
High-Resolution Mass Spectrometry-Based Metabolomics for Increased Grape Juice Metabolite Coverage
 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Grape Berry Collection
2.2. Sample Preparation
2.3. DI-FT-ICR-MS
2.4. RP-UHPLC-Q-ToF-MS
2.5. Data Analysis
3. Results and Discussion
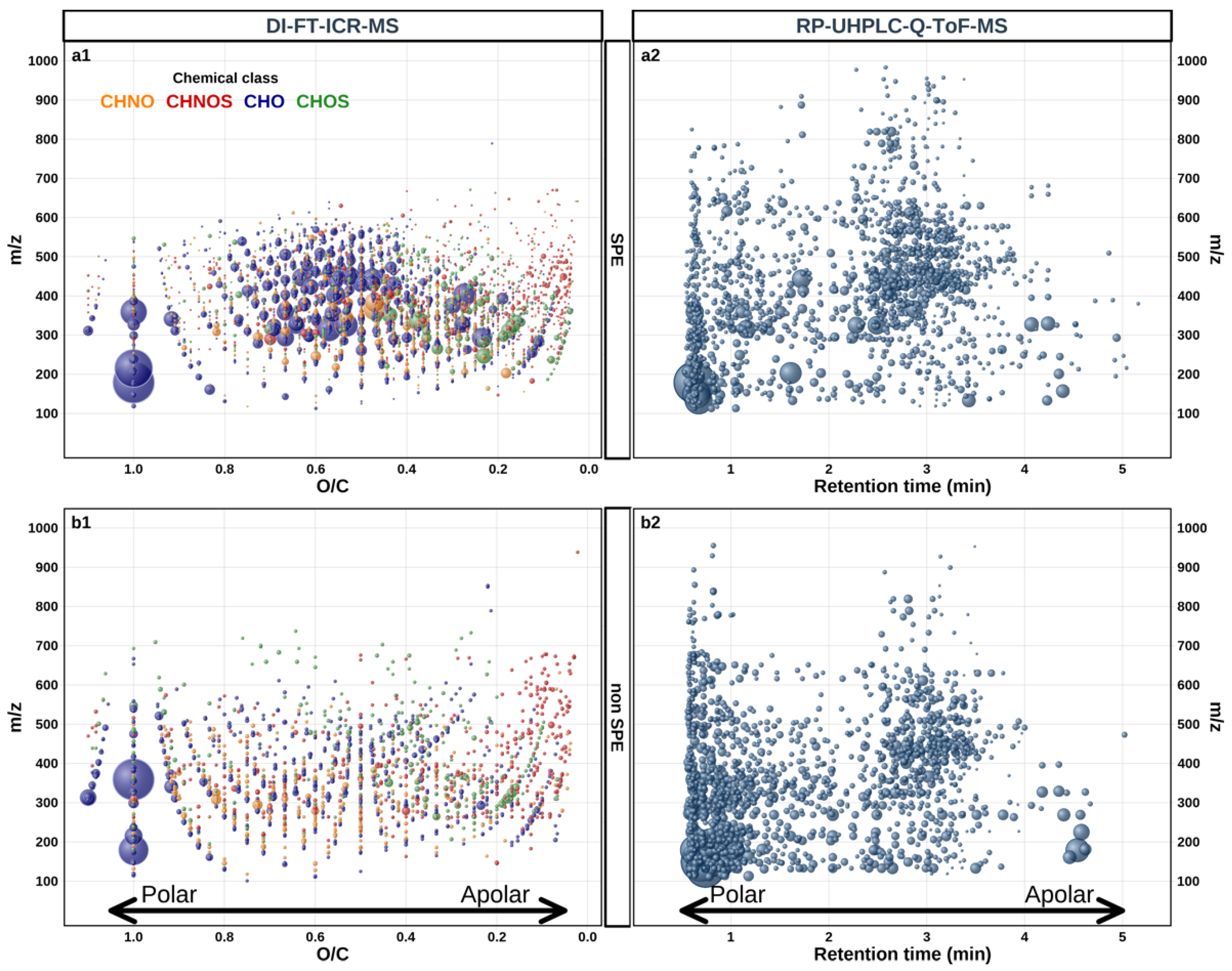
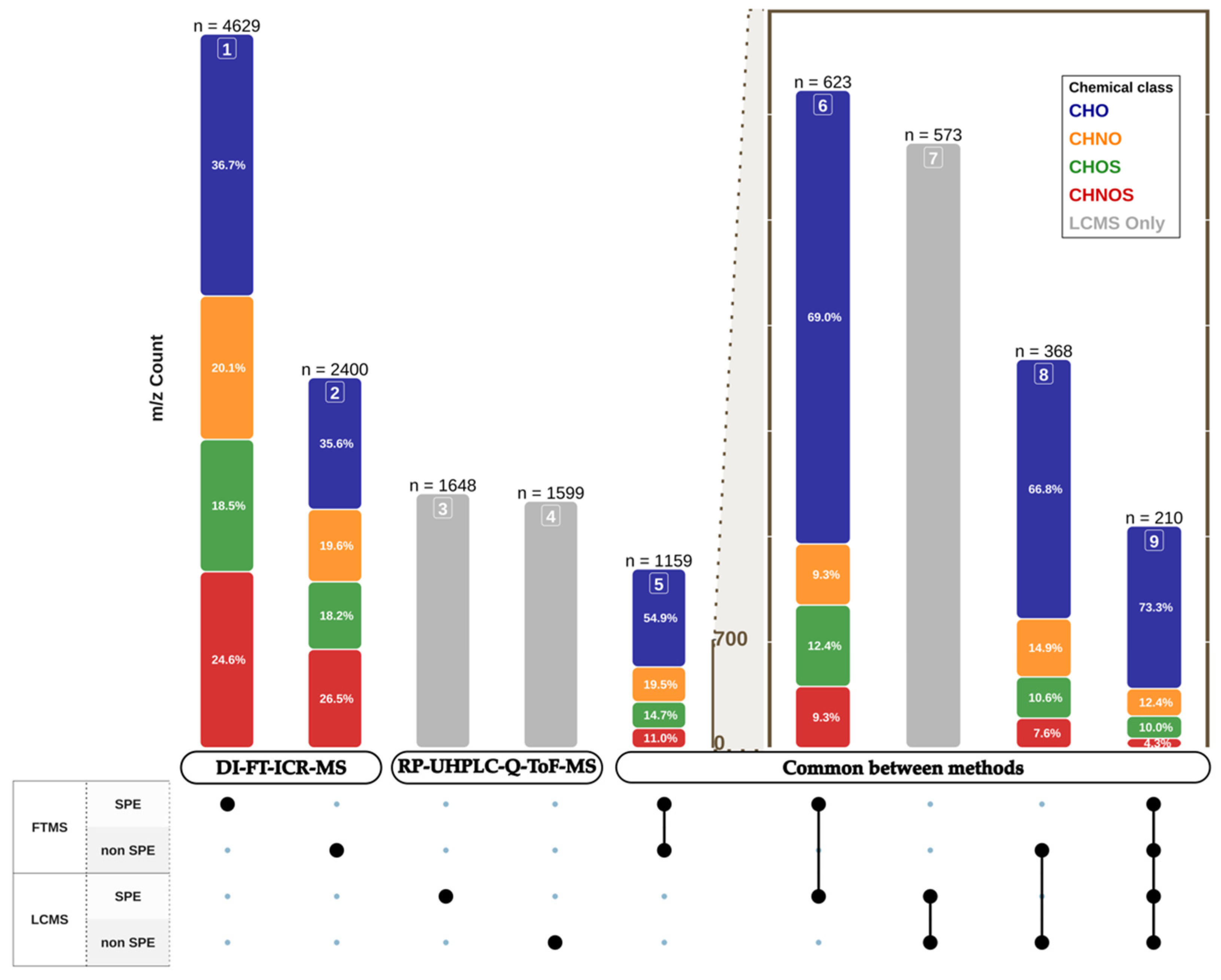
3.1. Global Metabolome of Grape Juices
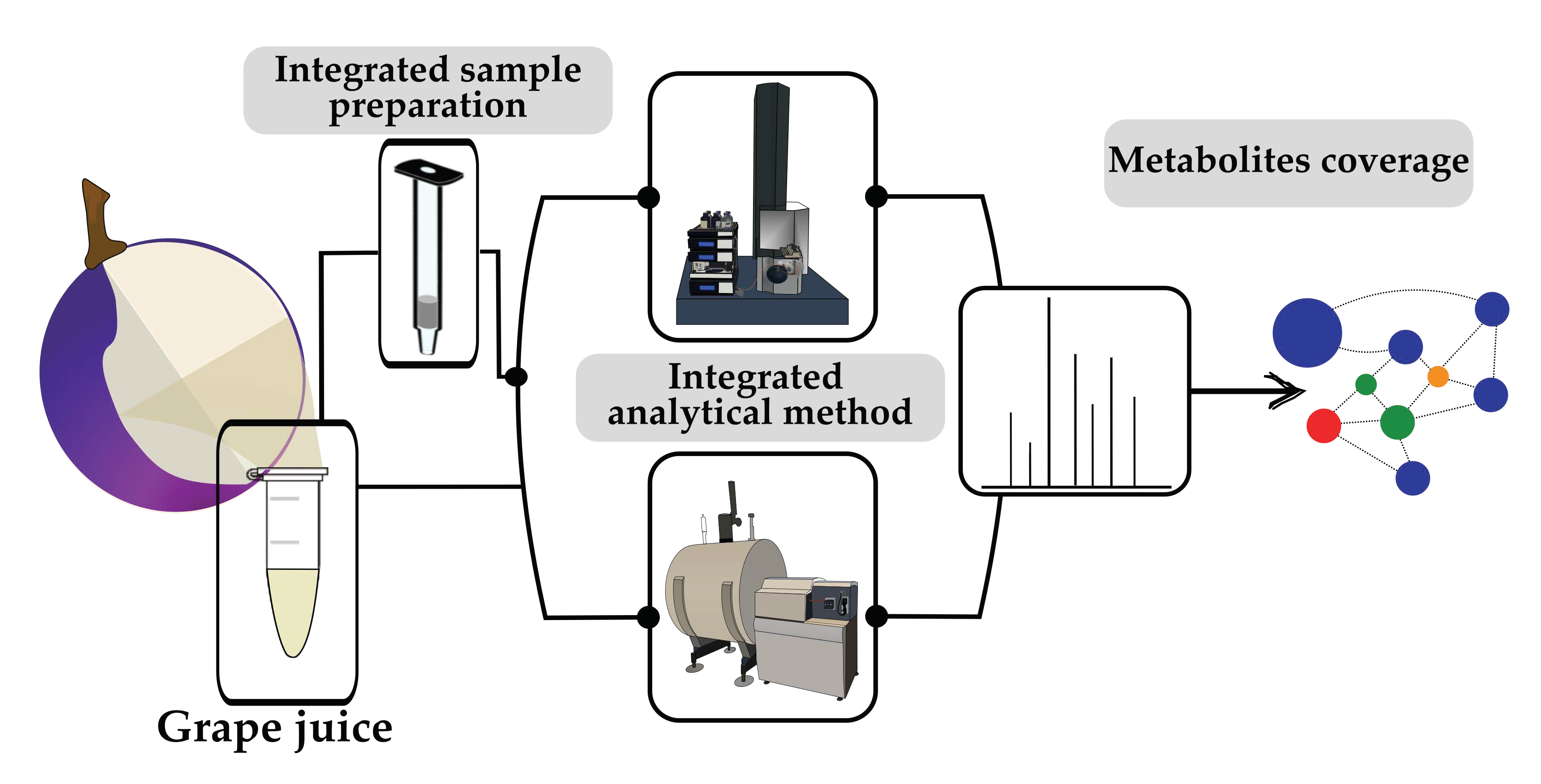
3.2. Aligning the Ultra-High-Resolution Power of DI-FT-ICR-MS with the UHPLC-Controlled High Resolution of RP-UHPLC-Q-ToF-MS
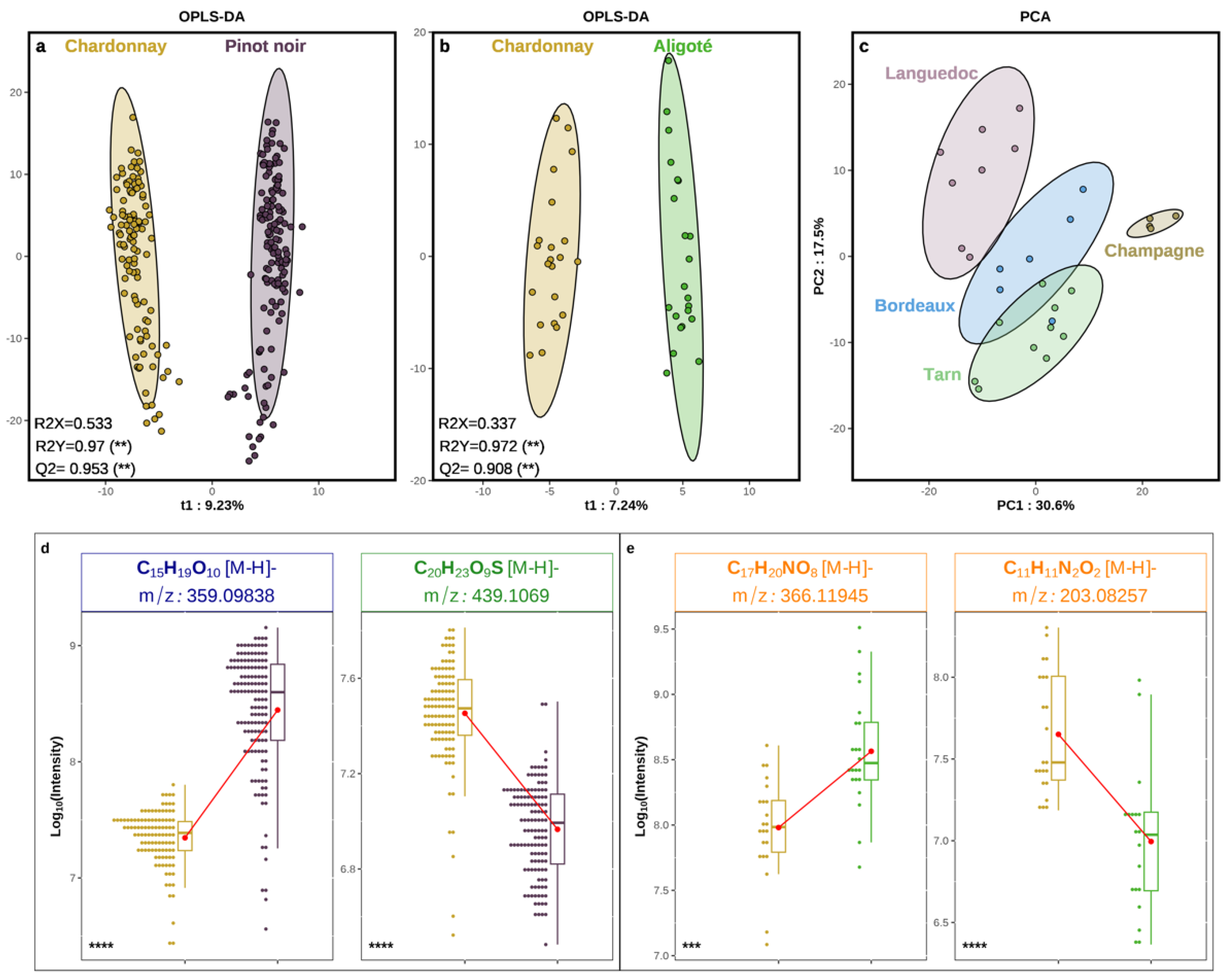
3.3. Grape Juice Discrimination by ESI (−) DI-FT-ICR-MS Applied to SPE Pretreated Samples
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Gerós, H. Biochemical Changes throughout Grape Berry Development and Fruit and Wine Quality. Food 2007, 1, 1–22. [Google Scholar]
- van Leeuwen, C.; Barbe, J.-C.; Darriet, P.; Destrac-Irvine, A.; Gowdy, M.; Lytra, G.; Marchal, A.; Marchand, S.; Plantevin, M.; Poitou, X.; et al. Aromatic Maturity Is a Cornerstone of Terroir Expression in Red Wine: This Article Is Published in Cooperation with Terclim 2022 (XIVth International Terroir Congress and 2nd ClimWine Symposium), 3–8 July 2022, Bordeaux, France. OENO One 2022, 56, 335–351. [Google Scholar] [CrossRef]
- Allamy, L.; van Leeuwen, C.; Pons, A. Impact of Harvest Date on Aroma Compound Composition of Merlot and Cabernet-Sauvignon Must and Wine in a Context of Climate Change: A Focus on Cooked Fruit Molecular Markers. OENO One 2023, 57, 99–112. [Google Scholar] [CrossRef]
- Gobert, A.; Tourdot-Maréchal, R.; Sparrow, C.; Morge, C.; Alexandre, H. Influence of Nitrogen Status in Wine Alcoholic Fermentation. Food Microbiol. 2019, 83, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Sefton, M.A.; Francis, I.L.; Williams, P.J. The Volatile Composition of Chardonnay Juices: A Study by Flavor Precursor Analysis. Am. J. Enol. Vitic. 1993, 44, 359–370. [Google Scholar] [CrossRef]
- Pinu, F.R.; Edwards, P.J.B.; Jouanneau, S.; Kilmartin, P.A.; Gardner, R.C.; Villas-Boas, S.G. Sauvignon Blanc Metabolomics: Grape Juice Metabolites Affecting the Development of Varietal Thiols and Other Aroma Compounds in Wines. Metabolomics 2014, 10, 556–573. [Google Scholar] [CrossRef]
- Hjelmeland, A.K.; Zweigenbaum, J.; Ebeler, S.E. Profiling Monoterpenol Glycoconjugation in Vitis vinifera L. Cv. Muscat of Alexandria Using a Novel Putative Compound Database Approach, High Resolution Mass Spectrometry and Collision Induced Dissociation Fragmentation Analysis. Anal. Chim. Acta 2015, 887, 138–147. [Google Scholar] [CrossRef]
- Cebrián-Tarancón, C.; Oliva, J.; Cámara, M.Á.; Alonso, G.L.; Rosario Salinas, M. Analysis of Intact Glycosidic Aroma Precursors in Grapes by High-Performance Liquid Chromatography with a Diode Array Detector. Foods 2021, 10, 191. [Google Scholar] [CrossRef]
- De Rosso, M.; Lonzarich, V.; Navarini, L.; Flamini, R. Identification of New Glycosidic Terpenols and Norisoprenoids (Aroma Precursors) in C. arabica L. Green Coffee by Using a High-Resolution Mass Spectrometry Database Developed in Grape Metabolomics. Curr. Res. Food Sci. 2022, 5, 336–344. [Google Scholar] [CrossRef]
- Wei, Y.; Chen, Z.; Zhang, X.K.; Duan, C.Q.; Pan, Q.H. Comparative Analysis of Glycosidic Aroma Compound Profiling in Three Vitis vinifera Varieties by Using Ultra-High-Performance Liquid Chromatography Quadrupole-Time-of-Flight Mass Spectrometry. Front. Plant Sci. 2021, 12, 694979. [Google Scholar] [CrossRef]
- Caffrey, A.J.; Lerno, L.A.; Zweigenbaum, J.; Ebeler, S.E. Direct Analysis of Glycosidic Aroma Precursors Containing Multiple Aglycone Classes in Vitis vinifera Berries. J. Agric. Food Chem. 2020, 68, 3817–3833. [Google Scholar] [CrossRef]
- Rienth, M.; Vigneron, N.; Darriet, P.; Sweetman, C.; Burbidge, C.; Bonghi, C.; Walker, R.P.; Famiani, F.; Castellarin, S.D. Grape Berry Secondary Metabolites and Their Modulation by Abiotic Factors in a Climate Change Scenario—A Review. Front. Plant Sci. 2021, 12, 262. [Google Scholar] [CrossRef] [PubMed]
- Evers, M.S.; Roullier-Gall, C.; Morge, C.; Sparrow, C.; Gobert, A.; Alexandre, H. Vitamins in Wine: Which, What for, and How Much? Compr. Rev. Food Sci. Food Saf. 2021, 20, 2991–3035. [Google Scholar] [CrossRef] [PubMed]
- Mattivi, F.; Guzzon, R.; Vrhovsek, U.; Stefanini, M.; Velasco, R. Metabolite Profiling of Grape: Flavonols and Anthocyanins. J. Agric. Food Chem. 2006, 54, 7692–7702. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Gil, A.M.; Gutiérrez-Gamboa, G.; Garde-Cerdán, T.; Pérez-Álvarez, E.P.; Moreno-Simunovic, Y. Characterization of Phenolic Composition in Carignan Noir Grapes (Vitis vinifera L.) from Six Wine-Growing Sites in Maule Valley, Chile. J. Sci. Food Agric. 2018, 98, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Deluc, L.G.; Quilici, D.R.; Decendit, A.; Grimplet, J.; Wheatley, M.D.; Schlauch, K.A.; Mérillon, J.M.; Cushman, J.C.; Cramer, G.R. Water Deficit Alters Differentially Metabolic Pathways Affecting Important Flavor and Quality Traits in Grape Berries of Cabernet Sauvignon and Chardonnay. BMC Genom. 2009, 10, 212. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic Stress Effects on Grapevine (Vitis vinifera L.): Focus on Abscisic Acid-Mediated Consequences on Secondary Metabolism and Berry Quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. “Metabonomics”: Understanding the Metabolic Responses of Living Systems to Pathophysiological Stimuli via Multivariate Statistical Analysis of Biological NMR Spectroscopic Data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef]
- Pinu, F.R. Grape and Wine Metabolomics to Develop New Insights Using Untargeted and Targeted Approaches. Fermentation 2018, 4, 92. [Google Scholar] [CrossRef]
- Toffali, K.; Zamboni, A.; Anesi, A.; Stocchero, M.; Pezzotti, M.; Levi, M.; Guzzo, F. Novel Aspects of Grape Berry Ripening and Post-Harvest Withering Revealed by Untargeted LC-ESI-MS Metabolomics Analysis. Metabolomics 2011, 7, 424–436. [Google Scholar] [CrossRef]
- Savoi, S.; Wong, D.C.J.; Degu, A.; Herrera, J.C.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Multi-Omics and Integrated Network Analyses Reveal New Insights into the Systems Relationships between Metabolites, Structural Genes, and Transcriptional Regulators in Developing Grape Berries (Vitis vinifera L.) Exposed to Water Deficit. Front. Plant Sci. 2017, 8, 1124. [Google Scholar] [CrossRef]
- Savoi, S.; Herrera, J.C.; Carlin, S.; Lotti, C.; Bucchetti, B.; Peterlunger, E.; Castellarin, S.D.; Mattivi, F. From Grape Berries to Wines: Drought Impacts on Key Secondary Metabolites. OENO One 2020, 54, 569–582. [Google Scholar] [CrossRef]
- Moro, L.; da Mota, R.V.; Purgatto, E.; Mattivi, F.; Arapitsas, P. Investigation of Brazilian Grape Juice Metabolomic Profile Changes Caused by Methyl Jasmonate Pre-Harvest Treatment. Int. J. Food Sci. Technol. 2023, 58, 3224–3233. [Google Scholar] [CrossRef]
- Narduzzi, L.; Stanstrup, J.; Mattivi, F. Comparing Wild American Grapes with Vitis vinifera: A Metabolomics Study of Grape Composition. J. Agric. Food Chem. 2015, 63, 6823–6834. [Google Scholar] [CrossRef]
- Theodoridis, G.; Gika, H.; Franceschi, P.; Caputi, L.; Arapitsas, P.; Scholz, M.; Masuero, D.; Wehrens, R.; Vrhovsek, U.; Mattivi, F. LC-MS Based Global Metabolite Profiling of Grapes: Solvent Extraction Protocol Optimisation. Metabolomics 2011, 8, 175–185. [Google Scholar] [CrossRef]
- Nelson, A.R.; Toyoda, J.; Chu, R.K.; Tolić, N.; Garayburu-Caruso, V.A.; Saup, C.M.; Renteria, L.; Wells, J.R.; Stegen, J.C.; Wilkins, M.J.; et al. Implications of Sample Treatment on Characterization of Riverine Dissolved Organic Matter. Environ. Sci. Process Impacts 2022, 24, 773–782. [Google Scholar] [CrossRef]
- Schmitt-Kopplin, P.; Hemmler, D.; Moritz, F.; Gougeon, R.D.; Lucio, M.; Meringer, M.; Müller, C.; Harir, M.; Hertkorn, N. Systems Chemical Analytics: Introduction to the Challenges of Chemical Complexity Analysis. Faraday Discuss. 2019, 218, 9–28. [Google Scholar] [CrossRef]
- Cooper, H.J.; Marshall, A.G. Electrospray Ionization Fourier Transform Mass Spectrometric Analysis of Wine. J. Agric. Food Chem. 2001, 49, 5710–5718. [Google Scholar] [CrossRef] [PubMed]
- Catharine, R.R.; Cunha, I.B.S.; Fogaça, A.O.; Facco, E.M.P.; Godoy, H.T.; Daudt, C.E.; Eberlin, M.N.; Sawaya, A.C.H.F. Characterization of Must and Wine of Six Varieties of Grapes by Direct Infusion Electrospray Ionization Mass Spectrometry. J. Mass. Spectrom. 2006, 41, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, R.D.; Lucio, M.; Frommberger, M.; Peyron, D.; Chassagne, D.; Alexandre, H.; Feuillat, F.; Voilley, A.; Cayot, P.; Gebefügi, I.; et al. The Chemodiversity of Wines Can Reveal a Metabologeography Expression of Cooperage Oak Wood. Proc. Natl. Acad. Sci. USA 2009, 106, 9174–9179. [Google Scholar] [CrossRef] [PubMed]
- Dou, Y.; Mäkinen, M.; Jänis, J. Analysis of Volatile and Nonvolatile Constituents in Gin by Direct-Infusion Ultrahigh-Resolution ESI/APPI FT-ICR Mass Spectrometry. J. Agric. Food Chem. 2023, 71, 7082–7089. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, B.G.; Tosato, F.; Folli, G.S.; de Leite, J.A.; Ventura, J.A.; Endringer, D.C.; Filgueiras, P.R.; Romão, W. Controlling the Quality of Grape Juice Adulterated by Apple Juice Using ESI(-)FT-ICR Mass Spectrometry. Microchem. J. 2019, 149, 104033. [Google Scholar] [CrossRef]
- Pieczonka, S.A.; Lucio, M.; Rychlik, M.; Schmitt-Kopplin, P. Decomposing the Molecular Complexity of Brewing. NPJ Sci. Food 2020, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Roullier-Gall, C.; Witting, M.; Tziotis, D.; Ruf, A.; Gougeon, R.D.; Schmitt-Kopplin, P. Integrating Analytical Resolutions in Non-Targeted Wine Metabolomics. Tetrahedron 2015, 71, 2983–2990. [Google Scholar] [CrossRef]
- Tziotis, D.; Hertkorn, N.; Schmitt-Kopplin, P. Kendrick-Analogous Network Visualisation of Ion Cyclotron Resonance Fourier Transform Mass Spectra: Improved Options for the Assignment of Elemental Compositions and the Classification of Organic Molecular Complexity. Eur. J. Mass. Spectrom. 2011, 17, 415–421. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org (accessed on 6 November 2023).
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Bararpour, N.; Gilardi, F.; Carmeli, C.; Sidibe, J.; Ivanisevic, J.; Caputo, T.; Augsburger, M.; Grabherr, S.; Desvergne, B.; Guex, N.; et al. DBnorm as an R Package for the Comparison and Selection of Appropriate Statistical Methods for Batch Effect Correction in Metabolomic Studies. Sci. Rep. 2021, 11, 5657. [Google Scholar] [CrossRef]
- Giordan, M. A Two-Stage Procedure for the Removal of Batch Effects in Microarray Studies. Stat. Biosci. 2014, 6, 73–84. [Google Scholar] [CrossRef]
- Walach, J.; Filzmoser, P.; Hron, K. Chapter Seven—Data Normalization and Scaling: Consequences for the Analysis in Omics Sciences. In Data Analysis for Omic Sciences: Methods and Applications; Jaumot, J., Bedia, C., Tauler, R.B.T.-C.A.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 82, pp. 165–196. ISBN 0166-526X. [Google Scholar]
- Uthe, H.; van Dam, N.M.; Hervé, M.R.; Sorokina, M.; Peters, K.; Weinhold, A. A Practical Guide to Implementing Metabolomics in Plant Ecology and Biodiversity Research. Adv. Bot. Res. 2021, 98, 163–203. [Google Scholar] [CrossRef]
- ter Braak, C.J.F. Redundancy Analysis Includes Analysis of Variance-Simultaneous Component Analysis (ASCA) and Outperforms Its Extensions. Chemom. Intell. Lab. Syst. 2023, 240, 104898. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 6 November 2023).
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R Package version 0.7.2. 2023. Available online: https://cran.r-project.org/package=rstatix (accessed on 6 November 2023).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rainer, J.; Vicini, A.; Salzer, L.; Stanstrup, J.; Badia, J.M.; Neumann, S.; Stravs, M.A.; Verri Hernandes, V.; Gatto, L.; Gibb, S.; et al. A Modular and Expandable Ecosystem for Metabolomics Data Annotation in R. Metabolites 2022, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Roullier-Gall, C.; Witting, M.; Gougeon, R.D.; Schmitt-Kopplin, P. High Precision Mass Measurements for Wine Metabolomics. Front. Chem. 2014, 2, 102. [Google Scholar] [CrossRef] [PubMed]
- Roullier-Gall, C.; Boutegrabet, L.; Gougeon, R.D.; Schmitt-Kopplin, P. A Grape and Wine Chemodiversity Comparison of Different Appellations in Burgundy: Vintage vs. Terroir Effects. Food Chem. 2014, 152, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Rodgers, R.P.; Marshall, A.G. Two- and Three-Dimensional van Krevelen Diagrams: A Graphical Analysis Complementary to the Kendrick Mass Plot for Sorting Elemental Compositions of Complex Organic Mixtures Based on Ultrahigh-Resolution Broadband Fourier Transform Ion Cyclotron Resonance. Anal. Chem. 2004, 76, 2511–2516. [Google Scholar] [CrossRef]
- Fougère, L.; Zubrzycki, S.; Elfakir, C.; Destandau, E. Characterization of Corn Silk Extract Using HPLC/HRMS/MS Analyses and Bioinformatic Data Processing. Plants 2023, 12, 721. [Google Scholar] [CrossRef]
- Perdue, E.M.; Green, N.W. Isobaric Molecular Formulae of C, H, and O: A View from the Negative Quadrants of van Krevelen Space. Anal. Chem. 2015, 87, 5079–5085. [Google Scholar] [CrossRef]
- Palacio Lozano, D.C.; Jones, H.E.; Ramirez Reina, T.; Volpe, R.; Barrow, M.P. Unlocking the Potential of Biofuels: Via Reaction Pathways in van Krevelen Diagrams. Green. Chem. 2021, 23, 8949–8963. [Google Scholar] [CrossRef]
- Forcisi, S.; Moritz, F.; Lucio, M.; Lehmann, R.; Stefan, N.; Schmitt-Kopplin, P. Solutions for Low and High Accuracy Mass Spectrometric Data Matching: A Data-Driven Annotation Strategy in Nontargeted Metabolomics. Anal. Chem. 2015, 87, 8917–8924. [Google Scholar] [CrossRef]
- King, R.; Bonfiglio, R.; Fernandez-Metzler, C.; Miller-Stein, C.; Olah, T. Mechanistic Investigation of Ionization Suppression in Electrospray Ionization. J. Am. Soc. Mass. Spectrom. 2000, 11, 942–950. [Google Scholar] [CrossRef]
- Cai, Y.; Concha, M.C.; Murray, J.S.; Cole, R.B. Evaluation of the Role of Multiple Hydrogen Adducts in Electrospray Mass Spectrometry. J. Am. Soc. Mass. Spectrom. 2002, 13, 1360–1369. [Google Scholar] [CrossRef]
- Gardiman, M.; De Rosso, M.; De Marchi, F.; Flamini, R. Metabolomic Profiling of Different Clones of Vitis vinifera L. Cv.“Glera” and “Glera Lunga” Grapes by High-Resolution Mass Spectrometry. Metabolomics 2023, 19, 25. [Google Scholar] [CrossRef]
- Meng, N.; Ren, Z.-Y.; Yang, X.-F.; Pan, Q.-H. Effects of Simple Rain-Shelter Cultivation on Fatty Acid and Amino Acid Accumulation in ‘Chardonnay’ Grape Berries. J. Sci. Food Agric. 2018, 98, 1222–1231. [Google Scholar] [CrossRef]
- Fernández-Cruz, E.; Cerezo, A.B.; Cantos-Villar, E.; Troncoso, A.M.; García-Parrilla, M.C. Time Course of L-Tryptophan Metabolites When Fermenting Natural Grape Musts: Effect of Inoculation Treatments and Cultivar on the Occurrence of Melatonin and Related Indolic Compounds. Aust. J. Grape Wine Res. 2019, 25, 92–100. [Google Scholar] [CrossRef]
- Gawel, R.; Day, M.; Van Sluyter, S.C.; Holt, H.; Waters, E.J.; Smith, P.A. White Wine Taste and Mouthfeel as Affected by Juice Extraction and Processing. J. Agric. Food Chem. 2014, 62, 10008–10014. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Aligoté | Chardonnay | Meunier | Pinot Noir | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | 2021 | 2019 | 2020 | 2021 | 2019 | 2020 | 2021 | 2019 | 2020 | 2021 | |
| Adige valley | 10 | |||||||||||
| Alsace | 2 | 4 | 4 | |||||||||
| Beaujolais | 1 | |||||||||||
| Bordeaux | 1 | 2 | 1 | 2 | 2 | 1 | 2 | 1 | 2 | 2 | ||
| Burgundy | 4 | 4 | 2 | 20 | 35 | 8 | 29 | 26 | 10 | |||
| Champagne | 2 | 1 | 1 | 1 | 2 | 1 | ||||||
| Douro | 4 | |||||||||||
| Gaillac | 1 | 2 | 2 | 1 | 2 | 2 | 1 | 2 | 2 | 1 | 2 | 2 |
| Languedoc | 2 | 2 | 6 | 6 | 2 | 2 | 6 | 6 | ||||
| Piedmont | 2 | 2 | 2 | 3 | ||||||||
| Rheingau | 14 | |||||||||||
| Uco Valley | 5 | 6 | 8 | 6 | ||||||||
| Württemberg | 2 | 2 | ||||||||||
| Method | DI-FT-ICR-MS | RP-UHPLC-Q-ToF-MS | |||||
|---|---|---|---|---|---|---|---|
| n (Total) | Chemical Class | n Chemical Class | n Total 1 | n Chemical Class 1 | n Isobars | ||
| Mean 2 | Range 2 | ||||||
| SPE | 623 (11.4%) | CHO | 430 (69%) | 1035 (62.8%) | 794 (76.7%) | 1.5 | 1–6 |
| CHNO | 58 (9.3%) | 71 (6.9%) | 1.1 | 1–3 | |||
| CHNOS | 58 (9.3%) | 77 (7.4%) | 1.2 | 1–6 | |||
| CHOS | 77 (12.4%) | 93 (9%) | 1.1 | 1–4 | |||
| non SPE | 368 (12.2%) | CHO | 246 (66.8%) | 825 (51.6%) | 623 (75.5%) | 1.8 | 1–13 |
| CHNO | 55 (14.9%) | 99 (12%) | 1.4 | 1–4 | |||
| CHNOS | 28 (7.6%) | 47 (5.7%) | 1.3 | 1–4 | |||
| CHOS | 39 (10.6%) | 56 (6.8%) | 1.2 | 1–3 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicolas, S.; Bois, B.; Billet, K.; Romanet, R.; Bahut, F.; Uhl, J.; Schmitt-Kopplin, P.; Gougeon, R.D. High-Resolution Mass Spectrometry-Based Metabolomics for Increased Grape Juice Metabolite Coverage. Foods 2024, 13, 54. https://doi.org/10.3390/foods13010054
Nicolas S, Bois B, Billet K, Romanet R, Bahut F, Uhl J, Schmitt-Kopplin P, Gougeon RD. High-Resolution Mass Spectrometry-Based Metabolomics for Increased Grape Juice Metabolite Coverage. Foods. 2024; 13(1):54. https://doi.org/10.3390/foods13010054
Chicago/Turabian StyleNicolas, Sébastien, Benjamin Bois, Kevin Billet, Rémy Romanet, Florian Bahut, Jenny Uhl, Philippe Schmitt-Kopplin, and Régis D. Gougeon. 2024. "High-Resolution Mass Spectrometry-Based Metabolomics for Increased Grape Juice Metabolite Coverage" Foods 13, no. 1: 54. https://doi.org/10.3390/foods13010054