
Effect of Air Drying on the Metabolic Profile of Fresh Wild and Artificial Cordyceps sinensis

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Pretreatment
2.3. UPLC-MS/MS Analysis
2.4. Data Processing and Metabolite Identification
3. Results
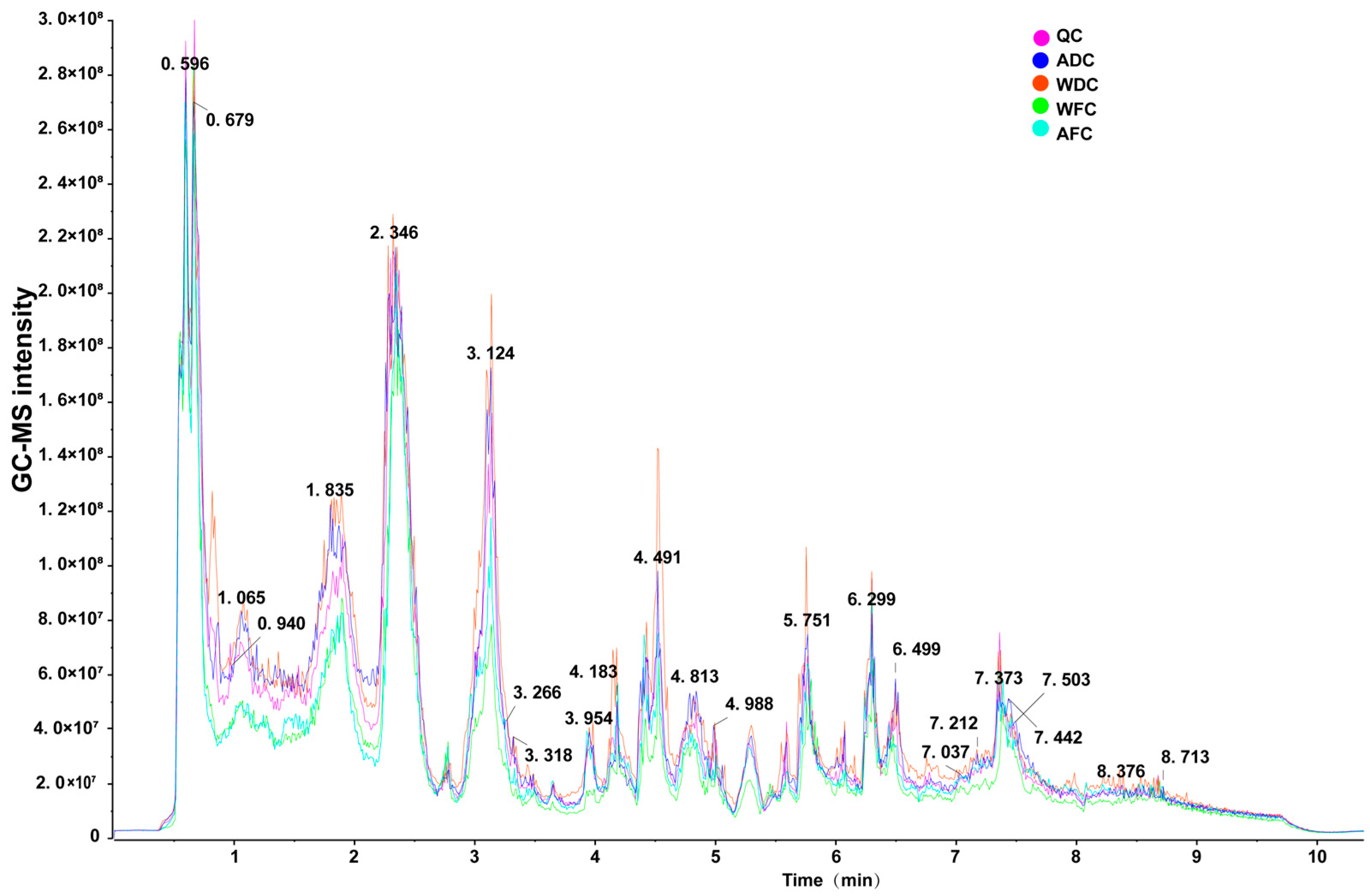
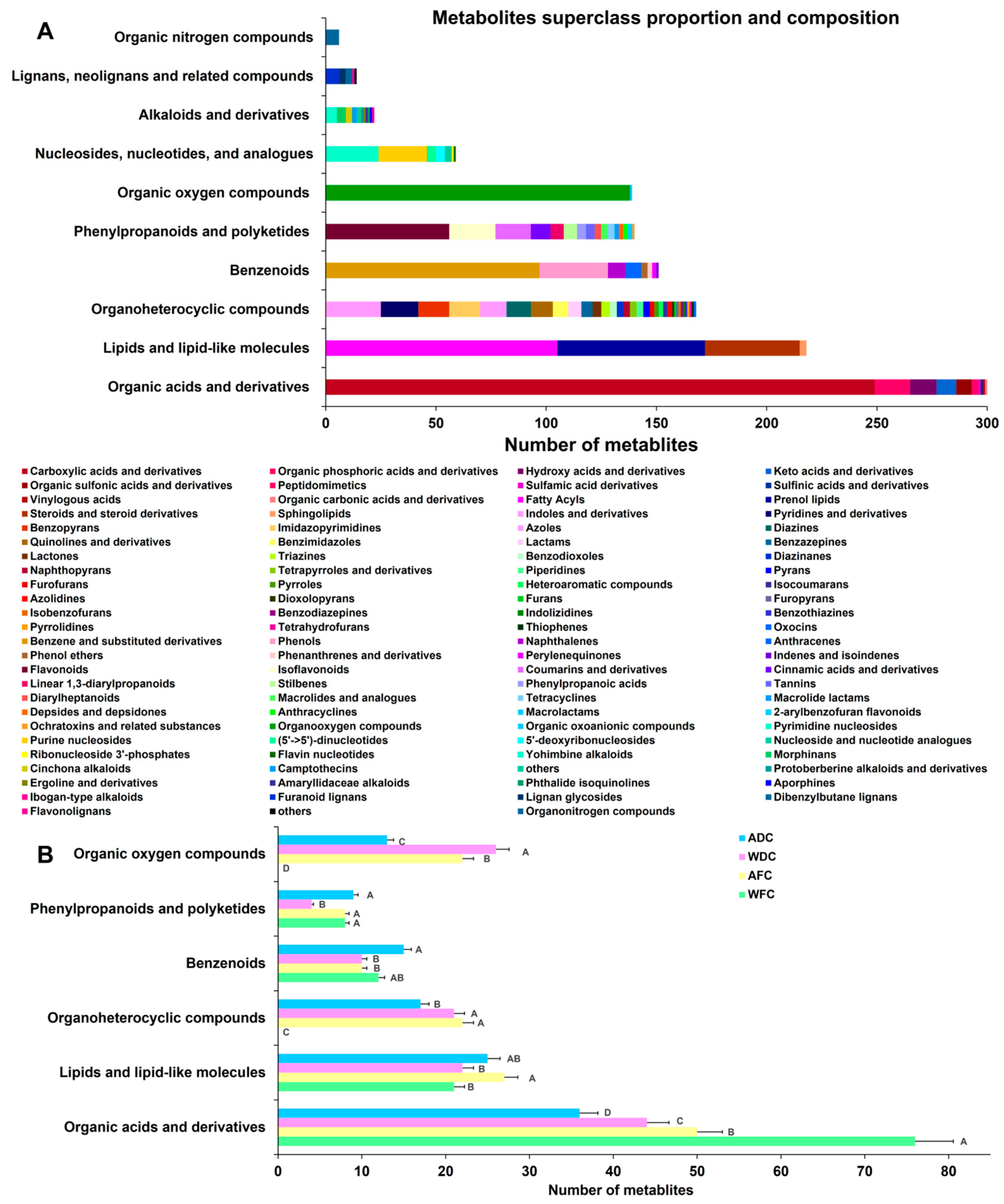
3.1. Overview of Metabolite Profiling
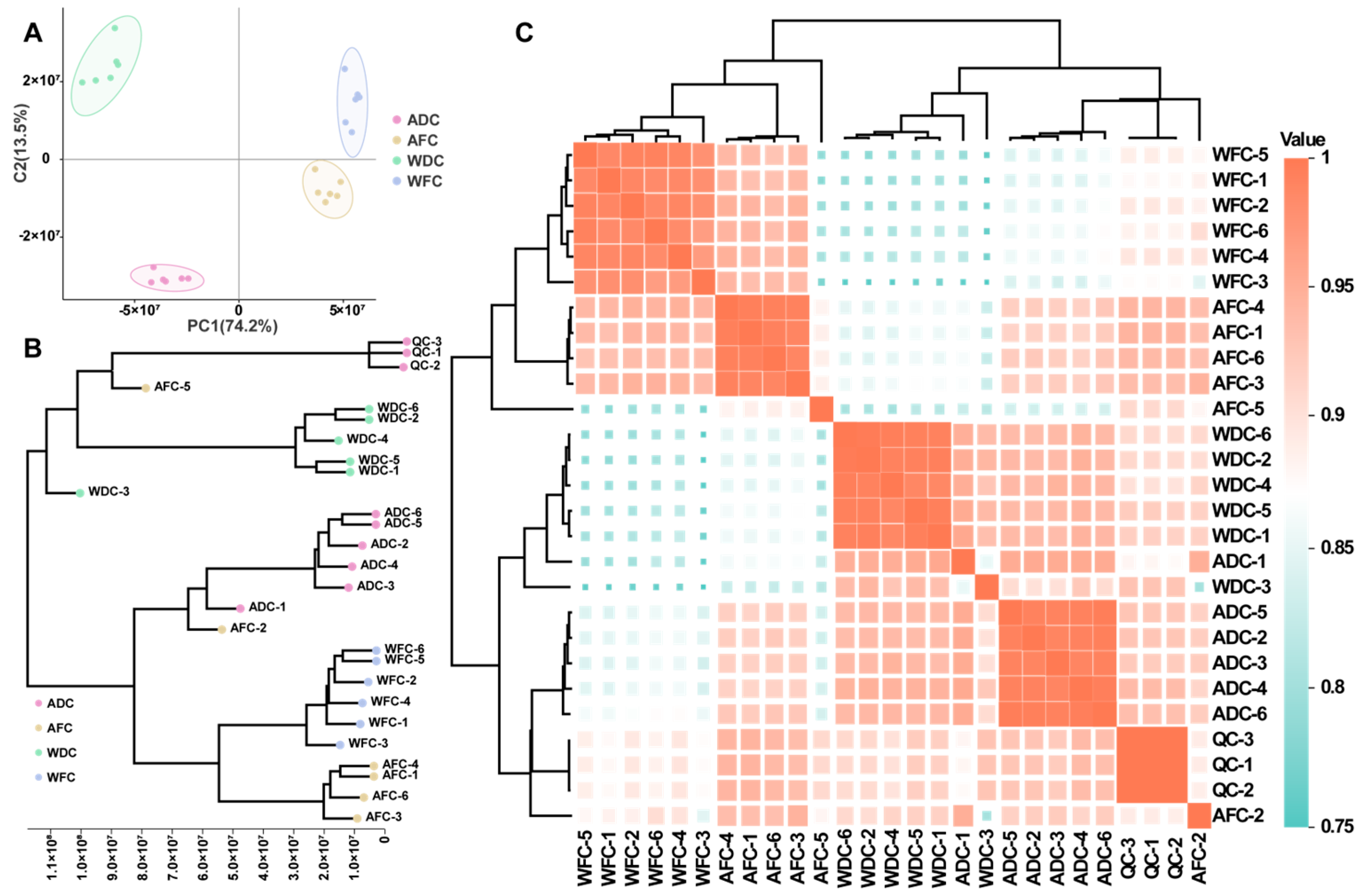
3.2. Multivariate Statistical Analysis
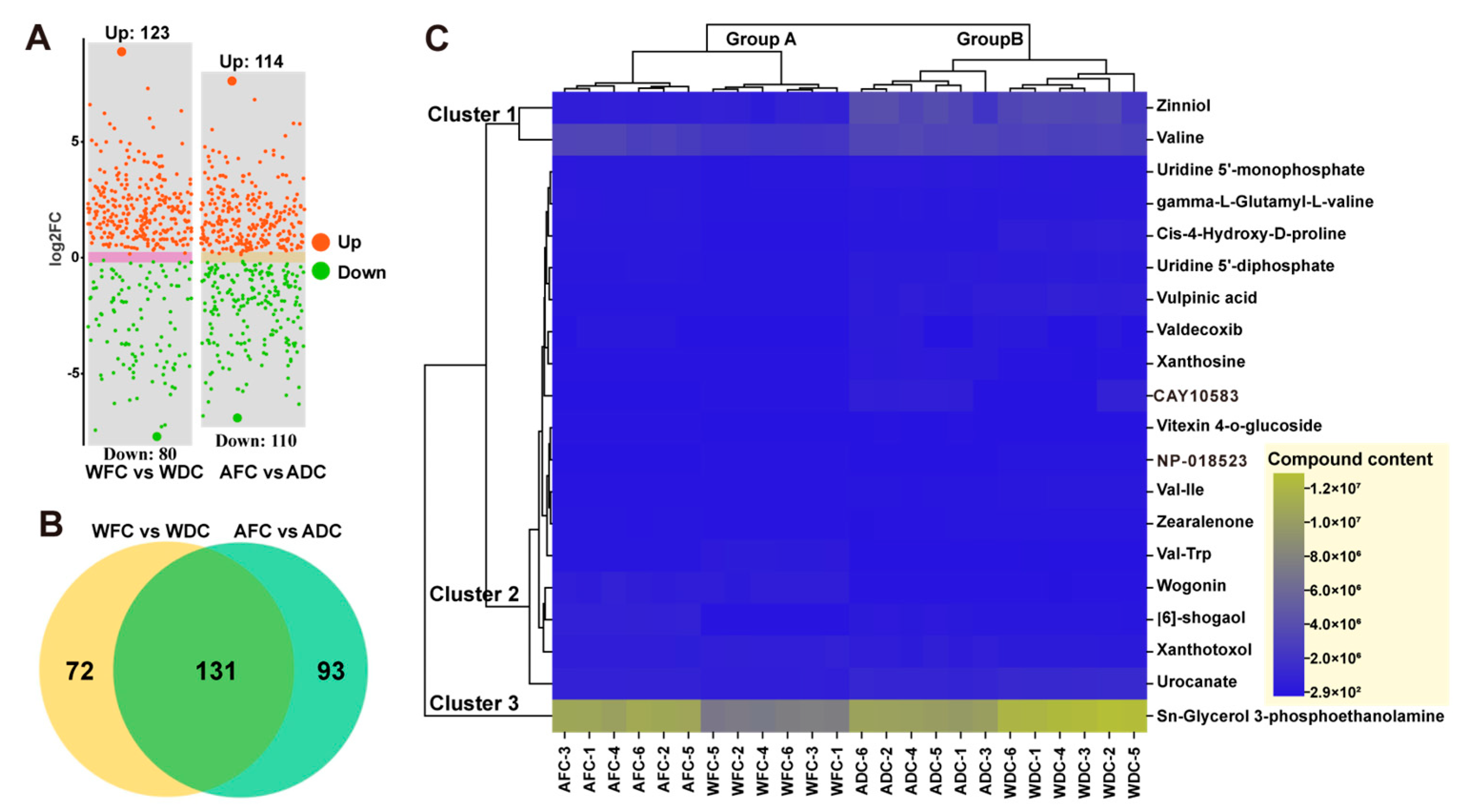
3.3. Screening of DAMs and Hierarchical Cluster Analysis (HCA)
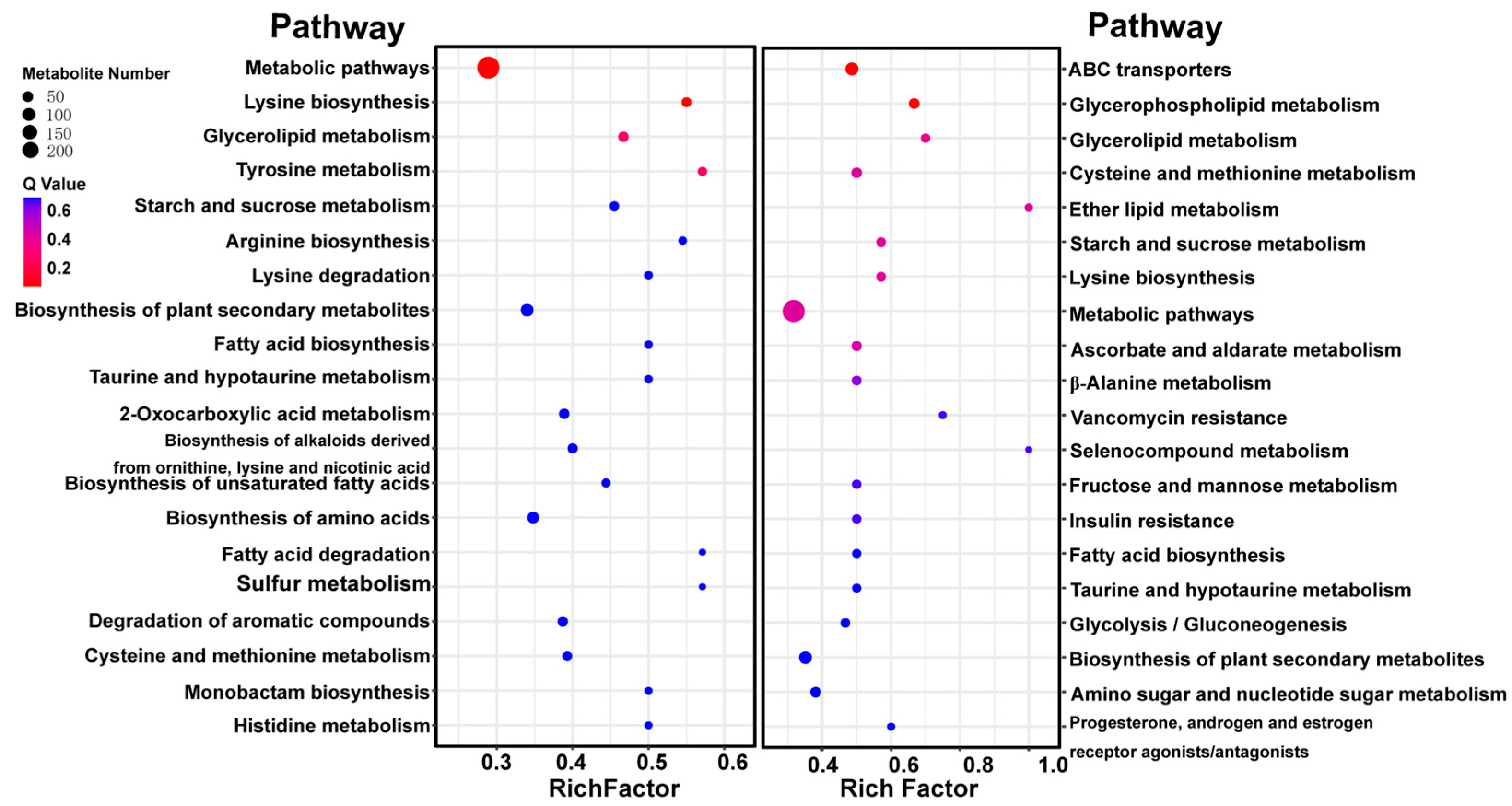
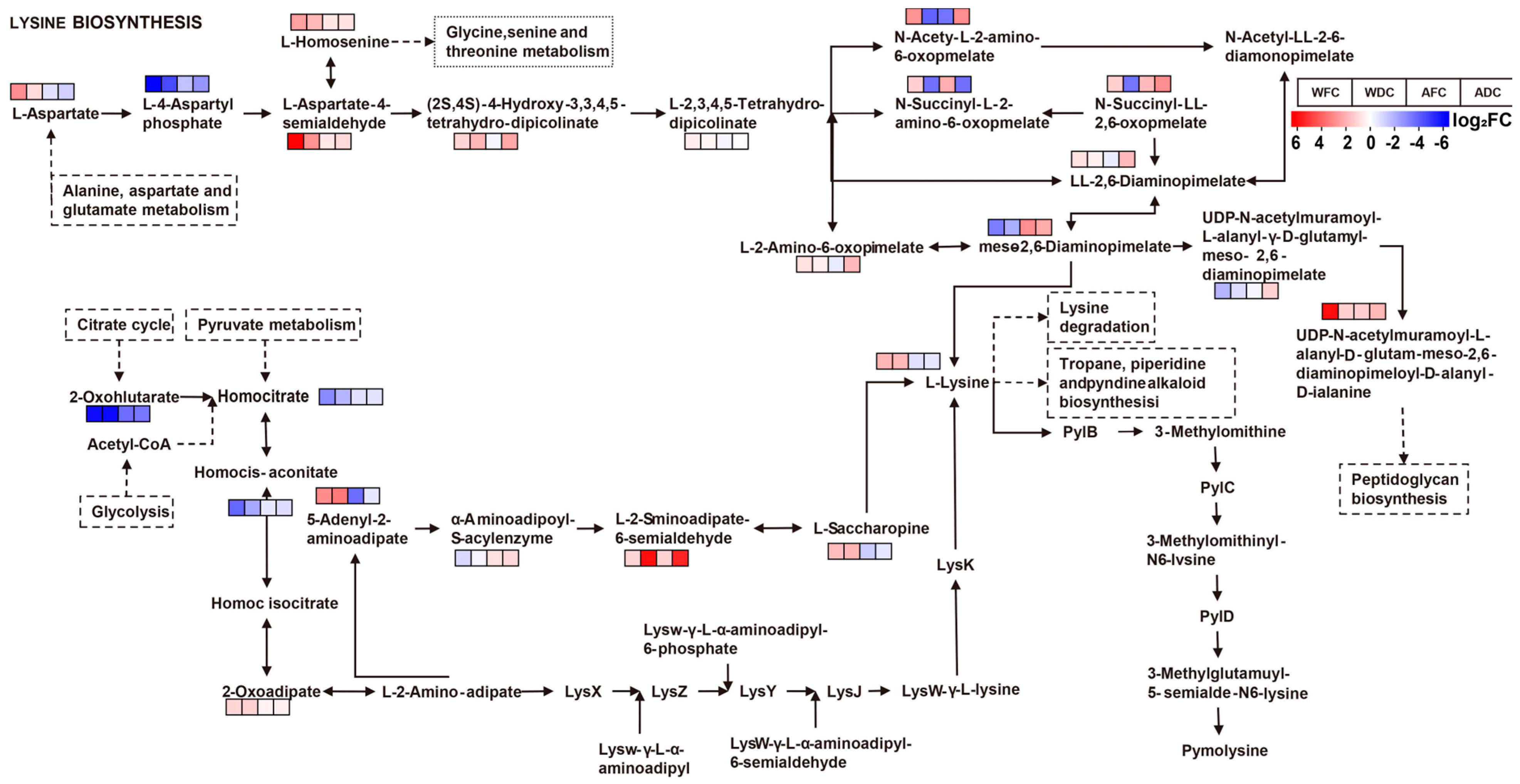
3.4. KEGG Annotation and Metabolite Enrichment Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hou, Y.; Jiang, J.G. Origin and concept of medicine food homology and its application in modern functional foods. Food Funct. 2013, 12, 1727–1741. [Google Scholar] [CrossRef] [PubMed]
- Shalik, R.S.; Maan, B.R.; Zuzana, M.; Eryuan, L. Habitat Ecology of Ophiocordyceps sinensis in Western Nepal. Mt. Res. Dev. 2017, 37, 216–223. [Google Scholar]
- Liu, L.; Guan, L.L.; Zhao, H.X.; Huang, Y.; Mou, Q.Y.; Liu, K.; Chen, T.Y.; Wang, X.Y.; Zhang, Y.; Wei, B.; et al. Modeling habitat suitability of Houttuynia cordata Thunb (Ceercao) using MaxEnt under climate change in China. Ecol. Inform. 2021, 63, 101324. [Google Scholar] [CrossRef]
- Zheng, Y.L.; Mei, Q.X.; Li, W.J.; Tang, Z.F.; Qian, Z.M.; Tao, S.C.; Dong, P.P. Discussion on the medicinal history and modern administration method of Cordyceps sinensis. J. Chin. Med. Mater. 2017, 40, 2722–2725. [Google Scholar]
- Li, G.R.; Guo, Y.F.; Yu, J.F.; Liang, G.H.; Fang, Q.M.; Li, W.J. Effects of different drying processes on quality of Chinese Cordyceps. J. Fungal Res. 2020, 18, 132–138. [Google Scholar]
- Wang, S.Y.; Shiao, M.S. Pharmacological functions of Chinese medicinal fungus Cordyceps sinensis and related species. J. Food Drug Anal. 2020, 8, e15. [Google Scholar] [CrossRef]
- Xu, J.; Huang, Y.; Chen, X.X.; Zheng, S.C.; Chen, P.; Mo, M.H. The mechanisms of pharmacological activities of Ophiocordyceps sinensis fungi. Phytother. Res. 2016, 30, 1572–1583. [Google Scholar] [CrossRef]
- Opeyemi, J.O.; Tang, J.; Tola, A.; Auberon, F.; Omolara, O.; Zhen, O.Y. The genus Cordyceps: An extensive review of its traditional uses, phytochemistry and pharmacology. Fitoterapia 2018, 129, 293–316. [Google Scholar]
- Ng, T.B.; Wang, H.X. Pharmacological actions of Cordyceps, a prized folk medicine. J. Pharm. Pharmacol. 2005, 57, 1509–1519. [Google Scholar] [CrossRef]
- Li, S.P.; Su, Z.R.; Dong, T.X.; Tsim, K.W.K. The fruiting body and its caterpillar host of Cordyceps sinensis show close resemblance in main constituents and anti-oxidation activity. Phytomedicine 2002, 9, 319–324. [Google Scholar] [CrossRef]
- Wang, J.Q.; Nie, S.P.; Kan, L.J.; Chen, H.H.; Cui, S.W.; Phillips, A.O.; Phillips, G.O.; Xie, M.Y. Comparison of structural features and antioxidant activity of polysaccharides from natural and cultured Cordyceps sinensis. Food Sci. Biotechnol. 2017, 26, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.L.; Yu, X.F.; Ge, Q.; Li, J.; Wang, D.J.; Wei, Y.; Zhen, O.Y. Antioxidant and anti-aging activities of polysaccharides from Cordyceps cicadae. Int. J. Biol. Macromol. 2020, 157, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Wang, J.Q.; Fang, Q.Y.; Dong, N.; Nie, S.P. Polysaccharide from natural Cordyceps sinensis ameliorated intestinal injury and enhanced antioxidant activity in immunosuppressed mice. Food Hydrocoll. 2019, 89, 661–667. [Google Scholar] [CrossRef]
- Jin, Y.; Meng, X.; Qiu, Z.D.; Su, Y.P.; Yu, P.; Qu, P. Anti-tumor and anti-metastatic roles of cordycepin, one bioactive compound of Cordyceps militaris. Saudi J. Biol. Sci. 2018, 25, 991–995. [Google Scholar] [CrossRef]
- Qian, G.M.; Pan, G.F.; Guo, J.Y. Anti-inflammatory and antinociceptive effects of cordymin, a peptide purified from the medicinal mushroom Cordyceps sinensis. Nat. Prod. Res. 2012, 26, 2358–2362. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.R.; Zhang, J.Y.; Cheng, X.H.; Zheng, J.X.; Zhang, Q.; Zheng, R.; You, L.Z.; Han, D.R.; Shang, H.C. Immunometabolism at the service of traditional Chinese medicine. Pharmacol. Res. 2022, 176, e106081. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hu, T.; Guo, Y.B.; Liang, Y. Preservation affects the vegetative growth and fruiting body production of Cordyceps militaris. World J. Microbiol. Biotechnol. 2018, 34, e166. [Google Scholar] [CrossRef]
- Shi, H.; Zhang, M.; Devahastin, S. New development of efficient processing techniques on typical medicinal fungi: A Review. Food Rev. Int. 2019, 36, 39–57. [Google Scholar] [CrossRef]
- Tian, Y.; Li, D.M.; Luo, W.H.; Zhu, Z.W.; Li, W.J.; Qian, Z.M.; Li, G.R.; Sun, D.W. Rapid freezing using atomized liquid nitrogen spray followed by frozen storage below glass transition temperature for Cordyceps sinensis preservation: Quality attributes and storage stability. LWT 2020, 123, 109066. [Google Scholar] [CrossRef]
- Qian, Z.M.; Fan, J.J.; Li, C.H.; Yang, F.Q.; Li, W.J.; Zhong, Z.G.; Yao, Z.Z. Effects of different drying conditions on protein in Cordyceps. China J. Chin. Mater. Med. 2019, 44, 1983–1988. [Google Scholar]
- Bok, J.W.; Lermer, L.; Chilton, J.; Klingeman, H.G.; Towers, G.N. Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 1999, 51, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.P.; Wang, J.H.; Liu, X.; Kuang, H.C.; Zhao, S.Y. Simultaneous determination of free ergosterol and ergosteryl esters in Cordyceps sinensis by HPLC. Food Chem. 2007, 105, 1755–1759. [Google Scholar] [CrossRef]
- Barido, F.H.; Jang, A.; Pak, J.I.; Kim, D.Y.; Lee, S.K. Investigation of taste-related compounds and antioxidative profiles of retorted samgyetang made from fresh and dried Cordyceps militaris mushrooms. Food Sci. Anim. Resour. 2020, 40, 772–784. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China 2020; China Medical Science Press: Beijing, China, 2020; pp. 115–117. [Google Scholar]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global metabolic profiling of animal and human tissues via UPLC-MS. Nat. Protoc. 2012, 8, 17–32. [Google Scholar] [CrossRef]
- Barri, T.; Dragsted, L.O. UPLC-ESI-QTOF/MS and multivariate data analysis for blood plasma and serum metabolomics: Effect of experimental artifacts and anticoagulant. Anal. Chim. Acta 2013, 768, 118–128. [Google Scholar] [CrossRef]
- Yang, F.Q.; Feng, K.; Zhao, J.; Li, S.P. Analysis of sterols and fatty acids in natural and cultured Cordyceps by one-step derivatization followed with gas chromatography-mass spectrometry. J. Pharm. Biomed. Anal. 2009, 49, 1172–1178. [Google Scholar] [CrossRef]
- Qiu, X.H.; Li, C.; Han, R.C. Analysis of volatile components in different Ophiocordyceps sinensis and insect host products. Molecules 2020, 25, 1603. [Google Scholar] [CrossRef]
- Hsu, T.H.; Shiao, L.H.; Hsieh, C.; Chang, D.M. A comparison of the chemical composition and bioactive ingredients of the Chinese medicinal mushroom DongChongXiaCao, its counterfeit and mimic, and fermented mycelium of Cordyceps sinensis. Food Chem. 2002, 78, 463–469. [Google Scholar] [CrossRef]
- Gomez, L.L.; Cobas, N.; Franco, I.; Martínez, S.S. Fatty acid profiles and lipid quality indices in canned European eels: Effects of processing steps, filling medium and storage. Food Res. Int. 2020, 136, 109601. [Google Scholar] [CrossRef]
- Wan, W.L.; Xu, J.; Shi, J.Y.; Zhang, X.; Wang, A.; Dong, X.J.; Miao, S.Y. The interaction of dietary niacin and lipid affects the growth, antioxidant capacity and lipid metabolism of male Eriocheir sinensis. Aquac. Rep. 2023, 30, e101541. [Google Scholar] [CrossRef]
- Li, C.; Li, Z.; Fan, M.; Cheng, W.; Long, Y.; Ding, T.; Ming, L. The composition of Hirsutella sinensis, anamorph of Cordyceps sinensis. J. Food Compos. Anal. 2006, 19, 800–805. [Google Scholar] [CrossRef]
- Li, R.; Jiang, X.L.; Guan, H.S. Optimization of mycelium biomass and exopolysaccharides production by Hirsutella sp in submerged fermentation and evaluation of exopolysaccharides antibacterial activity. Afr. J. Biotechnol. 2010, 9, 196–203. [Google Scholar]
- Fan, L.C.; Xian, C.N.; Tang, S.J.; Ding, W.; Xu, C.H.; Wang, X.C. Effect of frozen storage temperature on lipid stability of hepatopancreas of Eriocheir sinensis. LWT 2022, 154, e112513. [Google Scholar] [CrossRef]
- Hama, M.C.; Abdelsamed, I.; Elshamy, T.Y.; Yusuke, K.; Hirohumi, Y.; Kana, T.; Ayari, O.; Masaaki, N.J.; Ban, S.; Hiroshi, I.; et al. New alkaloidal metabolites from cultures of entomopathogenic fungus Cordyceps takaomontana NBRC 101754. Fitoterapia 2019, 139, 104364. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.; Fernie, A.R. Citrulline metabolism in plants. Amino Acids 2017, 49, 1543–1559. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.K.; Kwon, B. Immune regulation through tryptophan metabolism. Exp. Mol. Med. 2023, 55, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Sidney, M.M. Arginine Metabolism Revisited12. J. Nutr. 2016, 146, 2579S–2586S. [Google Scholar]
- María, E.; Alvarez, A.S.; László, S. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar]
- Opeyemi, J.; Olatunji, Y.F.; Oyenike, O.O.; Tang, J.; Wei, Y.; Zhen, O.Y.; Su, Z.L. Polysaccharides purified from Cordyceps cicadae protects PC12 cells against glutamate-induced oxidative damage. Carbohydr. Polym. 2016, 153, 187–195. [Google Scholar]
- Zeng, Z.; Mou, D.; Luo, L.; Zhong, W.; Duan, L.; Zou, X. Different cultivation environments affect the yield, bacterial community and metabolites of Cordyceps cicadae. Front. Microbiol. 2021, 12, 669785. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yue, P.; Tong, X.; Gao, T.; Peng, T.; Guo, J. Comparative analysis of fatty acid metabolism based on transcriptome sequencing of wild and cultivated Ophiocordyceps sinensis. Peer J. 2021, 9, e11681. [Google Scholar] [CrossRef] [PubMed]
- Marcos, J.; Renau, N.; Valverde, O.; Aznar-Lain, G.; Gracia-Rubio, I.; Gonzalez-Sepulveda, M.; Pérez-Jurado, L.A.; Ventura, R.; Segura, J.; Pozo, O.J. Targeting tryptophan and tyrosine metabolism by liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2016, 1434, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.J.; Fan, M.; Liu, Z.G.; Li, X.; Wang, H.J. Serine, glycine and one-carbon metabolism in cancer. Int. J. Oncol. 2020, 11, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Neila, H.M.; Emerson, S.L.; Pritesh, L. Dysregulation of tryptophan catabolism in metabolic syndrome. Metab. Syndr. Relat. Disord. 2018, 16, 135–142. [Google Scholar]
- Brosnan, M.E.; Brosnan, J.T. Histidine metabolism and function. J. Nutr. 2020, 150, 2570S–2575S. [Google Scholar] [CrossRef] [PubMed]
- Shauni, L.G.; Elien, H.; Kim, D.K.; Kim, R.K. The ins and outs of serine and glycine metabolism in cancer. Nat. Metab. 2021, 3, 131–141. [Google Scholar]
- Zhang, X.; Liu, Q.; Zhou, W.; Li, P.; Alolga, R.N.; Qi, L.W.; Yin, X.J. A comparative proteomic characterization and nutritional assessment of naturally and artificially cultivated Cordyceps sinensis. J. Proteom. 2018, 181, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Hyun, S.H.; Lee, S.Y.; Sung, G.H.; Kim, S.H.; Choi, H.K. Metabolic profiles and free radical scavenging activity of Cordyceps bassiana fruiting bodies according to developmental stage. PLoS ONE 2013, 8, e73065. [Google Scholar] [CrossRef]
- Venkatesh, M.; Xi, Q.X.; Goh, E.T.L.; Koh, S.K.; Toh, T.Y.J.; Barathi, V.A.; Mobashar, H.U.T.F.; Madhavi, L.S.C.; Jayasudha, V.; Darren, S.J.T.; et al. Rational substitution of ε-lysine for α-lysine enhances the cell and membrane selectivity of pore-forming melittin. J. Med. Chem. 2020, 63, 3522–3537. [Google Scholar]
- Wang, W.; Meng, X.; Wang, J.; Li, Y. Improved heart failure by rhein lysinate is associated with p38MAPK pathway. Exp. Ther. Med. 2018, 16, 2046–2051. [Google Scholar] [CrossRef]
- Wei, J.; Zhou, X.; Dong, M.; Yang, L.F.; Zhao, C.; Lu, R.L.; Bao, G.H.; Hu, F.L. Metabolites and novel compounds with anti-microbial or antiaging activities from Cordyceps fumosorosea. AMB Expr. 2022, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.G.; Wu, T.; Olatunji, O.J.; Tang, J.; Wei, Y.; Zhen, O.Y. N6-(2-hydroxyethyl)-adenosine from Cordyceps cicadae attenuates hydrogen peroxide induced oxidative toxicity in PC12 cells. Metab. Brain Dis. 2019, 34, 1325–1334. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Tatsuya, K.; Hideo, D.; Enoch, Y.P. Insight into cordycepin biosynthesis of Cordyceps militaris: Comparison between a liquid surface culture and a submerged culture through transcriptomic analysis. PLoS ONE 2017, 12, e0187052. [Google Scholar]
- Liu, Y.; Guo, Z.J.; Zhou, X.W. Chinese Cordyceps: Bioactive components, antitumor effects and underlying mechanism—A review. Molecules 2022, 27, 6576. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | ADC (Relative Abundance) | AFC (Relative Abundance) | WDC (Relative Abundance) | WFC (Relative Abundance) |
|---|---|---|---|---|
| Sn-Glycerol 3-phosphoethanolamine | 1.06 × 107 ± 9.48 × 104 C | 1.11 × 107 ± 9.17 × 104 B | 1.28 × 107 ± 1.43 × 105 A | 7.73 × 106 ± 9.88 × 104 D |
| Valine | 3.61 × 106 ± 2.86 × 104 A | 3.34 × 106 ± 1.22 × 105 B | 3.26 × 106 ± 3.88 × 104 B | 2.34 × 106 ± 3.32 × 104 C |
| Zinniol | 3.72 × 106 ± 3.04 × 105 A | 7.05 × 105 ± 2.70 × 104 B | 3.59 × 106 ± 2.18 × 105 A | 8.88 × 105 ± 7.58 × 104 B |
| Xanthotoxol | 6.65 × 105 ± 3.63 × 104 B | 6.70 × 105 ± 8.54 × 104 B | 5.22 × 105 ± 1.65 × 104 C | 7.80 × 105 ± 3.40 × 104 A |
| NP-018523 | 1.20 × 104 ± 7.87 × 102 C | 8.15 × 102 ± 1.26 × 102 D | 1.52 × 105 ± 6.86 × 103 A | 7.60 × 104 ± 2.56 × 103 B |
| Urocanate | 1.21 × 106 ± 2018 × 104 B | 9.68 × 105 ± 1.20 × 104 C | 1.45 × 106 ± 1.40 × 104 A | 6.61 × 105 ± 9.16 × 103 D |
| CAY-10583 | 6.01 × 105 ± 1.20 × 105 A | 2.00 × 104 ± 1.47 × 103 C | 3.23 × 105 ± 2.01 × 105 AB | 7.28 × 104 ± 4.19 × 103 B |
| Cis-4-Hydroxy-D-proline | 2.37 × 105 ± 2.32 × 103 C | 2.96 × 105 ± 2.72 × 103 B | 6.56 × 105 ± 1.52 × 104 A | 7.11 × 104 ± 8.93 × 102 D |
| Vulpinic acid | 5.63 × 105 ± 3.23 × 104 B | 1.41 × 105 ± 1.41 × 103 C | 7.13 × 105 ± 4.46 × 104 A | 1.77 × 105 ± 4.91 × 103 C |
| Uridine 5’-diphosphate | 5.09 × 105 ± 8.44 × 103 A | 2.78 × 105 ± 8.31 × 103 B | 4.93 × 105 ± 1.72 × 104 A | 1.52 × 105 ± 5.17 × 103 C |
| [6]-shogaol | 3.85 × 105 ± 3.28 × 104 B | 8.80 × 105 ± 1.48 × 104 A | 2.35 × 105 ± 1.07 × 104 C | 9.03 × 104 ± 6.88 × 103 D |
| γ-L-Glutamyl-L-valine | 2.82 × 105 ± 8.87 × 103 C | 3.96 × 105 ± 1.77 × 104 A | 3.38 × 105 ± 1.62 × 104 B | 1.72 × 105 ± 7.52 × 103 D |
| Uridine 5’-monophosphate | 4.39 × 105 ± 5.62 × 103 A | 3.69 × 105 ± 2.87 × 103 B | 3.67 × 105 ± 5.16 × 103 B | 2.04 × 105 ± 1.84 × 103 C |
| Xanthosine | 4.60 × 105 ± 1.82 × 104 A | 8.98 × 104 ± 1.27 × 104 B | 5.87 × 104 ± 3.18 × 103 C | 6.85 × 104 ± 5.10 × 103 BC |
| Zearalenone | 1.09 × 105 ± 1.39 × 104 B | 1.68 × 105 ± 1.60 × 104 A | 1.71 × 105 ± 2.20 × 103 A | 7.05 × 104 ± 3.76 × 103 C |
| Val-Ile | 1.29 × 105 ± 3.11 × 103 B | 6.93 × 104 ± 2.17 × 103 D | 2.75 × 105 ± 6.30 × 103 A | 1.06 × 105 ± 2.81 × 103 C |
| Vitexin 4-o-glucoside | 1.89 × 104 ± 9.78 × 103 B | 9.92 × 104 ± 9.71 × 102 A | 1.33 × 104 ± 5.41 × 103 B | 1.38 × 104 ± 3.66 × 102 B |
| Val-Trp | 9.89 × 103 ± 1.83 × 103 C | 1.54 × 105 ± 9.06 × 103 B | 3.16 × 104 ± 3.49 × 103 C | 5.17 × 105 ± 1.01 × 104 A |
| Wogonin | 7.53 × 104 ± 6.10 × 103 B | 6.88 × 105 ± 5.49 × 104 A | 6.62 × 104 ± 8.28 × 103 B | 6.12 × 105 ± 2.39 × 104 A |
| Valdecoxib | 3.45 × 105 ± 1.09 × 105 A | 2.00 × 105 ± 6.26 × 104 AB | 1.97 × 105 ± 8.74 × 104 AB | 1.15 × 104 ± 3.63 × 102 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Tang, C.; Xiao, M.; Cao, Z.; He, M.; Qi, J.; Li, Y.; Li, X. Effect of Air Drying on the Metabolic Profile of Fresh Wild and Artificial Cordyceps sinensis. Foods 2024, 13, 48. https://doi.org/10.3390/foods13010048
Wang T, Tang C, Xiao M, Cao Z, He M, Qi J, Li Y, Li X. Effect of Air Drying on the Metabolic Profile of Fresh Wild and Artificial Cordyceps sinensis. Foods. 2024; 13(1):48. https://doi.org/10.3390/foods13010048
Chicago/Turabian StyleWang, Tao, Chuyu Tang, Mengjun Xiao, Zhengfei Cao, Min He, Jianzhao Qi, Yuling Li, and Xiuzhang Li. 2024. "Effect of Air Drying on the Metabolic Profile of Fresh Wild and Artificial Cordyceps sinensis" Foods 13, no. 1: 48. https://doi.org/10.3390/foods13010048