
Post-Harvest Red- and Far-Red-Light Irradiation and Low Temperature Induce the Accumulation of Carotenoids, Capsaicinoids, and Ascorbic Acid in Capsicum annuum L. Green Pepper Fruit
 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Pigment and Water Contents
2.3. HPLC Analysis of Carotenoids
2.4. HPLC Analysis of Ascorbate
2.5. HPLC Analysis of Capsaicinoids
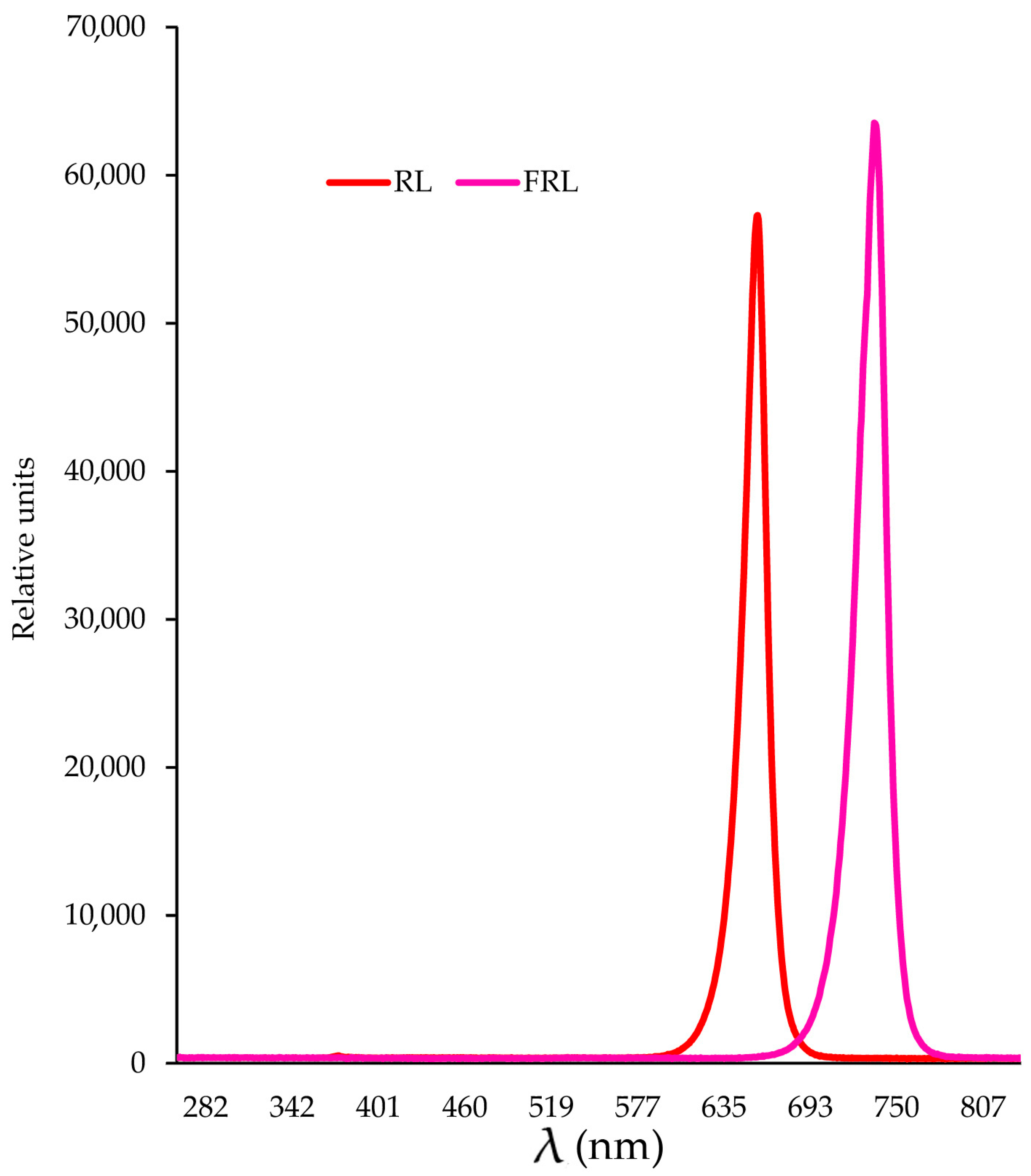
2.6. Determination of Photochemical Activity
2.7. RNA Extraction and RT-PCR
2.8. Statistical Data Processing
3. Results
3.1. Carotenoid, Pigment, and Water Contents
3.2. Primary Photochemical Processes of Photosynthesis
3.3. Ascorbate Content
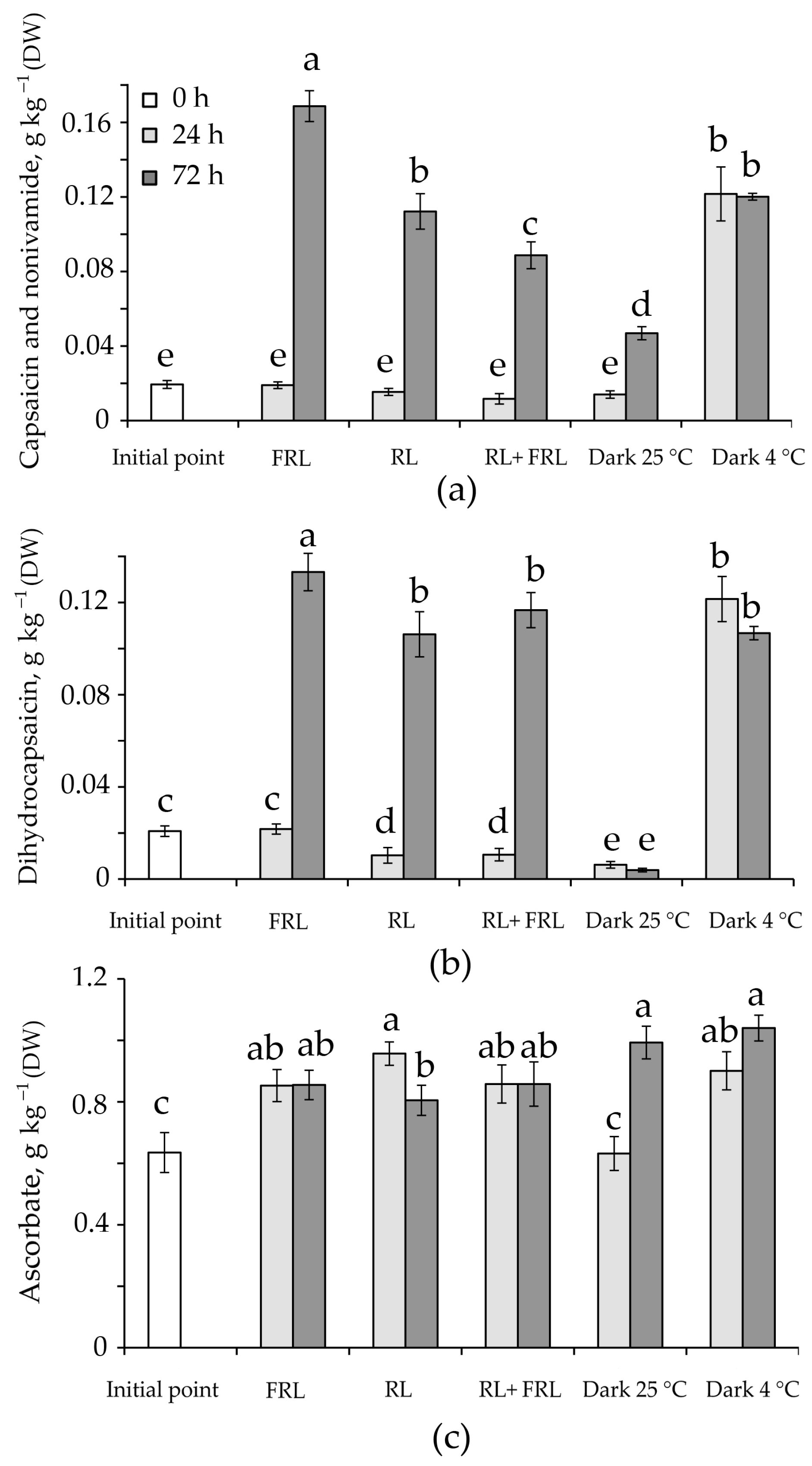
3.4. Content of Alkaloids
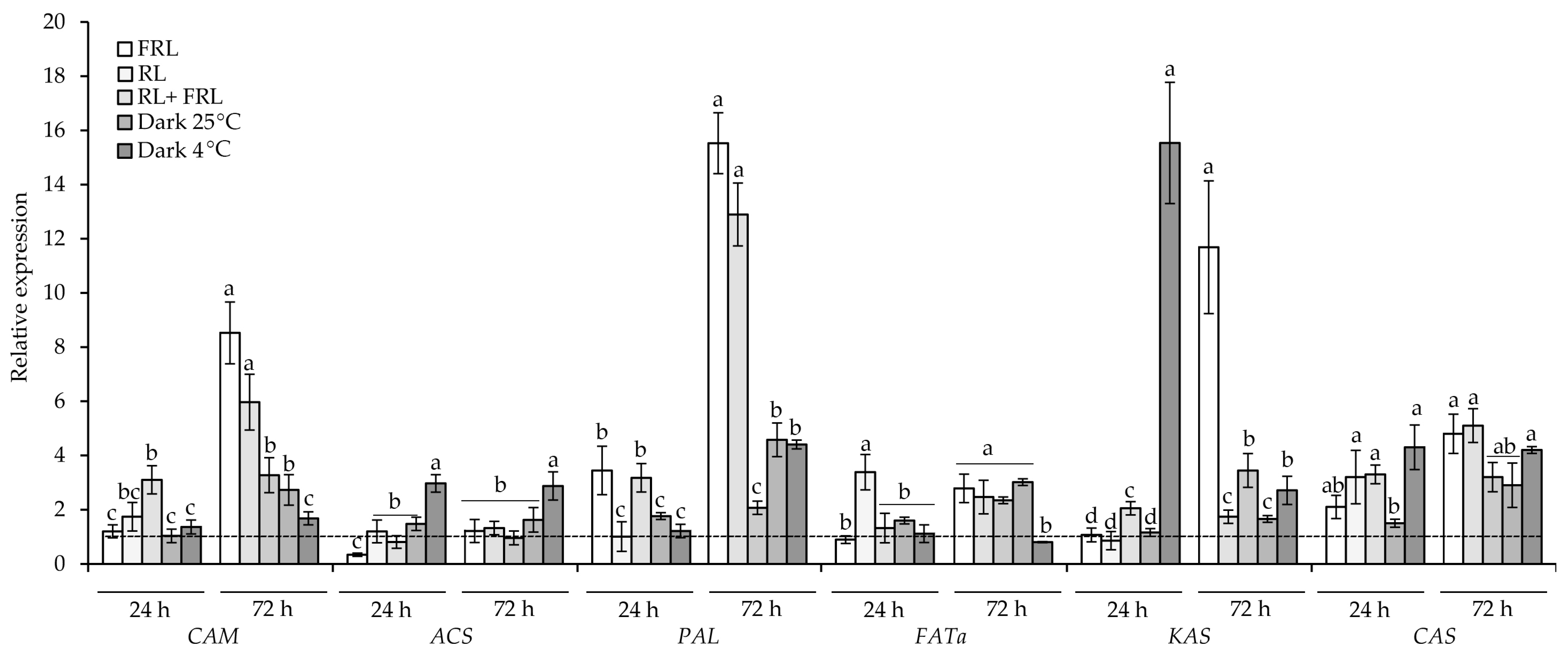
3.5. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Müller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical Review: Vegetables and Fruit in the Prevention of Chronic Diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Q.; Nguyen, P.; Lin, C. Cryptochrome-Mediated Light Responses in Plants. Enzymes 2014, 35, 167–189. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Kim, J.S. Anti-Carcinogenic Glucosinolates in Cruciferous Vegetables and Their Antagonistic Effects on Prevention of Cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef] [PubMed]
- Castillejo, N.; Martínez-Zamora, L.; Artés-Hernández, F. Postharvest UV Radiation Enhanced Biosynthesis of Flavonoids and Carotenes in Bell Peppers. Postharvest Biol. Technol. 2022, 184, 111774. [Google Scholar] [CrossRef]
- Arias, R.; Lee, T.-C.; Logendra, L.; Janes, H. Correlation of Lycopene Measured by HPLC with the L*, A*, B* Color Readings of a Hydroponic Tomato and the Relationship of Maturity with Color and Lycopene Content. J. Agric. Food Chem. 2000, 48, 1697–1702. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.; Hwang, I.K. Composition of Main Carotenoids in Korean Red Pepper (Capsicum annuum, L.) and Changes of Pigment Stability During the Drying and Storage Process. J. Food Sci. 2004, 69, FCT39–FCT44. [Google Scholar] [CrossRef]
- Guerra, M.; Magdaleno, R.; Casquero, P.A. Effect of Site and Storage Conditions on Quality of Industrial Fresh Pepper. Sci. Hortic. 2011, 130, 141–145. [Google Scholar] [CrossRef]
- Maroga, G.M.; Soundy, P.; Sivakumar, D. Different Postharvest Responses of Fresh-Cut Sweet Peppers Related to Quality and Antioxidant and Phenylalanine Ammonia Lyase Activities during Exposure to Light-Emitting Diode Treatments. Foods 2019, 8, 359. [Google Scholar] [CrossRef]
- Saini, R.K.; Zamany, A.J.; Keum, Y.-S. Ripening Improves the Content of Carotenoid, α-Tocopherol, and Polyunsaturated Fatty Acids in Tomato (Solanum lycopersicum L.) Fruits. 3 Biotech 2017, 7, 43. [Google Scholar] [CrossRef]
- Kapoor, L.; Simkin, A.J.; George Priya Doss, C.; Siva, R. Fruit Ripening: Dynamics and Integrated Analysis of Carotenoids and Anthocyanins. BMC Plant Biol. 2022, 22, 27. [Google Scholar] [CrossRef]
- Kim, D.; Moon, T.; Kwon, S.; Hwang, I.; Son, J.E. Supplemental Inter-Lighting with Additional Far-Red to Red and Blue Light Increases the Growth and Yield of Greenhouse Sweet Peppers (Capsicum annuum L.) in Winter. Hortic. Environ. Biotechnol. 2022, 64, 83–95. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Song, S.; Su, W.; Hao, Y.; Liu, H. Supplementary Red Light Results in the Earlier Ripening of Tomato Fruit Depending on Ethylene Production. Environ. Exp. Bot. 2020, 175, 104044. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, X.; Gao, X.; Wu, W.; Zhou, B. Light Induced Regulation Pathway of Anthocyanin Biosynthesis in Plants. Int. J. Mol. Sci. 2021, 22, 11116. [Google Scholar] [CrossRef]
- Voitsekhovskaja, O.V. Phytochromes and Other (Photo)Receptors of Information in Plants. Russ. J. Plant Physiol. 2019, 66, 351–364. [Google Scholar] [CrossRef]
- Jung, J.-H.; Domijan, M.; Klose, C.; Biswas, S.; Ezer, D.; Gao, M.; Khattak, A.K.; Box, M.S.; Charoensawan, V.; Cortijo, S.; et al. Phytochromes Function as Thermosensors in Arabidopsis. Science 2016, 354, 886–889. [Google Scholar] [CrossRef]
- Legris, M.; Klose, C.; Burgie, E.S.; Rojas, C.C.R.; Neme, M.; Hiltbrunner, A.; Wigge, P.A.; Schäfer, E.; Vierstra, R.D.; Casal, J.J. Phytochrome B Integrates Light and Temperature Signals in Arabidopsis. Science 2016, 354, 897–900. [Google Scholar] [CrossRef]
- Pashkovskiy, P.; Kreslavskii, V.; Khudyakova, A.; Kosobryukhov, A.; Kuznetsov, V.; Allakhverdiev, S. Influence of Phytochromes on MicroRNA Expression, Phenotype, and Photosynthetic Activity in A. Thaliana Phy Mutants under Light with Different Spectral Composition. Photosynthetica 2022, 61, 135–144. [Google Scholar] [CrossRef]
- Quail, P.H. Phytochromes. Curr. Biol. 2010, 20, R504–R507. [Google Scholar] [CrossRef]
- Jenkins, G.I. Signal Transduction in Responses to UV-B Radiation. Annu. Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar] [CrossRef]
- Bae, G.; Choi, G. Decoding of Light Signals by Plant Phytochromes and Their Interacting Proteins. Annu. Rev. Plant Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef]
- Lupi, A.C.D.; Lira, B.S.; Gramegna, G.; Trench, B.; Alves, F.R.R.; Demarco, D.; Peres, L.E.P.; Purgatto, E.; Freschi, L.; Rossi, M. Solanum Lycopersicum GOLDEN 2-LIKE 2 Transcription Factor Affects Fruit Quality in a Light- and Auxin-Dependent Manner. PLoS ONE 2019, 14, e0212224. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Hu, B.R. Phytohormones Act as Information-Bridges between Light and Temperature Signaling Transduction Pathways. Adv. Mater. Res. 2014, 1010–1012, 437–441. [Google Scholar] [CrossRef]
- Gavassi, M.A.; Monteiro, C.C.; Campos, M.L.; Melo, H.C.; Carvalho, R.F. Phytochromes Are Key Regulators of Abiotic Stress Responses in Tomato. Sci. Hortic. 2017, 222, 126–135. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Los, D.A.; Schmitt, F.-J.; Zharmukhamedov, S.K.; Kuznetsov, V.V.; Allakhverdiev, S.I. The Impact of the Phytochromes on Photosynthetic Processes. Biochim. Biophys. Acta BBA-Bioenerg. 2018, 1859, 400–408. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M. Photosynthesis, Morphology, Yield, and Phytochemical Accumulation in Basil Plants Influenced by Substituting Green Light for Partial Red and/or Blue Light. HortScience 2019, 54, 1769–1776. [Google Scholar] [CrossRef]
- Moreira-Rodríguez, M.; Nair, V.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. UVA, UVB Light, and Methyl Jasmonate, Alone or Combined, Redirect the Biosynthesis of Glucosinolates, Phenolics, Carotenoids, and Chlorophylls in Broccoli Sprouts. Int. J. Mol. Sci. 2017, 18, 2330. [Google Scholar] [CrossRef]
- Pérez-Ambrocio, A.; Guerrero-Beltrán, J.A.; Aparicio-Fernández, X.; Ávila-Sosa, R.; Hernández-Carranza, P.; Cid-Pérez, S.; Ochoa-Velasco, C.E. Effect of Blue and Ultraviolet-C Light Irradiation on Bioactive Compounds and Antioxidant Capacity of Habanero Pepper (Capsicum Chinense) during Refrigeration Storage. Postharvest Biol. Technol. 2018, 135, 19–26. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of Abiotic Stress Signals on Secondary Metabolites in Plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Christie, P.J.; Alfenito, M.R.; Walbot, V. Impact of Low-Temperature Stress on General Phenylpropanoid and Anthocyanin Pathways: Enhancement of Transcript Abundance and Anthocyanin Pigmentation in Maize Seedlings. Planta 1994, 194, 541–549. [Google Scholar] [CrossRef]
- Griffith, M.; Yaish, M.W.F. Antifreeze Proteins in Overwintering Plants: A Tale of Two Activities. Trends Plant Sci. 2004, 9, 399–405. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Plant Cell Membranes; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. A Guide to Carotenoid Analysis in Foods; ILSI Press: Washington, DC, USA, 2001. [Google Scholar]
- Ashikhmin, A.; Makhneva, Z.; Bolshakov, M.; Moskalenko, A. Incorporation of Spheroidene and Spheroidenone into Light-Harvesting Complexes from Purple Sulfur Bacteria. J. Photochem. Photobiol. B Biol. 2017, 170, 99–107. [Google Scholar] [CrossRef]
- Nováková, L.; Solich, P.; Solichová, D. HPLC Methods for Simultaneous Determination of Ascorbic and Dehydroascorbic Acids. TrAC Trends Anal. Chem. 2008, 27, 942–958. [Google Scholar] [CrossRef]
- Arce-Rodríguez, M.L.; Ochoa-Alejo, N. Silencing AT3 Gene Reduces the Expression of PAmt, BCAT, Kas, and Acl Genes Involved in Capsaicinoid Biosynthesis in Chili Pepper Fruits. Biol. Plant 2015, 59, 477–484. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Strokina, V.V.; Pashkovskiy, P.P.; Balakhnina, T.I.; Voloshin, R.A.; Alwasel, S.; Kosobryukhov, A.A.; Allakhverdiev, S.I. Deficiencies in Phytochromes A and B and Cryptochrome 1 Affect the Resistance of the Photosynthetic Apparatus to High-Intensity Light in Solanum Lycopersicum. J. Photochem. Photobiol. B Biol. 2020, 210, 111976. [Google Scholar] [CrossRef]
- Liu, C.; Wan, H.; Yang, Y.; Ye, Q.; Zhou, G.; Wang, X.; Ahammed, G.J.; Cheng, Y. Post-Harvest LED Light Irradiation Affects Firmness, Bioactive Substances, and Amino Acid Compositions in Chili Pepper (Capsicum annum L.). Foods 2022, 11, 2712. [Google Scholar] [CrossRef]
- Hui-jun, G.A.O.; Jia-qi, M.; Ya-juan, Z.; Juan, X.U. Regulation of Carotenoids Biosynthesis in Horticultural Crops. Acta Hortic. Sin. 2015, 42, 1633. [Google Scholar]
- Tan, T.; Li, S.; Fan, Y.; Wang, Z.; Ali Raza, M.; Shafiq, I.; Wang, B.; Wu, X.; Yong, T.; Wang, X.; et al. Far-Red Light: A Regulator of Plant Morphology and Photosynthetic Capacity. Crop J. 2022, 10, 300–309. [Google Scholar] [CrossRef]
- Gao, L.-M.; Li, Y.-F.; Han, R. He-Ne Laser Preillumination Improves the Resistance of Tall Fescue (Festuca Arundinacea Schreb.) Seedlings to High Saline Conditions. Protoplasma 2015, 252, 1135–1148. [Google Scholar] [CrossRef]
- Yang, F.; Feng, L.; Liu, Q.; Wu, X.; Fan, Y.; Raza, M.A.; Cheng, Y.; Chen, J.; Wang, X.; Yong, T.; et al. Effect of Interactions between Light Intensity and Red-to- Far-Red Ratio on the Photosynthesis of Soybean Leaves under Shade Condition. Environ. Exp. Bot. 2018, 150, 79–87. [Google Scholar] [CrossRef]
- Kim, D.; Son, J.E. Adding Far-Red to Red, Blue Supplemental Light-Emitting Diode Interlighting Improved Sweet Pepper Yield but Attenuated Carotenoid Content. Front. Plant Sci. 2022, 13, 938199. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue Light Added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Marin, E.; Galano, A.; Martínez, A. Cis Carotenoids: Colorful Molecules and Free Radical Quenchers. J. Phys. Chem. B 2013, 117, 4050–4061. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Ortiz, G.; Huq, E.; Rodríguez-Concepción, M. Direct Regulation of Phytoene Synthase Gene Expression and Carotenoid Biosynthesis by Phytochrome-Interacting Factors. Proc. Natl. Acad. Sci. USA 2010, 107, 11626–11631. [Google Scholar] [CrossRef] [PubMed]
- Pashkovskiy, P.; Vereshchagin, M.; Kreslavski, V.; Ivanov, Y.; Kumachova, T.; Ryabchenko, A.; Voronkov, A.; Kosobryukhov, A.; Kuznetsov, V.; Allakhverdiev, S.I. Effect of Phytochrome Deficiency on Photosynthesis, Light-Related Genes Expression and Flavonoid Accumulation in Solanum Lycopersicum under Red and Blue Light. Cells 2022, 11, 3437. [Google Scholar] [CrossRef]
- Zhang, Z.-X.; Zhao, S.-N.; Liu, G.-F.; Huang, Z.-M.; Cao, Z.-M.; Cheng, S.-H.; Lin, S.-S. Discovery of Putative Capsaicin Biosynthetic Genes by RNA-Seq and Digital Gene Expression Analysis of Pepper. Sci. Rep. 2016, 6, 34121. [Google Scholar] [CrossRef]
- Nishida, I. Plastid Metabolic Pathways for Fatty Acid Metabolism. In Molecular Biology and Biotechnology of Plant Organelles: Chloroplasts and Mitochondria; Daniell, H., Chase, C., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 543–564. ISBN 978-1-4020-3166-3. [Google Scholar]
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.; Qin, Y. Plant Low-Temperature Stress: Signaling and Response. Agronomy 2022, 12, 702. [Google Scholar] [CrossRef]
- Wang, X.; Li, W.; Li, M.; Welti, R. Profiling Lipid Changes in Plant Response to Low Temperatures. Physiol. Plant. 2006, 126, 90–96. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Time, (h) | Chl (a + b), g kg−1 (DW) | Car + Xan, g kg−1, (DW) | Y(II) | Fv/Fm | Water Loss, % | |
|---|---|---|---|---|---|---|
| Initial point | 0 | 14.3 ± 0.7 a | 0.14 ± 0.02 e | 0.22 ± 0.01 b | 0.687 ± 0.021 ab | |
| FRL | 24 | 11.2 ± 0.4 b | 0.31 ± 0.01 c | 0.22 ± 0.01 b | 0.663 ± 0.026 ab | 6.81 ± 0.61 ab |
| RL | 10.2 ± 0.3 bc | 0.51 ± 0.02 a | 0.15 ± 0.02 bc | 0.545 ± 0.023 c | 5.95 ± 0.42 b | |
| RL + FRL | 11.8 ± 0.5 b | 0.29 ± 0.01 c | 0.20 ± 0.02 b | 0.454 ± 0.021 d | 8.21 ± 0.51 a | |
| Dark 25 °C | 11.1 ± 0.6 b | 0.21 ± 0.01 d | 0.28 ± 0.02 a | 0.650 ± 0.021 b | 4.66 ± 0.21 c | |
| Dark 4 °C | 10.4 ± 0.4 bc | 0.25 ± 0.01 bc | 0.23 ± 0.03 ab | 0.662 ± 0.022 ab | 1.54 ± 0.21 e | |
| FRL | 72 | 11.6 ± 0.3 b | 0.19 ± 0.01 de | 0.20 ± 0.02 b | 0.655 ± 0.021 ab | 3.63 ± 0.05 d |
| RL | 12.8 ± 0.5 b | 0.32 ± 0.01 c | 0.12 ± 0.01 c | 0.322 ± 0.023 e | 4.02 ± 0.12 d | |
| RL + FRL | 9.0 ± 0.3 c | 0.42 ± 0.02 b | 0.08 ± 0.01 d | 0.161 ± 0.023 e | 5.48 ± 0.04 bc | |
| Dark 25 °C | 11.0 ± 0.5 b | 0.28 ± 0.01 c | 0.25 ± 0.02 ab | 0.736 ± 0.021 a | 4.72 ± 0.12 c | |
| Dark 4 °C | 11.1 ± 0.5 b | 0.29 ± 0.01 c | 0.20 ± 0.01 b | 0.68 ± 0.024 ab | 1.52 ± 0.11 e |
| Time (h) | 0 | 24 | 72 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Initial Point | FRL | RL | RL + FRL | Dark 25 °C | Dark 4 °C | FRL | RL | RL + FRL | Dark 25 °C | Dark 4 °C | |
| Neoxanthin | 8.31 ± 0.91 b | 8.81 ± 1.31 b | 6.11 ± 0.81 c | 11.12 ± 1.1 ab | 10.50 ± 1.12 ab | 8.31 ± 1.25 b | 13.61 ± 1.61 a | 12.41 ± 1.21 a | 9.81 ± 1.04 b | 8.75 ± 1.64 b | 9.20 ± 0.94 b |
| Violoxanthin | 18.01 ± 1.51 c | 28.12 ± 2.24 b | 24.12 ± 2.85 b | 17.16 ± 0.91 c | 20.30 ± 2.84 bc | 28.61 ± 2.35 b | 39.31 ± 3.24 a | 28.43 ± 2.84 b | 22.42 ± 2.85 b | 26.85 ± 2.72 b | 33.11 ± 1.24 b |
| Neochrome | 0 | 0 | 0 | 0 | 0 | 0 | 0.60 ± 0.12 a | 0 | 0 | 0 | 0 |
| Capsanthin | 0.11 ± 0.03 c | 0.21 ± 0.03 b | 0.10 ± 0.03 c | 0 | 0.20 ± 0.03 b | 0.21 ± 0.03 b | 0.50 ± 0.07 a | 0.11 ± 0.02 c | 0.11 ± 0.02 c | 0.35 ± 0.03 b | 0.21 ± 0.03 b |
| Capsorubin | 0.31 ± 0.05 b | 0.11 ± 0.05 d | 0 | 0.21 ± 0.04 c | 0.30 ± 0.04 bc | 0.40 ± 0.06 b | 0.91 ± 0.09 a | 0.50 ± 0.07 b | 0 | 0.40 ± 0.05 b | 0.21 ± 0.04 c |
| Cis-mutatoxanthin | 0 | 0 | 5.00 ± 1.85 a | 0 | 0 | 0 | 0.60 ± 0.35 b | 0 | 5.00 ± 0.26 a | 0 | 0 |
| Anteroxanthin | 0.31 ± 0.05 b | 0 | 0 | 0.11 ± 0.05 c | 0 | 0.10 ± 0.05 c | 0.41 ± 0.03 a | 0.11 ± 0.05 c | 0 | 0 | 0.10 ± 0.04 c |
| Luteinepoxide | 0 | 0 | 11.51 ± 1.59 a | 2.00 ± 0.54 c | 0 | 0.20 ± 0.02 e | 3.62 ± 0.12 b | 1.10 ± 0.25 d | 10.11 ± 1.24 a | 0.73 ± 0.02 d | 0.31 ± 0.01 e |
| Lutein | 57.73 ± 1.81 a | 48.32 ± 2.85 b | 24.72 ± 2.75 e | 49.86 ± 2.44 b | 47.71 ± 2.54 b | 35.51 ± 2.85 d | 36.43 ± 2.52 d | 42.13 ± 2.62 c | 30.22 ± 2.42 e | 45.78 ± 2.14 bc | 43.97 ± 2.04 bc |
| Zeaxanthin | 0 | 0 | 13.21 ± 1.85 a | 2.02 ± 0.81 c | 0.31 ± 0.24 d | 0 | 1.61 ± 0.21 c | 0.60 ± 0.28 d | 9.43 ± 1.84 b | 1.62 ± 0.15 c | 0 |
| Neolutein | 0.50 ± 0.24 d | 9.51 ± 1.24 c | 7.42 ± 1.28 c | 9.91 ± 1.19 c | 18.11 ± 2.45 a | 13.50 ± 1.52 b | 0.40 ± 0.03 d | 9.51 ± 2.04 c | 10.90 ± 2.12 c | 13.42 ± 1.84 b | 9.52 ± 1.72 c |
| α-carotene | trace | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| β-carotene | 14.61 ± 1.08 a | 4.92 ± 1.22 c | 7.81 ± 1.32 b | 7.61 ± 2.01 b | 2.57 ± 0.58 d | 13.16 ± 2.12 a | 2.00 ± 0.24 d | 5.10 ± 1.24 c | 2.00 ± 0.95 d | 2.10 ± 0.93 d | 3.41 ± 0.95 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pashkovskiy, P.; Sleptsov, N.; Vereschagin, M.; Kreslavski, V.; Rudometova, N.; Sorokoumov, P.; Ashikhmin, A.; Bolshakov, M.; Kuznetsov, V. Post-Harvest Red- and Far-Red-Light Irradiation and Low Temperature Induce the Accumulation of Carotenoids, Capsaicinoids, and Ascorbic Acid in Capsicum annuum L. Green Pepper Fruit. Foods 2023, 12, 1715. https://doi.org/10.3390/foods12081715
Pashkovskiy P, Sleptsov N, Vereschagin M, Kreslavski V, Rudometova N, Sorokoumov P, Ashikhmin A, Bolshakov M, Kuznetsov V. Post-Harvest Red- and Far-Red-Light Irradiation and Low Temperature Induce the Accumulation of Carotenoids, Capsaicinoids, and Ascorbic Acid in Capsicum annuum L. Green Pepper Fruit. Foods. 2023; 12(8):1715. https://doi.org/10.3390/foods12081715
Chicago/Turabian StylePashkovskiy, Pavel, Nikolay Sleptsov, Mikhail Vereschagin, Vladimir Kreslavski, Natalia Rudometova, Pavel Sorokoumov, Aleksandr Ashikhmin, Maksim Bolshakov, and Vladimir Kuznetsov. 2023. "Post-Harvest Red- and Far-Red-Light Irradiation and Low Temperature Induce the Accumulation of Carotenoids, Capsaicinoids, and Ascorbic Acid in Capsicum annuum L. Green Pepper Fruit" Foods 12, no. 8: 1715. https://doi.org/10.3390/foods12081715