
The Relationship between Preparation and Biological Activities of Animal-Derived Polysaccharides: A Comprehensive Review


Abstract
:1. Introduction
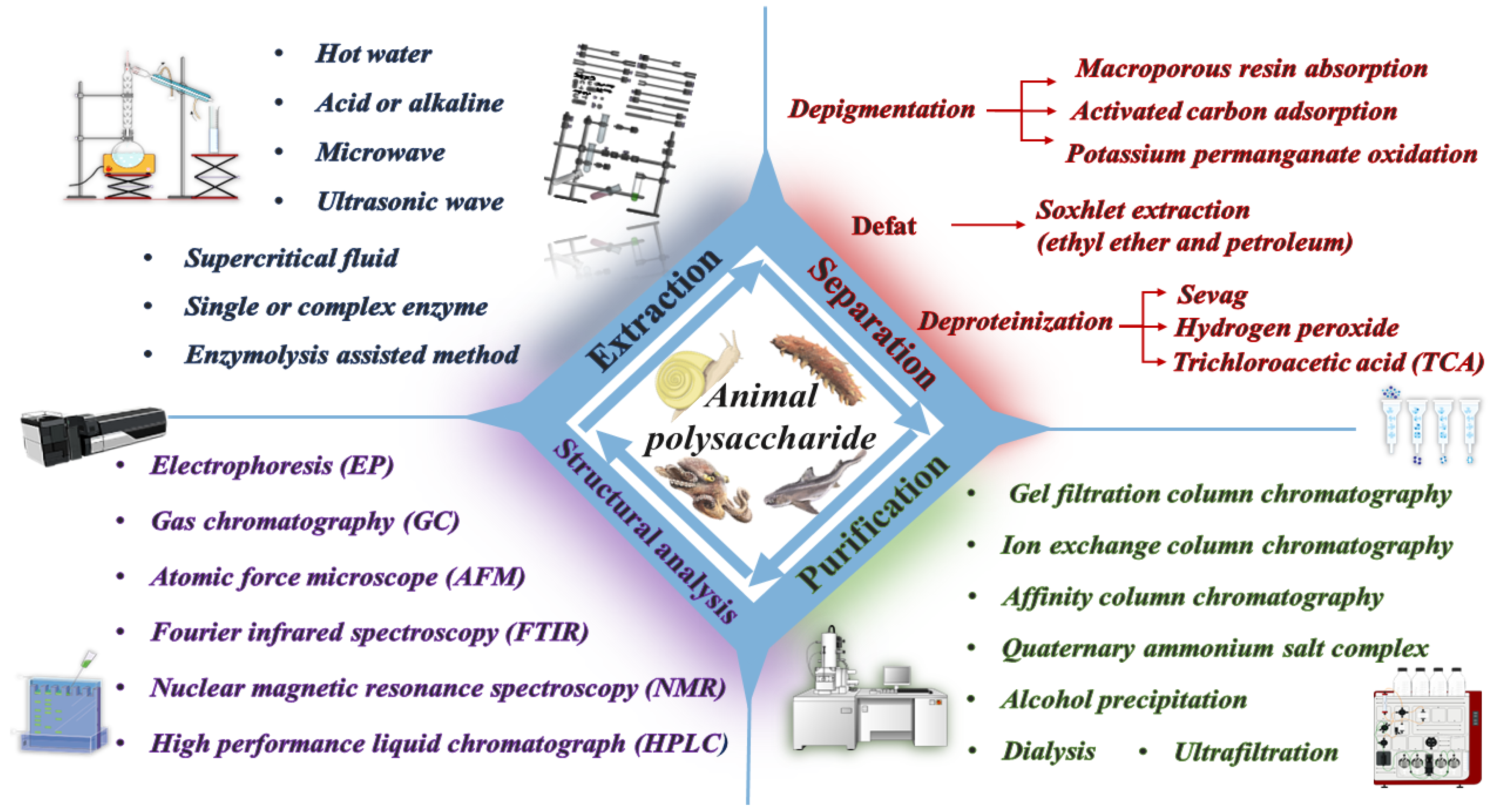
2. Extraction Methods of Animal Polysaccharides
2.1. Water Extraction Method
2.2. Acid and Alkaline Extraction Method
2.3. Enzyme Extraction Method
2.4. Combined Extraction Method
3. Method of Isolation and Purification of Animal Polysaccharides
4. Structural Characterization of Animal Polysaccharides
4.1. Monosaccharide Composition
4.2. Functional Groups and Chemical Bonds
4.3. Disaccharide Composition
4.4. Chain Conformation
5. Biological Activities of Animal Polysaccharides
5.1. Hypoglycemic Activity
5.2. Hypolipidemic Activity
5.3. Anti-Tumor Activity
5.4. Antioxidant Activity
5.5. Anti-Aging Activity
5.6. Antibacterial Activity
5.7. Immunological Activity
5.8. Anticoagulant Activity
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huang, Y.Z.; Chen, H.; Zhang, K.F.; Lu, Y.M.; Wu, Q.Z.; Chen, J.L.; Li, Y.; Wu, Q.X.; Chen, Y. Extraction, purification, structural characterization, and gut microbiota relationship of polysaccharides: A review. Int. J. Biol. Macromol. 2022, 213, 967–986. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, G.L. Antitumor activity of polysaccharides: An overview. Curr. Drug Targets 2018, 19, 89–96. [Google Scholar] [CrossRef]
- Barbosa, J.R.; de Carvalho Junior, R.N. Polysaccharides obtained from natural edible sources and their role in modulating the immune system: Biologically active potential that can be exploited against COVID-19. Trends Food Sci. Technol. 2021, 108, 223–235. [Google Scholar] [CrossRef]
- Wang, P.P.; Zhang, Z.Q.; Chi, L.L.; Yu, G.L. Advances in bioactivity of glycosaminoglycans derived from marine animals. Chin. J. Mar. Drugs 2022, 41, 82–90. [Google Scholar]
- Pomin, V.H.; Mulloy, B. Glycosaminoglycans and Proteoglycans. Pharmaceuticals 2018, 11, 27. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Cai, M.D.; Peng, H.Z.; Liu, M.Y.; Liu, X.N. Effect of glycosaminoglycans with different degrees of sulfation on chondrogenesis. West China J. Stomatol. 2023, 41, 395–404. [Google Scholar]
- Zappe, A.; Miller, R.L.; Struwe, W.B.; Pagel, K. State-of-the-art glycosaminoglycan characterization. Mass Spectrom. Rev. 2021, 41, 1040–1071. [Google Scholar] [CrossRef]
- Wang, L.C.; Di, L.Q.; Li, J.S.; Hu, L.H.; Cheng, J.M.; Wu, H. Elaboration in type, primary structure, and bioactivity of polysaccharides derived from mollusks. Crit. Rev. Food. Sci. Nutr. 2019, 59, 1091–1114. [Google Scholar] [CrossRef]
- Xiong, Q.P.; Song, Z.Y.; Hu, W.H.; Liang, J.; Jing, Y.; He, L.; Huang, S.; Wang, X.L.; Hou, S.Z.; Xu, T.T.; et al. Methods of extraction, separation, purification, structural characterization for polysaccharides from aquatic animals and their major pharmacological activities. Crit. Rev. Food. Sci. Nutr. 2020, 60, 48–63. [Google Scholar] [CrossRef]
- Li, T.; Zhang, J.W.; Lan, W.F.; Yu, L.; Bi, Y.F.; Song, S.X.; Xiong, B.Y.; Wang, H.Y. Polysaccharide decolorization: Methods, principles of action, structural and functional characterization, and limitations of current research. Trends Food Sci. Technol. 2023, 138, 284–296. [Google Scholar]
- Zheng, J.; Sun, K.H.; Gao, X.; Zhang, F.; Sun, P.P.; Hu, A.J.; Gao, J.Z. Extraction and vitro antioxidant activity analysis of polysaccharide from lumpus Cartilage. China Food Addit. 2018, 7, 114–121. [Google Scholar]
- Luan, X.H. Extraction Separation and Structural Characterization of Polysaccharides from Two Species of Solenidae. Master’s Thesis, Ocean University of China, Qingdao, China, 2015. [Google Scholar]
- Ticar, B.F.; Rohmah, Z.; Neri, T.A.N.; Pahila, I.G.; Vasconcelos, A.; Archer-Hartmann, S.A.; Reiter, C.E.N.; Dobruchowska, J.M.; Choi, B.D.; Heiss, C.; et al. Biocompatibility and structural characterization of glycosaminoglycans isolated from heads of silver-banded whiting (Sillago argentifasciata Martin & Montalban 1935). Int. J. Biol. Macromol. 2020, 151, 663–676. [Google Scholar] [PubMed]
- Getachew, A.T.; Lee, H.J.; Cho, Y.J.; Chae, S.J.; Chun, B.S. Optimization of polysaccharides extraction from Pacific oyster (Crassostrea gigas) using subcritical water: Structural characterization and biological activities. Int. J. Biol. Macromol. 2019, 121, 852–861. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.; Alfaro, A.C.; Baroutian, S.; Seyfoddin, A. Extraction of bioactive compounds from black-footed abalone (Haliotis iris) using subcritical water extraction. J. Chem. Technol. Biotechnol. 2022, 97, 3511–3519. [Google Scholar] [CrossRef]
- Chen, D.J.; Xu, W.L.; Su, W.; Ji, X.H. Extraction and monosaccharide composition of Andrias davidianus skin mucus polysaccharide. Nat. Prod. Res. Dev. 2015, 27, 1700–1705. [Google Scholar]
- Lu, Y.Q.; Chi, H.B.; Li, G.N.; Chen, Q.; Qiu, J.G.; Yu, H.; Fang, X.B.; Chen, X.E. Study on alkaline extraction process optimization of polysaccharides from coelomic fluid of Phasolosma esculenta and its antioxidant activity in vitro. Sci. Technol. Food Ind. 2021, 42, 204–210. [Google Scholar]
- Zhuo, X.Y.; Liu, W.H.; Yang, Z.Y. Advances of the animal-original polysaccharides: Extraction, purification and structural identification. Mod. Chem. Res. 2020, 3, 44–47. [Google Scholar]
- Cheng, S.W.; Yu, X.M.; Zhang, Y.X. Study on extraction of polysaccharides from Mytilus edulis and their antioxidant activity in vitro. Sci. Technol. Food Ind. 2010, 31, 132–134. [Google Scholar]
- Dong, Y.; Lin, H.Q.; Miao, S.; Lu, X. Advances in enzymatic extraction of polysaccharides. Sci. Technol. Food Ind. 2021, 42, 351–358. [Google Scholar]
- Maccari, F.; Galeotti, F.; Volpi, N. Isolation and structural characterization of chondroitin sulfate from bony fishes. Carbohydr. Polym. 2015, 129, 143–147. [Google Scholar] [CrossRef]
- Bai, M.Y.; Wang, Q.C.; Deng, S.M.; Zhao, X. Isolation, purification and structural characterization of a novel dermatan sulfate from Lapemis curtus skin. Chin. J. Mar. Drugs 2018, 37, 7–14. [Google Scholar]
- Wang, T.; Zhang, S.L.; Ren, S.Y.; Zhang, X.; Yang, F.; Chen, Y.; Wang, B. Structural characterization and proliferation activity of chondroitin sulfate from the sturgeon, Acipenser schrenckii. Int. J. Biol. Macromol. 2020, 164, 3005–3011. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Ahmadi, S.; Yu, C.X.; Zhang, L.M.; Hu, X.X.; Ye, X.Q.; Chen, S.G. A rapid method for extraction, purification and structure analysis of chondroitin sulfate from six marine tissues. Food Qual. Saf. 2022, 6, 1–10. [Google Scholar] [CrossRef]
- Yuan, J.; Yan, X.T.; Chen, X.; Jiang, X.Q.; Ye, K.Q.; Xiong, Q.P.; Kong, J.; Huang, Y.G.; Jiang, C.X.; Xu, T.T.; et al. A mild and efficient extraction method for polysaccharides from Sinonovacula constricta and study of their structural characteristic and antioxidant activities. Int. J. Biol. Macromol. 2020, 143, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wang, J.; He, C.B.; Wei, H.C.; Ma, Y.; Xiong, H.J. Preparation and antioxidant activities of polysaccharides obtained from abalone viscera by combination of enzymolysis and multiple separation methods. J. Food Sci. 2020, 85, 4260–4270. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Shi, S.; Li, F.F.; Du, X.; Kong, B.H.; Wang, H.; Xia, X.F. Physicochemical properties and antioxidant activity of polysaccharides obtained from sea cucumber gonads via ultrasound-assisted enzymatic techniques. Lebensm.-Wiss. Technol. 2022, 160, 113307. [Google Scholar] [CrossRef]
- Guo, L.; Liu, B.; Wen, C.R.; Song, S.; Zhu, B.W. Qualitative analysis of polysaccharides from different tissues of turbot (Scophthalmus maximus) by HPLC-MS/MS. Food Sci. 2017, 38, 171–178. [Google Scholar]
- Chen, W.W.; Jia, Z.B.; Zhu, J.J.; Zou, Y.R.; Huang, G.R.; Hong, Y. Optimization of ultrasonic-assisted enzymatic extraction of polysaccharides from thick-shell mussel (Mytilus coruscus) and their antioxidant activities. Int. J. Biol. Macromol. 2019, 140, 1116–1125. [Google Scholar] [CrossRef]
- Li, S.; Chen, J.; Song, Z.Y.; Li, Z.B.; Qi, Y.X.; Lu, W.; Zhao, Q.C. Extraction of polysaccharide by salting-out extraction coupled with enzymolysis from abalone viscera and monosaccharide composition analysis. J. Food Saf. Qual. 2017, 8, 4351–4356. [Google Scholar]
- Yu, Q.; Zheng, R.; Zhang, Y.; Li, B.; Liu, D.; Li, Y.P.; Jiao, L.L. Glycosaminoglycan analysis and in vitro immunological activity of the skin of Rana dybowskii Guenther. Chin. Tradit. Pat. Med. 2022, 44, 3728–3732. [Google Scholar]
- Muhindo, M.E.; Wang, Y.J.; Wu, J.F.; Wu, T.; Liu, R.; Sui, W.J.; Zhang, M. Optimization of extraction technology and physicochemical properties of polysaccharide from sea cucumber intestines. Sci. Technol. Food Ind. 2023, 44, 196–203. [Google Scholar]
- Yu, H.M. Extraction of Aspongopus chinensis Polysaccharides and Its Vitro Study on Anti-Breast Cancer In Vitro. Master’s Thesis, Guizhou University, Guiyang, China, 2023. [Google Scholar]
- Dang, C. The Development of Ready-to-Eat Loach Food and Extraction, Structural Analysis of Its Byproduct Polysaccharide. Master’s Thesis, Tianjin University of Science and Technology, Tianjin, China, 2017. [Google Scholar]
- Bai, X.; Gao, X.; Zhao, X.; Yu, M.X.; Hou, H. Preparation and structural analysis of chondroitin sulfate from sturgeon cartilage. Chin. J. Mar. Drugs 2022, 41, 28–36. [Google Scholar]
- Zhu, B.W.; Zhou, D.Y.; Tao, L.; Yan, S.; Yang, J.F.; Li, D.M.; Dong, X.P.; Murata, Y. Chemical composition and free radical scavenging activities of a sulphated polysaccharide extracted from abalone gonad (Haliotis Discus Hannai Ino). Food Chem. 2010, 121, 712–718. [Google Scholar] [CrossRef]
- Dong, F.K.; Quan, X.G.; Wang, Q.B.; Liu, Z.M.; Cui, T.; Wang, W.J.; Tang, D.M.; Zhang, R.M.; Zhang, C.; Wang, H.Y.; et al. Purification, structural characterization, and anticoagulant activity evaluation of chondroitin sulfate from codfish (Gadus macrocephalus) bones. Int. J. Biol. Macromol. 2022, 210, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.D.; Lin, F.D.; Huang, Y.Y.; Ye, J.; Xiao, M.T. Separation, purification, structural analysis and immune-enhancing activity of sulfated polysaccharide isolated from sea cucumber viscera. Int. J. Biol. Macromol. 2020, 155, 1003–1018. [Google Scholar] [CrossRef]
- Peng, C.E.; Wang, Q.B.; Jiao, R.M.; Xu, Y.Y.; Han, N.H.; Wang, W.S.; Zhu, C.X.; Li, F.C. A novel chondroitin sulfate E from Dosidicus gigas cartilage and its antitumor metastatic activity. Carbohydr. Polym. 2021, 262, 117971. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.G.; Xue, C.H.; Liang, Y.; Tang, Q.J.; Yu, G.L.; Chai, W.G. Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers. Carbohydr. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Li, X.M.; Li, L.; Cai, Y.; Yang, W.J.; Lin, L.S.; Li, Z.; Gao, N.; Purcell, S.; Wu, M.Y.; Zhao, J.H. Structural elucidation and biological activity of a highly regular fucosylated glycosaminoglycan from the edible sea cucumber Stichopus herrmanni. J. Agric. Food Chem. 2017, 65, 9315–9323. [Google Scholar] [CrossRef]
- Zhan, W.Q.; Liu, S.J.; Li, S.G.; Cai, S.L.; Lu, H.X.; Wang, Y.; Liu, Z.Y. Isolation, purification and structural characterization of glycosaminoglycans from Ruditapes philippinarum. Food Res. Dev 2022, 43, 31–37. [Google Scholar]
- Lin, J.Y.; Wang, J.; Zhang, P.P. Extraction and purification of glycosaminoglycan from Razor clam. J. Chin. Inst. Food Sci. Technol. 2014, 14, 121–128. [Google Scholar]
- Sun, X.P. Studies on the Extraction, Purification and Physiological Activity of Glycosaminoglycan from Mactra veneriformis. Master’s Thesis, Tianjin University of Science and Technology, Tianjin, China, 2011. [Google Scholar]
- Xu, J.L. Studies on the Extraction, Purification and Functional Activities of Glycosaminoglycan from Bullacta exarata. Master’s Thesis, Tianjin University of Science and Technology, Tianjin, China, 2012. [Google Scholar]
- Song, S.; Wang, L.L.; Wang, L.L.; Yu, Q.; Ai, C.Q.; Fu, Y.H.; Yan, C.H.; Wen, C.R.; Zhu, Z.J. Structural characterization and anticoagulant activity of two polysaccharides from Patinopecten yessoensis viscera. Int. J. Biol. Macromol. 2019, 136, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Zuo, G.G. Preparation, Structural Characterization and Antiaging Activity of Chondroitin Sulfate from Different Parts of Tilapia by-Product. Master’s Thesis, Guangdong Ocean University, Guangzhou, China, 2022. [Google Scholar]
- Zhao, S.W.; Qin, Y.; Wang, M.; Zhang, Y.; Jia, X.C.; Liu, Y.; Wang, S.M. Extraction, Isolation and Antioxidant Activity Analysis of Glycosaminoglycan from Chinese White Jade Snail. Shandong Chem. Ind. 2023, 52, 42–45. [Google Scholar]
- Sun, L.; Shi, X.M.; Cai, X.H.; Li, B.; Wang, H.P.; Liu, J.B.; Tang, S.S. Study on extraction and moisture retention of buckskin polysaccharides. J. Jilin Agric. Univ. 2019, 41, 207–212. [Google Scholar]
- Mou, J.J.; Zhuang, D.D.; Li, Q.; Song, W.G.; Yang, J. Comparison of chain conformation properties of bioactive fucosylated chondroitin sulfates from two different sea cucumbers. Int. J. Biol. Macromol. 2019, 133, 44–50. [Google Scholar] [CrossRef]
- Wang, Q.Y. Isolation, Structural Characterization and Mechanism of Immunoregulatory Active Polysaccharides from Sea Urchin and Deep-Sea Bacteria. Ph.D. Thesis, University of Chinese Academy of Sciences (Institute of Oceanography, Chinese Academy of Sciences), Beijing, China, 2023. [Google Scholar]
- Gao, D.X.; Zhou, L.X.; Deng, T.; Fu, X.R.; Wu, Y.B.; Yang, L.; Luo, L.; Hu, L.; Wu, M.Y. Chemical structure of glycosaminoglycan from the snail Achatina fulica mucus and its anticoagulant activity. J. Yunnan Natl. Univ. Nat. Sci. Ed. 2023, 32, 6–12. [Google Scholar]
- Xiong, Q.P. Preparation, Characterization of Sulfate Polysaccharides from Cipangopaludina chinensis and Its Action to Stabilize Atherosclerotic Plaques. Ph.D. Thesis, Guangzhou University of Chinese Medicine, Guangzhou, China, 2020. [Google Scholar]
- Chen, S.J.; Chen, H.; Gao, R.C.; Li, L.H.; Yu, G.; Hu, X.; Ma, H.X.; Deng, J.C. Extraction of hyaluronic acid from tilapia by ultrasonic assisted enzymatic hydrolysis. Acta Agric. Nucl. Sin. 2014, 28, 1446–1452. [Google Scholar]
- Shi, L. Bioactivities, isolation and purification methods of polysaccharides from natural products: A review. Int. J. Biol. Macromol. 2016, 92, 37–48. [Google Scholar] [CrossRef]
- Tang, W.; Liu, D.; Yin, J.D.; Nie, S.P. Consecutive and progressive purification of food-derived natural polysaccharide: Based on material, extraction process and crude polysaccharide. Trends Food Sci. Technol. 2020, 99, 76–87. [Google Scholar] [CrossRef]
- Huang, G.L.; Chen, F.; Yang, W.J.; Huang, H.L. Preparation, deproteinization and comparison of bioactive polysaccharides. Trends Food Sci. Technol. 2021, 109, 564–568. [Google Scholar] [CrossRef]
- Xin, E.N.; Wang, Y.; Li, Y.T.; Shao, T.; Du, H.L.; Zhang, T.B.; Guo, J.H. Preliminary purification and in vitro antioxidant activity of crude polysaccharides from Tenebrio molitor. Food Sci. Technol. 2022, 47, 221–226. [Google Scholar]
- Yu, S.; Li, S.S.; Song, Z.Y.; Sang, X.; Li, Y.; Fan, X.R.; Zhao, Q.C. Research on extraction separation purification methods of sea cucumber polysaccharides. Acad. Period. Farm Prod. Process. 2022, 7, 80–83. [Google Scholar]
- Li, S.Y.; Han, Q.; Su, Y.W.; Qi, Y.X.; Zhao, Q.C.; Li, Z.B. Physicochemical properties and antioxidant activity of graded ethanol precipitation polysaccharide from cooking liquor of Manila clam Ruditapes philippinarum. J. Dalian Ocean Univ. 2018, 33, 658–665. [Google Scholar]
- Dedhia, N.; Marathe, S.J.; Singhal, R.S. Food polysaccharides: A review on emerging microbial sources, bioactivities, nanoformulations and safety considerations. Carbohydr. Polym. 2022, 287, 119355. [Google Scholar] [CrossRef] [PubMed]
- Sha, X.M.; Li, J.W.; Hu, Z.Z.; Zhang, L.J.; Xie, Z.H.; Lu, L.; Yuan, T.; Tu, Z.C. Traceability and identification of fish gelatin from seven cyprinid fishes by high performance liquid chromatography and high-resolution mass spectrometry. Food Chem. 2023, 400, 133961. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.; Yin, Y.D.; Nie, S.P.; Xie, M.Y. Applications of infrared spectroscopy in polysaccharide structural analysis: Progress, challenge and perspective. Food Chem. X 2021, 12, 100168. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.C.; Yuan, S.M.; Zhang, Q.; Luo, M.; Qiao, D.L.; Jiang, F.T.; Qian, H. Improved sample preparation method on the morphology observation of hydrophilic polysaccharides for atomic force microscopy (AFM). Food Hydrocoll. 2023, 144, 109048. [Google Scholar] [CrossRef]
- Speciale, I.; Notaro, A.; Garcia-Vello, P.; Lorenzo, F.D.; Armiento, S.; Molinaro, A.; Marchetti, R.; Silipo, A.; De Castro, C. Liquid-state NMR spectroscopy for complex carbohydrate structural analysis: A hitchhiker’s guide. Carbohydr. Polym. 2022, 277, 118885. [Google Scholar] [CrossRef]
- Miao, W.; Li, N.; Wu, J.L. Food-polysaccharide utilization via in vitro fermentation: Microbiota, structure, and function. Curr. Opin. Food Sci. 2022, 48, 100911. [Google Scholar] [CrossRef]
- Yim, S.K.; Kim, K.; Kim, I.; Chun, S.H.; Oh, T.H.; Kim, J.U.; Kim, J.W.; Jung, W.H.; Moon, H.; Ku, B. Inhibition of SARS-CoV-2 virus entry by the crude polysaccharides of seaweeds and abalone viscera in vitro. Mar. Drugs 2021, 19, 219. [Google Scholar] [CrossRef]
- Wang, Z.C.; Zheng, Y.; Lai, Z.R.; Hu, X.L.; Wang, L.; Wang, X.Q.; Li, Z.T.; Gao, M.J.; Yang, Y.H.; Wang, Q. Effect of monosaccharide composition and proportion on the bioactivity of polysaccharides: A review. Int. J. Biol. Macromol. 2023, 254, 127955. [Google Scholar] [CrossRef]
- Wang, H.Y.; Qin, J.; Zhuang, D.; Liu, C.Y.; Zhang, Y.; Zhang, L.H.; Lv, G.P. Isolation, characterization, antioxidant and immunomodulating activity of polysaccharides from Achatina fulica mucus. Chin. J. Bioprocess Eng. 2020, 18, 324–331. [Google Scholar]
- Krichen, F.; Karoud, W.; Sila, A.; Abdelmalek, B.E.; Ghorbel, R.; Ellouz-Chaabouni, S.; Bougatef, A. Extraction, characterization and antimicrobial activity of sulfated polysaccharides from fish skins. Int. J. Biol. Macromol. 2015, 75, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.Y.Y.; Wang, J.Q.; Yin, J.Y.; Nie, S.P.; Xie, M.Y. A review of NMR analysis in polysaccharide structure and conformation: Progress, challenge and perspective. Food Res. Int. 2021, 143, 110290. [Google Scholar] [CrossRef] [PubMed]
- Seedevi, P.; Moovendhan, V.; Vairamai, S.; Shanmugam, A. Mucopolysaccharide from cuttlefish: Purification, chemical characterization and bioactive potential. Carbohydr. Polym. 2017, 167, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.Y.; Sun, X.J.; Chu, Q.; Zhu, W.Q.; Li, L.; Zheng, Z.J. Research progress in hypoglycemic effect and mechanism of animal polysaccharides. J. Food Saf. Qual. 2020, 11, 7674–7683. [Google Scholar]
- Xiang, X.W.; Jiang, Q.H.; Yang, H.S.; Zhou, X.X.; Chen, Y.F.; Chen, H.; Liu, S.L.; Chen, L. A review on shellfish polysaccharides: Extraction, characterization and amelioration of metabolic syndrome. Front. Nutr. 2022, 9, 974860. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Jia, X.; Wang, N.F.; Xiao, M.; Song, S.; Wu, S.F.; Li, Z.J.; Wang, S.J.; Cui, S.W.; Guo, Q.B. Insights into the structure-bioactivity relationships of marine sulfated polysaccharides: A review. Food Hydrocoll. 2022, 123, 107049. [Google Scholar] [CrossRef]
- Lu, W.J.; Yang, Z.F.; Chen, J.; Wang, D.; Zhang, Y. Recent advances in antiviral activities and potential mechanisms of sulfated polysaccharides. Carbohydr. Polym. 2021, 272, 118526. [Google Scholar] [CrossRef]
- Zhou, J.; Yan, J.Y.; Bai, Z.S.; Li, K.C.; Huang, K.X. Hypoglycemic activity and potential mechanism of a polysaccharide from the loach in streptozotocin-induced diabetic mice. Carbohydr Polym. 2015, 121, 199–206. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, Y.; Ye, X.Q.; Hu, Y.Q.; Ding, T.; Chen, S.G. Sulfation pattern of fucose branches affects the anti-hyperlipidemic activities of fucosylated chondroitin sulfate. Carbohydr. Polym. 2016, 147, 1–7. [Google Scholar] [CrossRef]
- Wu, R.Y.; Li, P.L.; Wang, Y.; Su, N.; Xiao, M.Y.; Li, X.J.; Shang, N. Structural analysis and anti-cancer activity of low-molecular-weight chondroitin sulfate from hybrid sturgeon cartilage. Carbohydr. Polym. 2022, 275, 118700. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, Y.H.; Hu, J.F.; Lv, Z.H. The research of glycosaminoglycans from Apostichopus Japonius on specific immune adjustment. Chin. J. Mar. Drugs 2015, 34, 53–58. [Google Scholar]
- Wang, J.Y.; He, Y.H.; Ma, Z.C.; Liu, X.; Li, X.X.; Wang, Q.K.; Ren, D.D.; Liu, S. Research progress on hypoglycemic mechanism of fucoidan. Chin. J. Mar. Drugs. 2023, 42, 71–80. [Google Scholar]
- Li, S.; Li, J.H.; Zhi, Z.J.; Wei, C.Y.; Wang, W.J.; Ding, T.; Ye, X.Q.; Hu, Y.Q.; Linhardt, R.J.; Chen, S.G. Macromolecular properties and hypolipidemic effects of four sulfated polysaccharides from sea cucumbers. Carbohydr. Polym. 2017, 173, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.X. Anti-Obesity Activity and Mechanism of Chondroitin Sulfate in Sturgeon Skull. Master’s Thesis, Jiangsu University, Nanjing, China, 2023. [Google Scholar]
- Gharibzahedi, S.M.T.; Marti-Quijal, F.J.; Barba, F.J.; Altintas, Z. Current emerging trends in antitumor activities of polysaccharides extracted by microwave- and ultrasound-assisted methods. Int. J. Biol. Macromol. 2022, 202, 494–507. [Google Scholar] [CrossRef]
- Li, R. Research of Anti-Tumor Activity and Immunomodulatory Effect of Glycosaminoglycan from Perna viridis. Master’s Thesis, Guangdong Ocean University, Guangzhou, China, 2010. [Google Scholar]
- Zhong, Q.W.; Wei, B.; Wang, S.J.; Ke, S.Z.; Chen, J.W.; Zhang, H.W.; Wang, H. The antioxidant activity of polysaccharides derived from marine organisms: An overview. Mar. Drugs. 2019, 17, 674. [Google Scholar] [CrossRef]
- Fernandes PA, R.; Coimbra, M.A. The antioxidant activity of polysaccharides: A structure-function relationship overview. Carbohydr. Polym. 2023, 314, 120965. [Google Scholar] [CrossRef]
- Guo, X.L.; Luo, J.J.; Qi, J.Y.; Zhao, X.Y.; An, P.; Luo, Y.T.; Wang, G.S. The role and mechanism of polysaccharides in anti-aging. Nutrients 2022, 14, 5330. [Google Scholar] [CrossRef]
- Gálvez-Martín, P.; Fernández, S.C.; Romero-Rueda, J.; Cabañas, J.; Torrent, A.; Castells, G.; Martinez-Puig, D. A novel hyaluronic acid matrix ingredient with regenerative, anti-aging and antioxidant capacity. Int. J. Mol. Sci. 2023, 24, 4774. [Google Scholar] [CrossRef]
- Alcântara, L.O.; Sousa, J.R.; Andrade, F.K.; Teixeira, E.H.; Cerqueira, M.Ă.; da Silva, A.L.C.; Filho, M.M.S.; de Souza, B.W.S. Extraction and characterization of hyaluronic acid from the eyeball of Nile Tilapia (Oreochromis niloticus). Int. J. Biol. Macromol. 2023, 226, 172–183. [Google Scholar] [CrossRef]
- Huang, G.L. The antiviral activity of polysaccharides and their derivatives. Int. J. Biol. Macromol. 2018, 115, 77–82. [Google Scholar]
- Jridi, M.; Nasri, R.; Marzougui, Z.; Abdelhedi, O.; Hamdi, M.; Nasri, M. Characterization and assessment of antioxidant and antibacterial activities of sulfated polysaccharides extracted from cuttlefish skin and muscle. Int. J. Biol. Macromol. 2019, 123, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.X.; Shen, M.Y.; Morris, G.A.; Xie, J.H. Sulfated polysaccharides: Immunomodulation and signaling mechanisms. Trends Food Sci. Technol. 2019, 92, 1–11. [Google Scholar] [CrossRef]
- Briard, B.; Fontaine, T.; Samir, P.; Place, D.E.; Muszkieta, L.; Malireddi, R.K.S.; Karki, R.; Chrisstgen, S.; Bomme, P.; Vogel, P.; et al. Galactosaminogalactan activates the inflammasome to provide host protection. Nature 2020, 588, 688–692. [Google Scholar] [CrossRef]
- Yin, Y.G.; Han, Y.Z.; Ding, H.W. Review on advancement of animal polysaccharides. Food Sci. 2006, 3, 256–263. [Google Scholar]
- de Moura, H.C.; Novello, C.R.; Balbinot-Alfaro, E.; Düsman, E.; Barddal, H.P.O.; Almeida, I.V.; Vicentini, V.E.P.; Prentice-Hernández, C.; Alfaro, A.T. Obtaining glycosaminoglycans from tilapia (Oreochromis niloticus) scales and evaluation of its anticoagulant and cytotoxic activities. Food Res. Int. 2021, 140, 110012. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, H.; Capitani, F.; Krichen, F.; Mantovani, V.; Amor, I.B.; Galeotti, F.; Maccari, F.; Nedjar, N.; Volpi, N.; et al. Studies on European eel skin sulfated glycosaminoglycans: Recovery, structural characterization and anticoagulant activity. Int. J. Biol. Macromol. 2018, 115, 891–899. [Google Scholar] [CrossRef]
- Liu, X.X.; Hao, J.J.; Shan, X.D.; Zhang, X.; Zhao, X.L.; Li, Q.Y.; Wang, X.J.; Cai, C.; Li, G.Y.; Yu, G.L. Antithrombotic activities of fucosylated chondroitin sulfates and their depolymerized fragments from two sea cucumbers. Carbohydr. Polym. 2016, 152, 343–350. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sources | Type | Extraction Methods | Purification Methods | Crude Extraction Rate (%) | Molecular Weights (kDa) | References |
|---|---|---|---|---|---|---|
| Skin of Rana dybowskii Guenther | GAG | HW | SepharoseTMCL-6B | - | 141 | [31] |
| Sea cucumber intestines | Polysaccharide | HW | - | 4.43 | 4.91 × 106 | [32] |
| Aspongopus chinensis | Polysaccharide | HW | - | 5.067 | - | [33] |
| Loach head | Polysaccharide | A | Sephadex G150 | 2.41 | 7.11 × 105 | [34] |
| Sturgeon cartilage | CS | E | - | 5.7 | 111.48 | [35] |
| Abalone gonad | Sulfated polysaccharide | PE | Source 30Q and Sephacryl-S 100,200 | - | 12.5 | [36] |
| Codfish (Gadus macrocephalus) bones | CS | APE | DEAE Sepharose™Fast Flow | 0.05 | 12.3 | [37] |
| Sea cucumber viscera | Sulfated polysaccharide | PE | DEAE-52 Sephadex G-200 | 4.9 | 18.8 | [38] |
| Dosidicus gigas cartilage | CS E | Actinase E enzymolysis | DEAE-Sephadex | 15.3 | 696 | [39] |
| P. graeffei S. tremulus H. vagabunda I. badionotus (sea cucumber body wall) | fCS | PE | DEAE-52 | 11.0 6.3 7.0 9.9 | 73 81 100 109 | [40] |
| Body wall of sea cucumber Stichopus herrmanni | FG | PE | FPA98 (OH-) | 7.5 | 6.37 × 104 | [41] |
| Ruditapes philippinarum | GAG | NPE | DEAE-650M | - | - | [42] |
| Razor clam | GAG | TE | DEAE-52 | 2.51 | - | [43] |
| Mactra veneriformis | GAG | NP and TE | - | 0.448 | - | [44] |
| Bullacta exarata | GAG | N, P, and TE | - | 0.265 | - | [45] |
| Patinopecten yessoensis viscera | Polysaccharide | T and PSE | DEAE, Sepharose CL-6B | 1 | 63 | [46] |
| Tilapia head, fins, spine, and tail | CS | Savinase 16L and 2709 alkaline protease SE | - | 1.02 0.81 0.38 0.93 | 1.94065 × 105 1.30967 × 105 1.07574 × 105 1.61347 × 105 | [47] |
| Muscle, viscera, and ovum of Chinese White Jade Snail | GAG | T and PSE | - | 1.961 0.5731 0.7759 | - | [48] |
| Buckskin | Polysaccharide | AP and PSE | - | 1.231 | - | [49] |
| A. japonicus and H. mexicana (sea cucumber body wall) | fCS | A—TCE | Q Sepharose Fast Flow | - | 54.3 56.9 | [50] |
| Sea urchin gonad | Polysaccharide | HW—E/HW—A—E | DEAE Fast Flow Sephadex G75, G200 | 11.1 6.2 9.4 9.0 | 4–600 | [51] |
| Snail Achatina fulica mucus | GAG | AP—ACE | FPA98 (OH-) | 10 | 3.343 × 105 | [52] |
| Cipangopaludina chinensis | Polysaccharide | Ultrasonic synchronous coupling papain enzymolysis | Q Sepharose Fast Flow Sephacryls-400 | 13.57 | 91.1 | [53] |
| Vitreous body tilapia | HA | Ultrasonic—compound proteinase enzymolysis | HiTrapTM DEAE-FF | 11.44 | 100 | [54] |
| Sources | Type | Monosaccharide Composition | Proportion of Monosaccharides | References |
|---|---|---|---|---|
| Skin of Rana dybowskii Guenther | GAG | Man, GlcA, Glc, GalN, and Gal | 0.41:2.67:0.16:1:0.25 | [31] |
| Sea cucumber intestines | Polysaccharide | Man, Ara, Gal, Glc, and Fuc | 22.3%, 19.31%, 11.78%, 3.22%, and 42.57% | [32] |
| Abalone gonad | Sulfated polysaccharide | Gal, Fuc, and Rha | 46.9%, 32.7%, and 20.4%, | [36] |
| Sea cucumber viscera | Sulfated polysaccharide | Man, GlcN, GlcA, GalNAc, Glc, Gal, and Fuc | 1.00:1.41:0.88:2.14:1.90:1.12:1.24 | [38] |
| P. graeffei S. tremulus H. vagabunda I. badionotus (sea cucumber body wall) | fCS | GlcA, GalNAc, and Fuc | 1.0:0.8:1.5 1.0:1.1:0.9 1.0:0.8:1.2 1.0:0.7:0.9 | [40] |
| Body wall of sea cucumber Stichopus herrmanni | FG | GlcA, GalNAc, and Fuc | 1:1.13:1.09 | [41] |
| Sea urchin gonad | Polysaccharide | Man, GlcN, and Glc | 5.18:1.00:4.13 7.78:1.91:1.00 5.06:1.00:8.01 | [51] |
| Snail Achatina fulica mucus | GAG | GalNAc and IdoA | - | [52] |
| Sources | Type | Experimental Model | In Vitro/In Vivo | Experimental Data | Biological Activity | References |
|---|---|---|---|---|---|---|
| Skin of Rana dybowskii Guenther | GAG | Mice | In vitro | 50–400 μg/mL, T-lymphocyte proliferation ↑ 200–400 μg/mL, B-lymphocyte proliferation ↓ | Immunological activity | [31] |
| Aspongopus chinensis | Polysaccharide | Mice breast cancer cells 4T1 Human breast cancer cells MDA-MB-453, MCF-7 | In vitro | 24 h, IC50: 10.260, 14.170, and 6.986 mg/mL 48 h, IC50: 5.719, 8.99, 6.302 mg/mL | Anti-tumor activity | [33] |
| Codfish (Gadus macrocephalus) bones | CS | - | In vitro | 0.5 μg/mL, clotting times: 52 s and 25 s 50 μg/mL, clotting time: 46.3 s and 19.9 s | Anticoagulant activity | [37] |
| Sea cucumber viscera | Sulfated polysaccharide | Mice macrophage cells RAW264.7 | In vitro | 400 μg/mL, IL-1β,IL-6,TNF-α: 28.79, 289.36, and 673.56 pg/mL ↑ | Immunological activity | [38] |
| Dosidicus gigas cartilage | CS E | Mice | In vivo | 50–100 μg/mouse, 4T1 cancer cells metastasizing to the lung ↓ | Anti-tumor activity | [39] |
| Tilapia head, fins, spine, and tail | CS | Caenorhabditis elegans | In vivo | 0–1 mg/mL, lifespan of C. elegans increased from 12.33 to 18.50 h | Anti-aging activity | [47] |
| Sea urchin gonad | Polysaccharide | Macrophage cells RAW264.7 | In vitro | 6.25–800 μg/mL, IL-6, COX-2, TNF-α ↑ | Immunological activity | [51] |
| Skin of gray triggerfish and smooth hound | sulfated polysaccharide | - | - | 50 mg/mL, Enterobacter sp., Salmonella enterica ↓ 150 mg/mL, Alternaria solani, Fusarium solani ↓ | Antibacterial activity | [72] |
| Loach (Misgurnus anguillicaudatus) | Polysaccharide | Mice | In vivo | 100 and 200 mg/kg, serum insulin levels modest but significant elevation | Hypoglycemic activity | [77] |
| Sea cucumber | sulfated polysaccharides | Mice | In vivo | 40 mg/kg, HDL-C levels, 67.7%, 36.7%, and 25.8% ↑ LDL-C levels, 13.2%, 32.9%, and 19.8% ↓ | Hypolipidemic activity | [78] |
| Hybrid sturgeon cartilage | SCS | Mice | In vivo | 800 μg/g, tumor biomarkers CEA decreased from 1237.22% to 612.48%, expression of CA19-9 decreased from 57.37% to 17.95% | Anti-tumor activity | [79] |
| Apostichopus Japonius | GAG | Mice | In vitro | 0.5–10 μg/mL, Splenic lymphocytes proliferation ↑ | Immunological activity | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Yang, C.; Liu, R.; Sui, W.; Zhu, Q.; Jin, Y.; Wu, T.; Zhang, M. The Relationship between Preparation and Biological Activities of Animal-Derived Polysaccharides: A Comprehensive Review. Foods 2024, 13, 173. https://doi.org/10.3390/foods13010173
Yang B, Yang C, Liu R, Sui W, Zhu Q, Jin Y, Wu T, Zhang M. The Relationship between Preparation and Biological Activities of Animal-Derived Polysaccharides: A Comprehensive Review. Foods. 2024; 13(1):173. https://doi.org/10.3390/foods13010173
Chicago/Turabian StyleYang, Bochun, Conghao Yang, Rui Liu, Wenjie Sui, Qiaomei Zhu, Yan Jin, Tao Wu, and Min Zhang. 2024. "The Relationship between Preparation and Biological Activities of Animal-Derived Polysaccharides: A Comprehensive Review" Foods 13, no. 1: 173. https://doi.org/10.3390/foods13010173