Pro- and Antioxidant Effect of Food Items and Matrices during Simulated In Vitro Digestion


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, including Enzymes Used for the Digestions and Analyses
2.2. Food Items and Matrices
2.3. In Vitro Digestion
2.3.1. Stock Solutions and Simulated Digestive Fluids
2.3.2. Simulated Gastrointestinal Digestion
2.3.3. Gastric Phase of Digestion
2.3.4. Intestinal Phase of Digestion
2.4. Antioxidant Capacity Measurements
2.4.1. FRAP
2.4.2. ABTS
2.4.3. Peroxide Formation
2.4.4. MDA
2.4.5. DPPH
2.4.6. Anti-Pro-Oxidant Score
2.5. Data Treatment and Statistical Analysis
3. Results
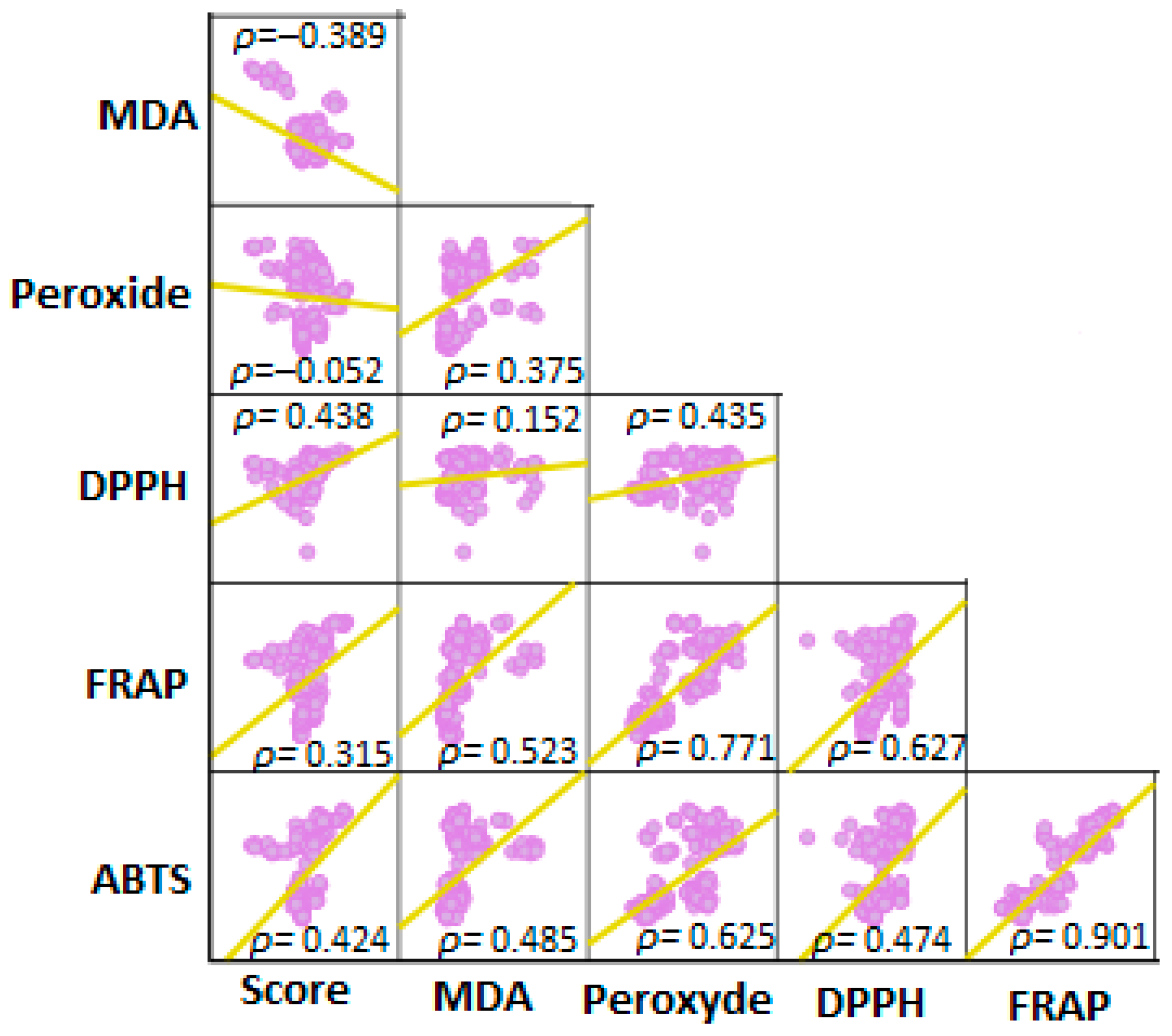
3.1. Data Distribution and Results from the Overall Multivariate Test
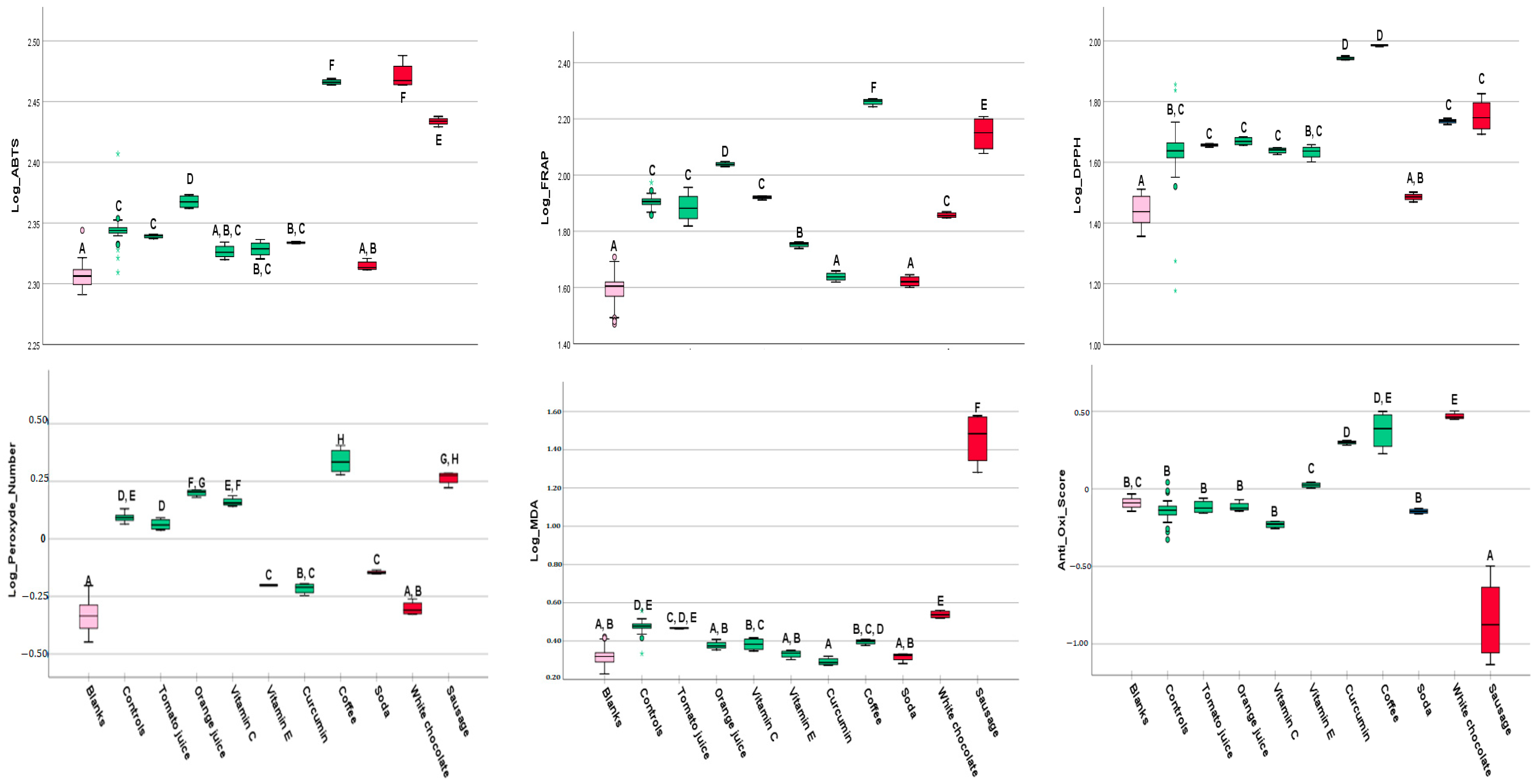
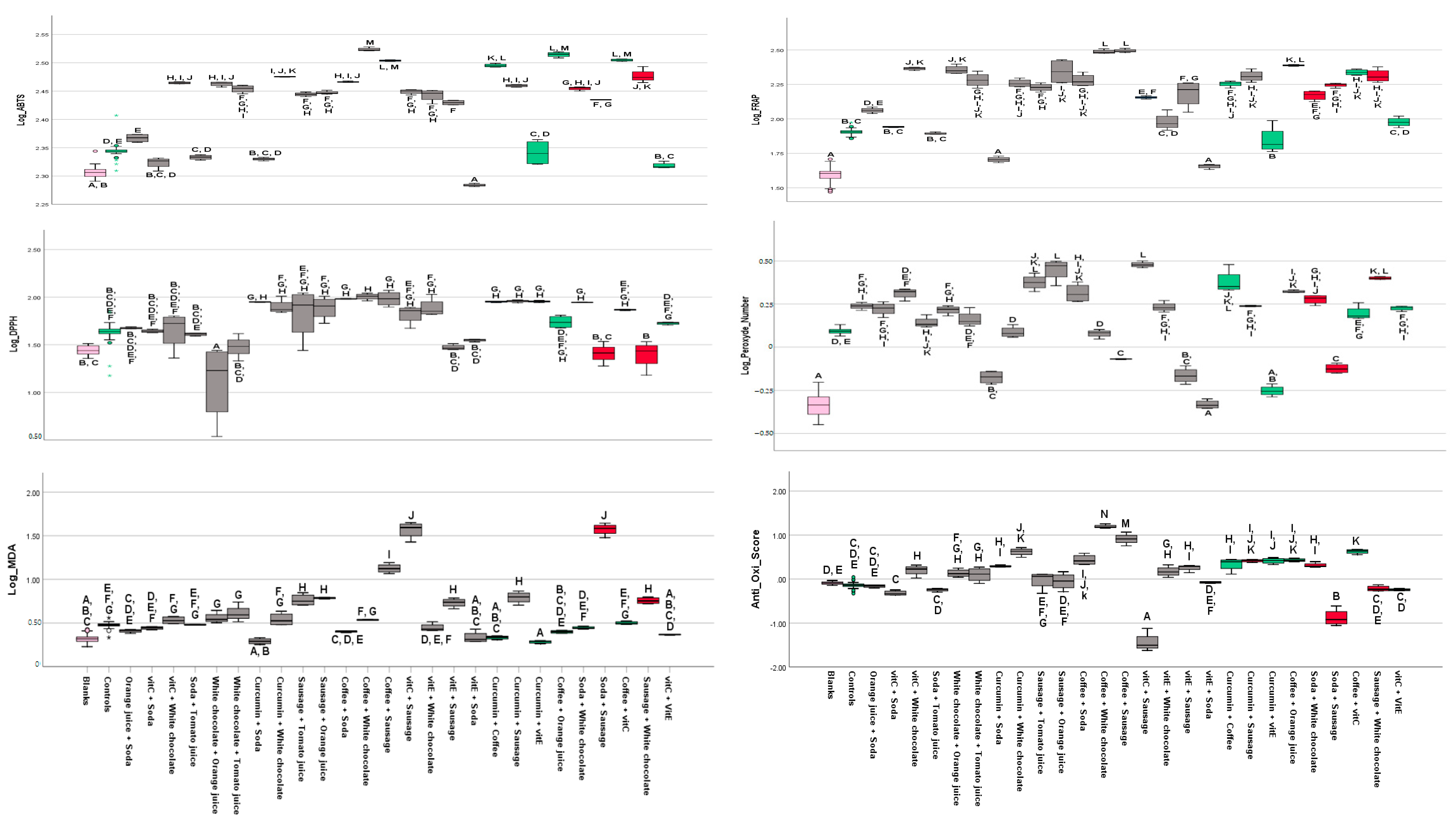
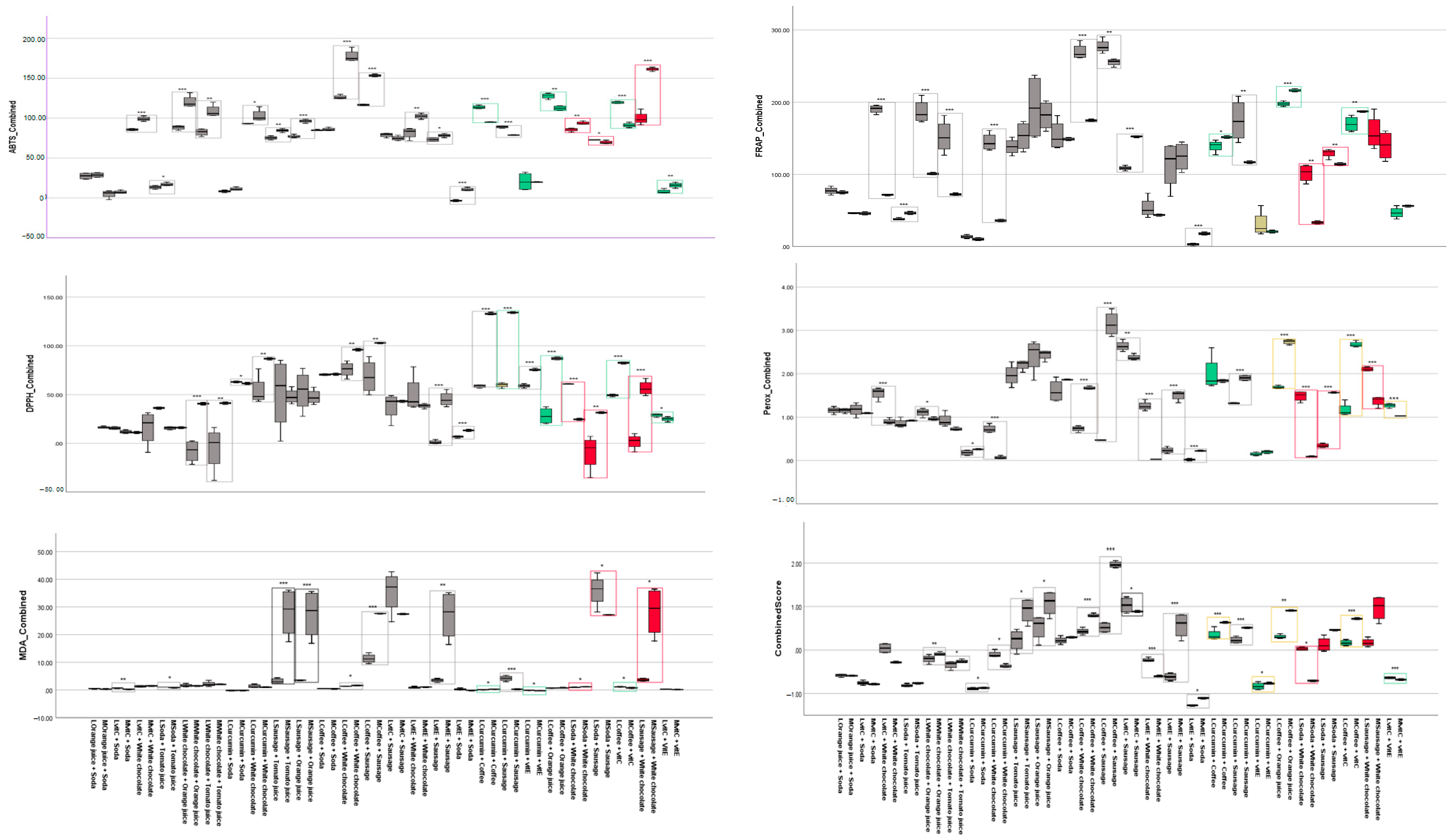
3.2. Findings from Assays
3.2.1. FRAP
3.2.2. ABTS
3.2.3. DPPH

3.2.4. MDA
3.2.5. Peroxide Assay


4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxid. Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural. Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 45–63. [Google Scholar]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.K. Does the Interdependence between Oxidative Stress and Inflammation Explain the Antioxidant Paradox? Oxid. Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef]
- Vahid, F.; Shivappa, N.; Hekmatdoost, A.; Hebert, J.R.; Poustchi, H.; Shamsipour, A.; Eslamparast, T.; Meibodi, M.; Rahmani, D. Association of Pro-inflammatory Dietary Intake and Non-Alcoholic Fatty Liver Disease: Findings from Iranian case-control study. Int. J. Vitam. Nutr. Res. 2018, 88, 144–150. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Patlevič, P.; Vašková, J.; Švorc, P., Jr.; Vaško, L.; Švorc, P. Reactive oxygen species and antioxidant defense in human gastrointestinal diseases. Int. Med. Res. 2016, 5, 250–258. [Google Scholar] [CrossRef]
- Axelrad, J.E.; Lichtiger, S.; Yajnik, V. Inflammatory bowel disease and cancer: The role of inflammation, immunosuppression, and cancer treatment. World J. Gastroenterol. 2016, 22, 4794–4801. [Google Scholar] [CrossRef]
- Moura, F.A.; de Andrade, K.Q.; dos Santos, J.C.F.; Araújo, O.R.P.; Goulart, M.O.F. Antioxidant therapy for treatment of inflammatory bowel disease: Does it work? Redox Biol. 2015, 6, 617–639. [Google Scholar] [CrossRef] [PubMed]
- Dziąbowska-Grabias, K.; Sztanke, M.; Zając, P.; Celejewski, M.; Kurek, K.; Szkutnicki, S.; Korga, P.; Bulikowski, W.; Sztanke, K. Antioxidant Therapy in Inflammatory Bowel Diseases. Antioxidants 2021, 10, 412. [Google Scholar] [CrossRef]
- Mansouri, A.; Reiner, Ž.; Ruscica, M.; Tedeschi-Reiner, E.; Radbakhsh, S.; Bagheri Ekta, M.; Sahebkar, A. Antioxidant Effects of Statins by Modulating Nrf2 and Nrf2/HO-1 Signaling in Different Diseases. J. Clin. Med. 2022, 11, 1313. [Google Scholar] [CrossRef] [PubMed]
- Bohn, T.; Bonet, M.L.; Borel, P.; Keijer, J.; Landrier, J.F.; Milisav, I.; Ribot, J.; Riso, P.; Winklhofer-Roob, B.; Sharoni, Y.; et al. Mechanistic aspects of carotenoid health benefits—Where are we now? Nutr. Res. Rev. 2021, 34, 276–302. [Google Scholar] [CrossRef] [PubMed]
- Kaulmann, A.; Bohn, T. Bioactivity of Polyphenols: Preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases-Promises, Perspectives, and Pitfalls. Oxid. Med. Cell. Longev. 2016, 2016, 9346470. [Google Scholar] [CrossRef]
- Miranda-Díaz, A.G.; García-Sánchez, A.; Cardona-Muñoz, E.G. Foods with Potential Prooxidant and Antioxidant Effects Involved in Parkinson’s Disease. Oxid. Med. Cell. Longev. 2020, 2020, 6281454. [Google Scholar] [CrossRef]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef]
- Li, X.; Wei, X.; Sun, Y.; Du, J.; Li, X.; Xun, Z.; Li, Y.C. High-fat diet promotes experimental colitis by inducing oxidative stress in the colon. Am. J. Physiol. Gatroint. Liver Physiol. 2019, 317, G453–G462. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Wang, M.; Liu, Y.; Liang, Y.; Naruse, K.; Takahashi, K. Systematic Understanding of Pathophysiological Mechanisms of Oxidative Stress-Related Conditions—Diabetes Mellitus, Cardiovascular Diseases, and Ischemia–Reperfusion Injury. Front. Cardiov. Med. 2021, 8, 649785. [Google Scholar] [CrossRef]
- Sánchez-Terrón, G.; Morcuende, D.; Martínez, R.; Ruiz-Carrascal, J.; Estévez, M. Glucose boosts protein oxidation/nitration during simulated gastric digestion of myofibrillar proteins by creating a severe pro-oxidative environment. Food Chem. 2022, 397, 133805. [Google Scholar] [CrossRef] [PubMed]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Pavan Jukić, D.; Rotim, C.; Trebše, P.; Starc, A. Prooxidant activities of antioxidants and their impact on health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Goupy, P.; Dufour, C.; Loonis, M.; Dangles, O. Quantitative Kinetic Analysis of Hydrogen Transfer Reactions from Dietary Polyphenols to the DPPH Radical. J. Agric. Food Chem. 2003, 51, 615–622. [Google Scholar] [CrossRef]
- Sun, T.; Tanumihardjo, S.A. An Integrated Approach to Evaluate Food Antioxidant Capacity. J. Food Sci. 2007, 72, R159–R165. [Google Scholar] [CrossRef]
- Iddir, M.; Vahid, F.; Merten, D.; Larondelle, Y.; Bohn, T. Influence of Proteins on the Absorption of Lipophilic Vitamins, Carotenoids and Curcumin—A Review. Mol. Nutr. Food Res. 2022, 66, 2200076. [Google Scholar] [CrossRef]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef]
- Diep, T.T.; Yoo, M.J.Y.; Rush, E. Effect of In Vitro Gastrointestinal Digestion on Amino Acids, Polyphenols and Antioxidant Capacity of Tamarillo Yoghurts. Int. J. Mol. Sci. 2022, 23, 2526. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M. Extractable and Non-Extractable Antioxidants. Molecules 2019, 24, 1933. [Google Scholar] [CrossRef]
- Vahid, F.; Brito, A.; Le Coroller, G.; Vaillant, M.; Samouda, H.; Bohn, T.; on behalf of ORISCAV Working Group. Dietary Intake of Adult Residents in Luxembourg Taking Part in Two Cross-Sectional Studies—ORISCAV-LUX (2007–2008) and ORISCAV-LUX 2 (2016–2017). Nutrients 2021, 13, 4382. [Google Scholar] [CrossRef]
- Vahid, F.; Hoge, A.; Hébert, J.R.; Bohn, T. Association of diet quality indices with serum and metabolic biomarkers in participants of the ORISCAV-LUX-2 study. Eur. J. Nutr. 2023. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assuncao, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carriere, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef] [PubMed]
- Amarowicz, R.; Pegg, R.B. Chapter One—Natural antioxidants of plant origin. In Advances in Food and Nutrition Research; Ferreira, I.C.F.R., Barros, L., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 90, pp. 1–81. [Google Scholar]
- Nenadis, N.; Tsimidou, M.Z. Assessing the activity of natural food antioxidants. In Oxidation in Foods and Beverages and Antioxidant Applications; Decker, E.A., Ed.; Woodhead Publishing: Sawston, UK, 2010; pp. 332–367. [Google Scholar] [CrossRef]
- Gutteridge, J.M.; Halliwell, B. The measurement and mechanism of lipid peroxidation in biological systems. Trends Biochem. Sci. 1990, 15, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Devasagayam, T.; Boloor, K.; Ramasarma, T. Methods for estimating lipid peroxidation: An analysis of merits and demerits. Indian J. Biochem. Biophys. 2003, 40, 300–308. [Google Scholar]
- Kedare, S.B.; Singh, R.P. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef]
- Hernández-Ruiz, Á.; García-Villanova, B.; Guerra-Hernández, E.; Amiano, P.; Ruiz-Canela, M.; Molina-Montes, E. A Review of A Priori Defined Oxidative Balance Scores Relative to Their Components and Impact on Health Outcomes. Nutrients 2019, 11, 774. [Google Scholar] [CrossRef]
- Kaulmann, A.; Jonville, M.-C.; Schneider, Y.-J.; Hoffmann, L.; Bohn, T. Carotenoids, polyphenols and micronutrient profiles of Brassica oleraceae and plum varieties and their contribution to measures of total antioxidant capacity. Food Chem. 2014, 155, 240–250. [Google Scholar] [CrossRef]
- Melo, P.S.; Arrivetti, L.d.O.R.; de Alencar, S.M.; Skibsted, L.H. Antioxidative and prooxidative effects in food lipids and synergism with α-tocopherol of açaí seed extracts and grape rachis extracts. Food Chem. 2016, 213, 440–449. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; He, Y.; Jia, L.; Yang, C.S.; Reiter, R.J.; Zhang, J. Antioxidant and Pro-Oxidant Activities of Melatonin in the Presence of Copper and Polyphenols In Vitro and In Vivo. Cells 2019, 8, 903. [Google Scholar] [CrossRef]
- Cömert, E.D.; Gökmen, V. Effect of food combinations and their co-digestion on total antioxidant capacity under simulated gastrointestinal conditions. Curr. Res. Food Sci. 2022, 5, 414–422. [Google Scholar] [CrossRef]
- Mancuso, C.; Pani, G.; Calabrese, V. Bilirubin: An endogenous scavenger of nitric oxide and reactive nitrogen species. Redox Rep. 2006, 11, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Luckring, E.J.; Parker, P.D.; Hani, H.; Grace, M.H.; Lila, M.A.; Pierce, J.G.; Adin, C.A. In Vitro Evaluation of a Novel Synthetic Bilirubin Analog as an Antioxidant and Cytoprotective Agent for Pancreatic Islet Transplantation. Cell Transplant. 2020, 29. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, T.W.; Saleh, M.; Higginson, D.S.; Paul, B.D.; Juluri, K.R.; Snyder, S.H. Bilirubin and glutathione have complementary antioxidant and cytoprotective roles. Proc. Natl. Acad. Sci. USA 2009, 106, 5171–5176. [Google Scholar] [CrossRef] [PubMed]
- Asad, S.F.; Singh, S.; Ahmad, A.; Khan, N.U.; Hadi, S.M. Prooxidant and antioxidant activities of bilirubin and its metabolic precursor biliverdin: A structure–activity study. Chem.-Biol. Interact. 2001, 137, 59–74. [Google Scholar] [CrossRef]
- Bouayed, J.; Bohn, T. Exogenous antioxidants—Double-edged swords in cellular redox state: Health beneficial effects at physiologic doses versus deleterious effects at high doses. Oxid. Med. Cell. Longev. 2010, 3, 228–237. [Google Scholar] [CrossRef]
- Öztürk-Kerimoğlu, B.; Nacak, B.; Özyurt, V.H.; Serdaroğlu, M. Protein oxidation and in vitro digestibility of heat-treated fermented sausages: How do they change with the effect of lipid formulation during processing? J. Food Biochem. 2019, 43, e13007. [Google Scholar] [CrossRef]
- Morrissey, P.A.; Tichivangana, J.Z. The antioxidant activities of nitrite and nitrosylmyoglobin in cooked meats. Meat Sci. 1985, 14, 175–190. [Google Scholar] [CrossRef]
- Van Hecke, T.; Vanden Bussche, J.; Vanhaecke, L.; Vossen, E.; Van Camp, J.; De Smet, S. Nitrite Curing of Chicken, Pork, and Beef Inhibits Oxidation but Does Not Affect N-Nitroso Compound (NOC)-Specific DNA Adduct Formation during in Vitro Digestion. J. Agric. Food Chem. 2014, 62, 1980–1988. [Google Scholar] [CrossRef]
- Kanner, J. Oxidative processes in meat and meat products: Quality implications. Meat Sci. 1994, 36, 169–189. [Google Scholar] [CrossRef]
- Honikel, K.-O. The use and control of nitrate and nitrite for the processing of meat products. Meat Sci. 2008, 78, 68–76. [Google Scholar] [CrossRef]
- Özbay, S.; Şireli, U.T. The effect of ascorbic acid, storage period and packaging material on the formation of volatile N-nitrosamine in sausages. J. Food Sci. Technol. 2022, 59, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Noisa, P.; Yongsawatdigul, J. Chemical and Cellular Antioxidant Activities of In Vitro Digesta of Tilapia Protein and Its Hydrolysates. Foods 2020, 9, 833. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiov. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Papac-Milicevic, N.; Busch, C.J.L.; Binder, C.J. Chapter One—Malondialdehyde Epitopes as Targets of Immunity and the Implications for Atherosclerosis. In Advances in Immunology; Alt, F.W., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 131, pp. 1–59. [Google Scholar]
- Nieva-Echevarría, B.; Goicoechea, E.; Guillén, M.D. Food lipid oxidation under gastrointestinal digestion conditions: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef] [PubMed]
- Katz, D.L.; Doughty, K.; Ali, A. Cocoa and chocolate in human health and disease. Antioxid. Redox Signal. 2011, 15, 2779–2811. [Google Scholar] [CrossRef]
- Phillips, K.M.; Carlsen, M.H.; Blomhoff, R. Total Antioxidant Content of Alternatives to Refined Sugar. J. Am. Diet. Assoc. 2009, 109, 64–71. [Google Scholar] [CrossRef]
- Benzie, I.F.; Choi, S.W. Antioxidants in food: Content, measurement, significance, action, cautions, caveats, and research needs. Adv. Food Nutr. Res. 2014, 71, 1–53. [Google Scholar] [CrossRef]
- Dong, J.W.; Cai, L.; Xing, Y.; Yu, J.; Ding, Z.T. Re-evaluation of ABTS*+ Assay for Total Antioxidant Capacity of Natural Products. Nat. Prod. Commun. 2015, 10, 2169–2172. [Google Scholar] [CrossRef]
- Amarowicz, R. Antioxidant activity of Maillard reaction products. Eur. J. Lipid Sci. Technol. 2009, 111, 109–111. [Google Scholar] [CrossRef]
- Walker, R.B.; Everette, J.D. Comparative Reaction Rates of Various Antioxidants with ABTS Radical Cation. J. Agric. Food Chem. 2009, 57, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.W.; Hedges, N. Measuring lipid oxidation. In Understanding and Measuring the Shelf-Life of Food; Steele, R., Ed.; Woodhead Publishing: Sawston, UK, 2004; pp. 289–316. [Google Scholar] [CrossRef]
- Ganhão, R.; Estévez, M.; Morcuende, D. Suitability of the TBA method for assessing lipid oxidation in a meat system with added phenolic-rich materials. Food Chem. 2011, 126, 772–778. [Google Scholar] [CrossRef]
- Prasad, K.; Dhar, I. Oxidative stress as a mechanism of added sugar-induced cardiovascular disease. Int. J. Angiol. 2014, 23, 217–226. [Google Scholar] [PubMed]
- Martini, S.; Conte, A.; Bottazzi, S.; Tagliazucchi, D. Mediterranean diet vegetable foods protect meat lipids from oxidation during in vitro gastro-intestinal digestion. Int. J. Food Sci. Nutr. 2020, 71, 424–439. [Google Scholar] [CrossRef] [PubMed]
- Van Hecke, T.; Wouters, A.; Rombouts, C.; Izzati, T.; Berardo, A.; Vossen, E.; Claeys, E.; Van Camp, J.; Raes, K.; Vanhaecke, L.; et al. Reducing Compounds Equivocally Influence Oxidation during Digestion of a High-Fat Beef Product, which Promotes Cytotoxicity in Colorectal Carcinoma Cell Lines. J. Agric. Food Chem. 2016, 64, 1600–1609. [Google Scholar] [CrossRef]
- Yashin, A.; Yashin, Y.; Wang, J.Y.; Nemzer, B. Antioxidant and Antiradical Activity of Coffee. Antioxidants 2013, 2, 230–245. [Google Scholar] [CrossRef]
- Farias-Pereira, R.; Park, C.S.; Park, Y. Mechanisms of action of coffee bioactive components on lipid metabolism. Food Sci. Biotechnol. 2019, 28, 1287–1296. [Google Scholar] [CrossRef]
- Gómez-Ruiz, J.Á.; Leake, D.S.; Ames, J.M. In Vitro Antioxidant Activity of Coffee Compounds and Their Metabolites. J. Agric. Food Chem. 2007, 55, 6962–6969. [Google Scholar] [CrossRef]
- Sánchez-González, I.; Jiménez-Escrig, A.; Saura-Calixto, F. In vitro antioxidant activity of coffees brewed using different procedures (Italian, espresso and filter). Food Chem. 2005, 90, 133–139. [Google Scholar] [CrossRef]
- Kim, J.-H.; Ahn, D.U.; Eun, J.B.; Moon, S.H. Antioxidant Effect of Extracts from the Coffee Residue in Raw and Cooked Meat. Antioxidants 2016, 5, 21. [Google Scholar] [CrossRef]
- Franke, A.A.; Cooney, R.V.; Henning, S.M.; Custer, L.J. Bioavailability and antioxidant effects of orange juice components in humans. J. Agric. Food Chem. 2005, 53, 5170–5178. [Google Scholar] [CrossRef]
- Foroudi, S.; Potter, A.S.; Stamatikos, A.; Patil, B.S.; Deyhim, F. Drinking orange juice increases total antioxidant status and decreases lipid peroxidation in adults. J. Med. Food 2014, 17, 612–617. [Google Scholar] [CrossRef]
- Simons, L.A.; Von Konigsmark, M.; Balasubramaniam, S. What dose of vitamin E is required to reduce susceptibility of LDL to oxidation? Aust. N. Z. J. Med. 1996, 26, 496–503. [Google Scholar] [CrossRef]
- Liu, D.; Shi, J.; Colina Ibarra, A.; Kakuda, Y.; Jun Xue, S. The scavenging capacity and synergistic effects of lycopene, vitamin E, vitamin C, and β-carotene mixtures on the DPPH free radical. LWT-Food Sci. Technol. 2008, 41, 1344–1349. [Google Scholar] [CrossRef]
- Ak, T.; Gülçin, İ. Antioxidant and radical scavenging properties of curcumin. Chem. Biol. Interact. 2008, 174, 27–37. [Google Scholar] [CrossRef]
- Müller, M.; Canfora, E.E.; Blaak, E.E. Gastrointestinal Transit Time, Glucose Homeostasis and Metabolic Health: Modulation by Dietary Fibers. Nutrients 2018, 10, 275. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Item | Quantity Used per Simulated Digestion | Nutrients (per 100 mL for Liquids and 100 g for Solids) | Ingredients as Listed on Food Labels |
|---|---|---|---|
| Tomato juice | 2 mL | Fats (<0.5 g), carbohydrates (3 g), starch (0.6 g), proteins (0.8 g), fiber (0.6 g), total energy (16 kcal) | Tomato juice (99.5%), salt (0.5%) |
| Orange juice | 2 mL | Fats (0 g), carbohydrates (8.7 g), proteins (0.7 g), vitamin C (12 mg), total energy (42 kcal) | Juice 100% |
| Vitamin C | 2.5 mg | - | L-ascorbic acid (99%) |
| Vitamin E | 1.5 mg | - | DL-alpha-tocopherol (>97%) |
| Coffee | 2 mL | ≈8 mg caffeine *, ≈100 mg of total polyphenols ** | Coffee, 100% Arabica |
| Curcumin | 12.5 mg | - | Curcumin (98%) |
| Sausage | 2 g | Fats (29 g), saturated fats (11 g), carbohydrates (7.5 g), proteins (9 g), salt (1.8 g), total energy (329 kcal) | Pork liver 25%, pork liver 22%, pork fat, water, pork rind, potato starch, dextrose, salt, cream, sugar, onions, preservatives (potassium acetate, sodium nitrite), spices, emulsifiers (citric acid esters from mono- and diglycerides from fatty acids), antioxidants (ascorbic acid, sodium ascorbate), thickener (xanthan), spice extracts, hemoglobin, aroma |
| Soda | 2 mL | Fats (0 g), carbohydrates/of which sugars (6.6 g/6.6 g), proteins (0 g), total energy (28 kcal) | Sparkling water, sugar, acidifiers: citric acid, malic acid, acidity corrector; sodium gluconate, natural lemon-green lemon aroma, sweetener: steviol glycoside |
| White chocolate | 2 g | Fats (25 g), saturated fats (22 g), carbohydrates/of which sugars (55/55 g), proteins (5.7 g), total energy (561 kcal) | Sugar, cocoa butter, whole milk powder, skim milk powder, emulsifier (soya lecithin), flavoring |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vahid, F.; Wagener, L.; Leners, B.; Bohn, T. Pro- and Antioxidant Effect of Food Items and Matrices during Simulated In Vitro Digestion. Foods 2023, 12, 1719. https://doi.org/10.3390/foods12081719
Vahid F, Wagener L, Leners B, Bohn T. Pro- and Antioxidant Effect of Food Items and Matrices during Simulated In Vitro Digestion. Foods. 2023; 12(8):1719. https://doi.org/10.3390/foods12081719
Chicago/Turabian StyleVahid, Farhad, Lisi Wagener, Bernadette Leners, and Torsten Bohn. 2023. "Pro- and Antioxidant Effect of Food Items and Matrices during Simulated In Vitro Digestion" Foods 12, no. 8: 1719. https://doi.org/10.3390/foods12081719