar] [CrossRef] [PubMed]
- Raigόn, M.D.; Rodríguez-Burruezo, A.; Prohens, J. Effects of organic and conventional cultivation methods on composition of eggplant fruits. J. Agric. Food Chem. 2010, 58, 6833–6840. [Google Scholar] [CrossRef] [PubMed]
- Anton, D.; Matt, D.; Pedastsaar, P.; Bender, I.; Kazimierczak, R.; Roasto, M.; Kaart, T.; Luik, A.; Püssa, T. Three-year comparative study of polyphenol contents and antioxidant capacities in fruits of tomato (Lycopersicon esculentum Mill.) cultivars grown under organic and conventional conditions. J. Agric. Food Chem. 2014, 62, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- Głowacka, A.; Rozpara, E.; Hallmann, E. The dynamic of polyphenols concentrations in organic and conventional sour cherry fruits: Results of a 4-year field study. Molecules 2020, 25, 3729. [Google Scholar] [CrossRef]
- Casanova, J.M.; dos Santos Nascimento, L.B.; Casanova, L.M.; Leal-Costa, M.V.; Costa, S.S.; Tavares, E.S. Differential distribution of flavonoids and phenolic acids in leaves of Kalanchoe delagoensis Ecklon & Zeyher (Crassulaceae). Microsc. Microanal. 2020, 26, 1061–1068. [Google Scholar] [PubMed]
- Ali, M.; Bahadur, S.; Hussain, A.; Saeed, S.; Khuram, I.; Ullah, M.; Shao, J.; Akhtar, N. Foliar epidermal micromorphology and its taxonomic significance in Polygonatum (Asparagaceae) using scanning electron microscopy. Microsc. Res. Tech. 2020, 83, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- European Parliament. Regulation (EU) 2018/848 of the European Parliament and of the Council of 30 May 2018 on Organic Production and Labelling of Organic Products and Repealing Council Regulation (EC) No 834/2007; European Parliament: Strasbourg, France, 2008. [Google Scholar]
- (PN-EN 14346:2011); Fruit and Vegetable Preserves. Sample Preparation and Physicochemical Test Methods. Determination of Dry Matter Content by Gravimetric Method. PSC: Warsaw, Poland, 2011.
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Rusaczonek, A.; Czarnocka, W.; Kacprzak, S.; Witoń, D.; Ślesak, I.; Szechyńska-Hebda, M.; Gawroński, P.; Karpiński, S. Role of phytochromes A and B in the regulation of cell death and acclimatory responses to UV stress in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 6679–6695. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Polish Pharmacopoeia VI; The Polish Pharmaceutical Society: Warsaw, Poland, 2002; pp. 896–898.
- Toiu, A.; Vlase, L.; Vodnar, D.C.; Gheldiu, A.-M.; Oniga, I. Solidago graminifolia L. Salisb. (Asteraceae) as a valuable source of bioactive polyphenols: HPLC profile, in vitro antioxidant and antimicrobial potential. Molecules 2019, 24, 2666. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, E.; Sabała, P. Organic and conventional herbs quality reflected by their antioxidant compounds concentration. Appl. Sci. 2020, 10, 3468. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophyll and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzym. 1987, 148, 331–382. [Google Scholar]
- Chen, C.W.; Ho, C.T. Antioxidant properties of polyphenols extracted from green and black teas. J. Food Lipids 1995, 2, 35–46. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Meth. Enzymol. 1999, 299, 15–27. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Nala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Kamińska, I.; Górecka, M.; Bederska-Błaszczyk, M. Evaluation of heavy metal-induced responses in Silene vulgaris ecotypes. Protoplasma 2019, 256, 1279–1297. [Google Scholar] [CrossRef]
- Hallmann, E.; Rozpara, E.; Słowianek, M.; Leszczyńska, J. The effect of organic and conventional farm management on the allergenic potency and bioactive compounds status of apricots (Prunus armeniaca L.). Food Chem. 2019, 279, 171–178. [Google Scholar] [CrossRef]
- Kazimierczak, R.; Hallmann, E.; Rembiałkowska, E. Effects of organic and conventional production systems on the content of bioactive substances in four species of medicinal plants. Biol. Agric. Hortic. 2014, 31, 118–127. [Google Scholar] [CrossRef]
- Aflatuni, A. The effect of manure composted with drum composter on aromatic plants. Acta Hort. 1993, 344, 63–68. [Google Scholar] [CrossRef]
- Tahami, M.K.; Jahan, M.; Khalilzadeh, H.; Mehdizadeh, M. Plant growth promoting rhizobacteria in an ecological cropping system: A study on basil (Ocimum basilicum L.) essential oil production. Ind. Crops Prod. 2017, 107, 97–104. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M.; Masabni, J.G. Responses of sweet basil to different daily light integrals in photosynthesis, morphology, yield, and nutritional quality. Hort. Sci. 2018, 53, 496–503. [Google Scholar] [CrossRef]
- Masamba, K.G.; Nguyen, M. Determination and comparison of vitamin C, calcium, and potassium in four selected conventionally and organically grown fruits and vegetables. Afr. J. Biotechnol. 2008, 7, 2915–2919. [Google Scholar]
- Evans, N.A.; Bonaventure, K.M.; Saajah, J.K. Effect of soil amendments on the nutritional quality of three commonly cultivated lettuce varieties in Ghana. Afr. J. Agric. Res. 2015, 10, 1796–1804. [Google Scholar] [CrossRef]
- Chiappero, J.; Cappellari, L.d.R.; Palermo, T.B.; Giordano, W.; Khan, N.; Banchio, E. Antioxidant status of medicinal and aromatic plants under the influence of growth-promoting rhizobacteria and osmotic stress. Ind. Crops Prod. 2021, 167, 113541. [Google Scholar] [CrossRef]
- Milić Komić, S.; Živanović, B.; Dumanović, J.; Kolarž, P.; Sedlarević Zorić, A.; Morina, F.; Vidović, M.; Veljović Jovanović, S. Differential antioxidant response to supplemental UV-B irradiation and sunlight in three basil varieties. Int. J. Mol. Sci. 2023, 24, 15350. [Google Scholar] [CrossRef]
- Kwiatkowski, C.A.; Haliniarz, M.; Harasim, E.; Kołodziej, B.; Yakimovich, A. Foliar applied biopreparations as a natural method to increase the productivity of garden thyme (Thymus vulgaris L.) and to improve the quality of herbal raw material. Acta Sci. Pol. Hortorum Cultus 2020, 19, 107–118. [Google Scholar] [CrossRef]
- Ren, F.; Reilly, K.; Gaffney, M.; Kerry, J.P.; Hossain, M.; Rai, D.K. Evaluation of polyphenolic content and antioxidant activity in two onion varieties grown under organic and conventional production systems. J. Sci. Food Agric. 2017, 97, 2982–2990. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.H.; El-Mahdy, M.A.; Abd-Ellah, M.F.; Helal, G.K.; Khalifa, F.; Hamada, F.M. Influence of p-coumaric acid on doxorubicin-induced oxidative stress in rat’s heart. Pharmacol. Res. 2003, 48, 461–465. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. P-coumaric acid kills bacteria through dual damage mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Braglia, R.; Costa, P.; Di Marco, G.; D’Agostino, A.; Redi, E.L.; Scuderi, F.; Gismondi, A.; Canini, A. Phytochemicals and quality level of food plants grown in an aquaponics system. J. Sci. Food Agric. 2021, 102, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Xiong, D.; Chen, J.; Yu, T.; Gao, W.; Ling, X.; Li, Y.; Peng, S.; Huang, J. SPAD-based leaf nitrogen estimation is impacted by environmental factors and crop leaf characteristics. Sci. Rep. 2015, 5, 13389. [Google Scholar] [CrossRef]
- Ievinsh, G.; Andersone-Ozola, U.; Zeipiņa, S. Comparison of the effects of compost and vermicompost soil amendments in organic production of four herb species. Biol. Agric. Hortic. 2020, 4, 267–282. [Google Scholar] [CrossRef]
- Dadan, M.; Tylewicz, U.; Tappi, S.; Rybak, K.; Witrowa-Rajchert, D.; Dalla Rosa, M. Effect of ultrasound, steaming, and dipping on bioactive compound contents and antioxidant capacity of basil and parsley. Pol. J. Food Nutr. Sci. 2021, 71, 311–321. [Google Scholar] [CrossRef]
- Li, L.H.; Lee, J.C.-Y.; Leung, H.H.; Lam, W.C.; Fu, Z.; Lo, A.C.Y. Lutein supplementation for eye diseases, nutrients. Nutrients 2020, 12, 1721. [Google Scholar] [CrossRef]
- Onofrei, V.; Burducea, M.; Lobiuc, A.; Teliban, G.-C.; Ranghiuc, G.; Robu, T. Influence of organic foliar fertilization on antioxidant activity and content of polyphenols in Ocimum basilicum L. Acta Pol. Pharm. 2017, 74, 611–615. [Google Scholar]
- Hęś, M. Comparison of the antioxidant properties of spices from ecological and conventional cultivations. J. Res. Appl. Agric. Eng. 2022, 67, 5–12. [Google Scholar] [CrossRef]
- Vignesh, R.; Venkatesh, N.R.; Meenakshisudaram, B.; Jayapradha, R. Novel instant organic fertilizer and analysis of its growth effects on spinach. J. Biol. Sci. 2012, 3, 105–110. [Google Scholar] [CrossRef]
- Nascimento, L.B.d.S.; Brunetti, C.; Agati, G.; Iacono, C.L.; Detti, C.; Giordani, E.; Ferrini, F.; Gori, A. Short-term pre-harvest UV-B supplement enhances the polyphenol content and antioxidant capacity of Ocimum basilicum leaves during storage. Plants 2020, 9, 797. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Fasseas, C. The dense indumentum with its polyphenol content may replace the protective role of the epidermis in some young xeromorphic leaves. Can. J. Bot. 1996, 74, 347–351. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Compounds Groups/Experimental Combination | 2019 | 2020 | 2021 | p-Value | |||
|---|---|---|---|---|---|---|---|
| ORG | CONV | ORG | CONV | ORG | CONV | C × Y | |
| Polyphenols | 4.34 1 ± 0.93 a2 | 4.16 ± 0.59 a | 4.71 ± 1.25 a | 4.33 ± 2.01 a | 6.66 ± 0.80 a | 6.57 ± 0.83 a | N.S. 3 |
| Phenolic acid | 3.97 ± 0.56 a | 4.09 ± 0.58 a | 4.56 ± 1.11 a | 4.01 ± 2.24 a | 5.70 ± 0.90 a | 5.60 ± 0.80 a | N.S. |
| Flavonoids | 0.44 ± 0.25 a | 0.35 ± 0.06 a | 0.35 ± 0.13 a | 0.35 ± 0.07 a | 1.12 ± 0.10 a | 1.18 ± 0.13 a | N.S. |
| Carotenoids | 0.66 ± 0.16 b | 0.80 ± 0.07 ab | 0.75 ± 0.11 b | 0.91 ± 0.18 a | 0.45 ± 0.10 c | 1.19 ± 0.17 a | <0.0001 |
| Chlorophylls | 2.25 ± 0.53 b | 2.64 ± 0.53 ab | 2.16 ± 0.31 b | 3.15 ± 0.45 a | 1.86 ± 0.73 c | 2.79 ± 1.11 a | <0.0001 |
| Dry matter | 13.34 ± 3.59 a | 11.01 ± 2.32 b | 11.52 ± 2.95 b | 10.95 ± 4.59 c | 12.06 ± 5.15 a | 9.68 ± 4.91 c | 0.0045 |
| Bioactive compounds groups/experimental combination | C (cultivation) | Y (year) | p-value | ||||
| ORG | CONV | 2019 | 2020 | 2021 | C | Y | |
| Polyphenols | 5.24 ± 1.44 A | 4.99 ± 1.70 A | 4.25 ± 0.78 b | 4.52 ± 1.69 b | 6.62 ± 0.82 a | N.S. | <0.0001 |
| Phenolic acids | 4.74 ± 1.14 A | 4.56 ± 1.59 A | 4.03 ± 0.57 b | 4.29 ± 1.79 b | 5.65 ± 0.85 a | N.S. | 0.0005 |
| Flavonoids | 0.64 ± 0.38 A | 0.61 ± 0.40 A | 0.40 ± 0.19 b | 0.35 ± 0.11 b | 1.15 ± 0.12 a | N.S. | <0.0001 |
| Carotenoids | 0.72 ± 0.23 B | 0.85 ± 0.28 A | 0.73 ± 0.14 a | 0.83 ± 0.17 a | 0.82 ± 0.40 a | 0.002 | N.S. |
| Chlorophylls | 2.44 ± 0.76 A | 2.55 ± 0.81 A | 2.44 ± 0.57 a | 2.66 ± 0.63 a | 2.33 ± 1.05 a | N.S. | N.S. |
| Dry matter | 11.97 ± 4.17 A | 10.54 ± 4.15 B | 12.17 ± 3.24 a | 10.73 ± 3.86 b | 10.87 ± 5.17 b | 0.0001 | <0.0001 |
| Activity/Experimental Combination | H2O2 | SOD | CAT | |
|---|---|---|---|---|
| µmol 100 mg−1 | units mg−1 | µmol H2O2 min−1 mg−1 | ||
| 2019 | ORG | 6.53 ± 0.74 b | 59.36 ± 6.00 a | 40.19 ± 3.47 a |
| CONV | 8.88 ± 0.79 a | 79.83 ± 4.72 a | 20.38 ± 5.12 b | |
| 2020 | ORG | 5.84 ± 0.83 bc | 58.21 ± 6.79 a | 40.10 ± 6.44 a |
| CONV | 9.07 ± 0.16 a | 79.27 ± 3.80 a | 26.98 ± 3.26 b | |
| 2021 | ORG | 5.76 ± 0.94 c | 55.26 ± 3.96 a | 37.53 ± 4.12 a |
| CONV | 7.44 ± 0.46 ab | 75.02 ± 3.07 a | 22.23 ± 6.86 b | |
| C (cultivation) | ORG | 6.04 ± 0.91 B | 57.61 ± 5.69 B | 39.28 ± 5.00 A |
| CONV | 8.47 ± 0.90 A | 78.04 ± 4.47 A | 23.19 ± 5.98 B | |
| Y (year) | 2019 | 7.70 ± 1.40 a | 69.60 ± 11.57 a | 30.28 ± 10.83 a |
| 2020 | 7.46 ± 1.72 a | 68.74 ± 11.88 a | 33.54 ± 8.31 a | |
| 2021 | 6.60 ± 1.12 b | 65.14 ± 10.50 b | 29.88 ± 9.52 b | |
| p-value | C | <0.0001 | <0.0001 | <0.0001 |
| Y | <0.0001 | <0.0001 | 0.0004 | |
| C × Y | <0.0001 | N.S. | 0.0035 | |
| Individual Compounds/Experimental Combination | 2019 | 2020 | 2021 | C (Cultivation) | Y (Year) | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ORG | CONV | ORG | CONV | ORG | CONV | ORG | CONV | 2019 | 2020 | 2021 | C | Y | C × Y | |
| Gallic acid | 0.35 1 ± 0.06 a2 | 0.33 ± 0.04 a | 0.34 ± 0.09 a | 0.35 ± 0.16 a | 0.11 ± 0.01 a | 0.12 ± 0.01 a | 0.27 ± 0.13 a | 0.27 ± 0.14 a | 0.34 ± 0.05 a | 0.34 ± 0.13 a | 0.11 ± 0.01 b | N.S. 3 | <0.0001 | N.S. |
| p-Hydroxybenzoic | 0.34 ± 0.07 b | 0.21 ± 0.03 c | 0.26 ± 0.11 c | 0.34 ± 0.19 b | 1.58 ± 0.13 a | 1.41 ± 0.13 a | 0.73 ± 0.62 a | 0.65 ± 0.55 a | 0.27 ± 0.08 b | 0.30 ± 0.16 b | 1.50 ± 0.15 a | N.S. | <0.0001 | 0.018 |
| Caffeic | 0.14 ± 0.05 b | 0.13 ± 0.03 b | 0.19 ± 0.06 ab | 0.15 ± 0.05 b | 0.22 ± 0.08 ab | 0.33 ± 0.13 a | 0.18 ± 0.08 a | 0.20 ± 0.12 a | 0.13 ± 0.04 b | 0.17 ± 0.06 b | 0.28 ± 0.12 a | N.S. | <0.0001 | 0.0141 |
| Rosmarinic | 2.87 ± 0.17 a | 1.88 ± 0.22 a | 2.38 ± 0.32 a | 2.52 ± 0.16 a | 2.28 ± 0.19 a | 2.01 ± 0.14 a | 2.18 ± 0.31 a | 2.12 ± 0.41 a | 2.12 ± 0.23 a | 2.27 ± 0.41 a | 2.21 ± 0.24 a | N.S. | N.S. | N.S. |
| p-Coumaric | 0.06 ± 0.04 c | 0.23 ± 0.04 a | 0.10 ± 0.07 b | 0.15 ± 0.05 b | 0.04 ± 0.00 c | 0.29 ± 0.02 a | 0.07 ± 0.05 b | 0.22 ± 0.08 a | 0.14 ± 0.09 a | 0.13 ± 0.09 a | 0.16 ± 0.12 a | <0.0001 | N.S. | <0.0001 |
| Ferulic | 0.054 ± 0.002 a | 0.063 ± 0.001 a | 0.054 ± 0.002 a | 0.053 ± 0.002 a | 0.025 ± 0.001 a | 0.014 ± 0.001 a | 0.044 ± 0.002 a | 0.043 ± 0.003 a | 0.058 ± 0.001 a | 0.053 ± 0.002 a | 0.020 ± 0.001 b | N.S. | <0.0001 | N.S. |
| Benzoic | 0.14 ± 0.11 a | 0.12 ± 0.03 a | 0.11 ± 0.06 a | 0.13 ± 0.08 a | 0.09 ± 0.01 a | 0.04 ± 0.01 a | 0.11 ± 0.08 a | 0.10 ± 0.06 a | 0.13 ± 0.08 a | 0.12 ± 0.07 a | 0.07 ± 0.03 b | N.S. | 0.025 | N.S. |
| Kaempferol-3-O-glucoside | 0.028 ± 0.001 a | 0.037 ± 0.001 a | 0.027 ± 0.001 a | 0.025 ± 0.001 a | 0.043 ± 0.001 a | 0.044 ± 0.001 a | 0.033 ± 0.001 a | 0.035 ± 0.001 a | 0.032 ± 0.001 b | 0.026 ± 0.001 c | 0.044 ± 0.001 a | N.S. | <0.0001 | N.S. |
| Myricetin | 0.038 ± 0.010 a | 0.058 ± 0.010 a | 0.054 ± 0.030 a | 0.059 ± 0.030 a | 0.034 ± 0.001 a | 0.106 ± 0.050 a | 0.042 ± 0.020 a | 0.075 ± 0.040 a | 0.048 ± 0.010 c | 0.056 ± 0.030 b | 0.070 ± 0.050 a | N.S. | 0.0007 | N.S. |
| Luteolin | 0.017 ± 0.001 b | 0.024 ± 0.001 a | 0.021 ± 0.001 a | 0.024 ± 0.010 a | 0.019 ± 0.001 ab | 0.030 ± 0.001 a | 0.019 ± 0.001 b | 0.026 ± 0.010 a | 0.020 ± 0.010 a | 0.022 ± 0.010 a | 0.024 ± 0.010 a | <0.0001 | N.S. | 0.046 |
| Quercetin | 0.039 ± 0.020 a | 0.042 ± 0.020 a | 0.037 ± 0.020 a | 0.032 ± 0.010 ab | 0.019 ± 0.001 b | 0.045 ± 0.001 a | 0.032 ± 0.020 a | 0.040 ± 0.010 a | 0.041 ± 0.020 a | 0.035 ± 0.020 a | 0.032 ± 0.010 a | 0.036 | N.S. | 0.005 |
| Quercetin-3-O-glucoside | 0.119 ± 0.120 b | 0.118 ± 0.080 b | 0.070 ± 0.060 c | 0.073 ± 0.060 c | 0.781 ± 0.050 a | 0.746 ± 0.080 a | 0.324 ± 0.034 a | 0.312 ± 0.032 a | 0.119 ± 0.010 b | 0.071 ± 0.060 c | 0.764 ± 0.070 a | N.S. | <0.0001 | <0.0001 |
| Kaempferol | 0.029 ± 0.001 a | 0.032 ± 0.010 a | 0.021 ± 0.001 b | 0.022 ± 0.001 b | 0.028 ± 0.001 a | 0.017 ± 0.001 c | 0.026 ± 0.010 a | 0.023 ± 0.010 a | 0.030 ± 0.010 a | 0.021 ± 0.001 b | 0.022 ± 0.010 b | N.S. | 0.0004 | 0.007 |
| Lutein | 0.13 ± 0.02 a | 0.11 ± 0.06 a | 0.17 ± 0.05 a | 0.12 ± 0.07 a | 0.10 ± 0.01 a | 0.07 ± 0.02 a | 0.13 ± 0.04 a | 0.10 ± 0.05 B | 0.12 ± 0.04 b | 0.15 ± 0.06 a | 0.09 ± 0.02 b | 0.022 | 0.0032 | N.S. |
| Zeaxanthin | 0.027 ± 0.001 a | 0.027 ± 0.010 a | 0.027 ± 0.001 a | 0.024 ± 0.010 a | 0.023 ± 0.001 a | 0.020 ± 0.001 a | 0.026 ± 0.001 a | 0.023 ± 0.010 a | 0.027 ± 0.010 a | 0.026 ± 0.001 a | 0.021 ± 0.001 a | N.S. | N.S. | N.S. |
| Beta-carotene | 0.52 ± 0.15 b | 0.40 ± 0.03 c | 0.57 ± 0.16 b | 0.40 ± 0.07 c | 0.20 ± 0.02 d | 0.94 ± 0.02 a | 0.43 ± 0.21 b | 0.58 ± 0.28 a | 0.46 ± 0.12 a | 0.49 ± 0.15 a | 0.57 ± 0.39 a | 0.0004 | N.S. | <0.0001 |
| Chlorophyll a | 2.05 ± 0.46 ab | 1.82 ± 0.47 b | 2.63 ± 0.28 a | 1.78 ± 0.35 b | 1.41 ± 0.46 c | 2.30 ± 0.82 a | 2.03 ± 0.64 a | 1.97 ± 0.63 a | 1.93 ± 0.48 a | 2.21 ± 0.53 a | 1.85 ± 0.80 a | N.S. | N.S. | <0.0001 |
| Chlorophyll b | 0.24 ± 0.07 a | 0.15 ± 0.05 ab | 0.16 ± 0.05 ab | 0.11 ± 0.02 b | 0.06 ± 0.00 c | 0.05 ± 0.01 c | 0.15 ± 0.09 a | 0.10 ± 0.05 b | 0.19 ± 0.08 a | 0.13 ± 0.04 b | 0.06 ± 0.01 c | 0.0002 | <0.0001 | 0.044 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hallmann, E.; Rusaczonek, A.; Muszyńska, E.; Ziółkowski, D.; Kuliński, S.; Jasek, J.; Ponder, A. A Long-Term Study on Chemical Compounds and Their Location in Sweet Basil Leaves from Organic and Conventional Producers. Foods 2024, 13, 383. https://doi.org/10.3390/foods13030383
Hallmann E, Rusaczonek A, Muszyńska E, Ziółkowski D, Kuliński S, Jasek J, Ponder A. A Long-Term Study on Chemical Compounds and Their Location in Sweet Basil Leaves from Organic and Conventional Producers. Foods. 2024; 13(3):383. https://doi.org/10.3390/foods13030383
Chicago/Turabian StyleHallmann, Ewelina, Anna Rusaczonek, Ewa Muszyńska, Daniel Ziółkowski, Sebastian Kuliński, Jakub Jasek, and Alicja Ponder. 2024. "A Long-Term Study on Chemical Compounds and Their Location in Sweet Basil Leaves from Organic and Conventional Producers" Foods 13, no. 3: 383. https://doi.org/10.3390/foods13030383