
Differences and Similarities in Lipid Composition, Nutritional Value, and Bioactive Potential of Four Edible Chlorella vulgaris Strains

 , ,
, ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Microalgae Strain and Culture Media
2.3. Lipid Extraction
2.4. Fatty Acid Analysis by GC-MS
2.5. C18- Liquid Chromatography–Mass Spectrometry (C18–LC–MS)
2.6. DPPH● and ABTS●+ Radicals Scavenging Assays
2.7. Anti-Inflammatory Activity
2.8. Statistical Analysis
3. Results
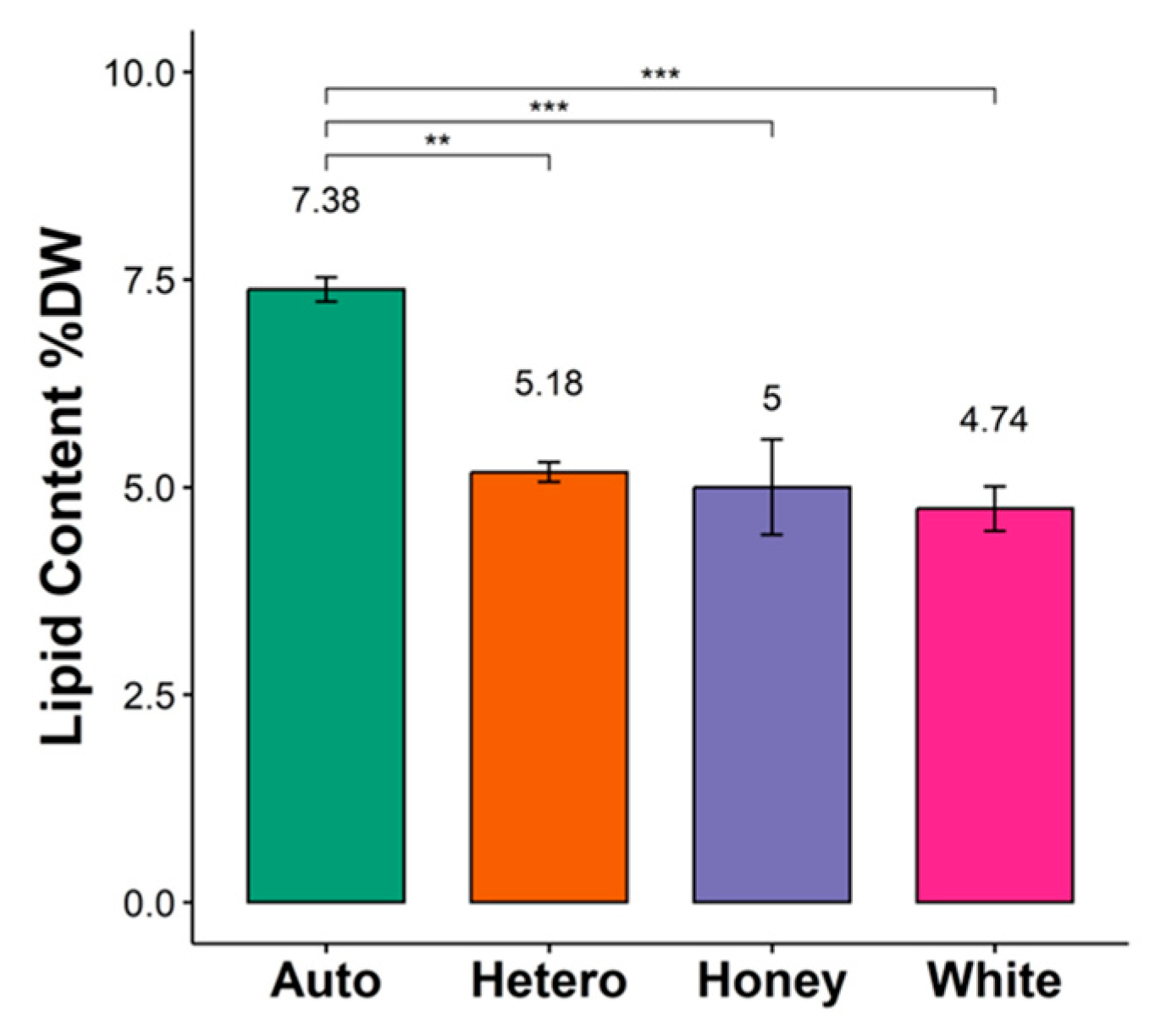
3.1. Lipid Content and Fatty Acid Profile of C. vulgaris Strains
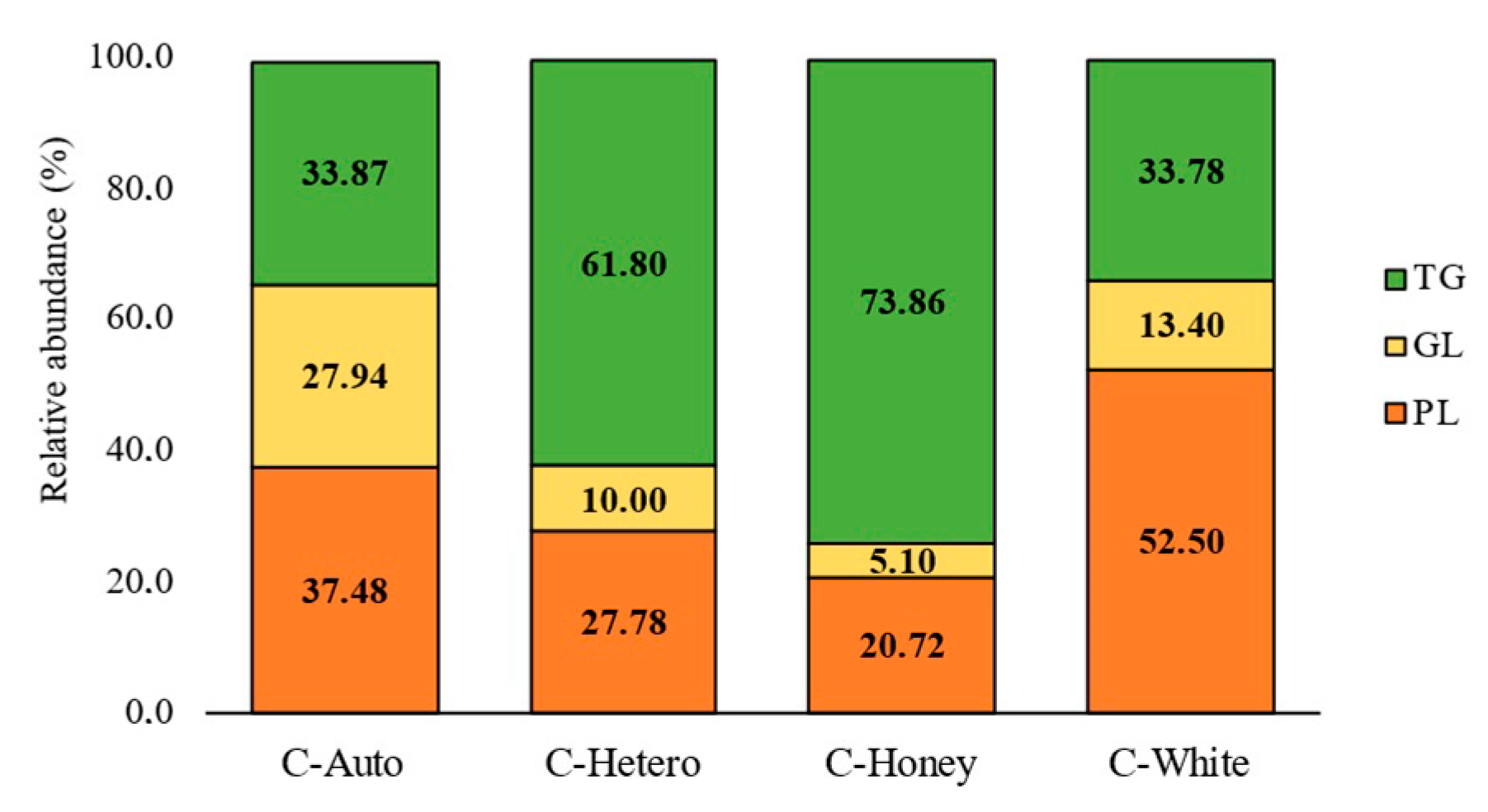
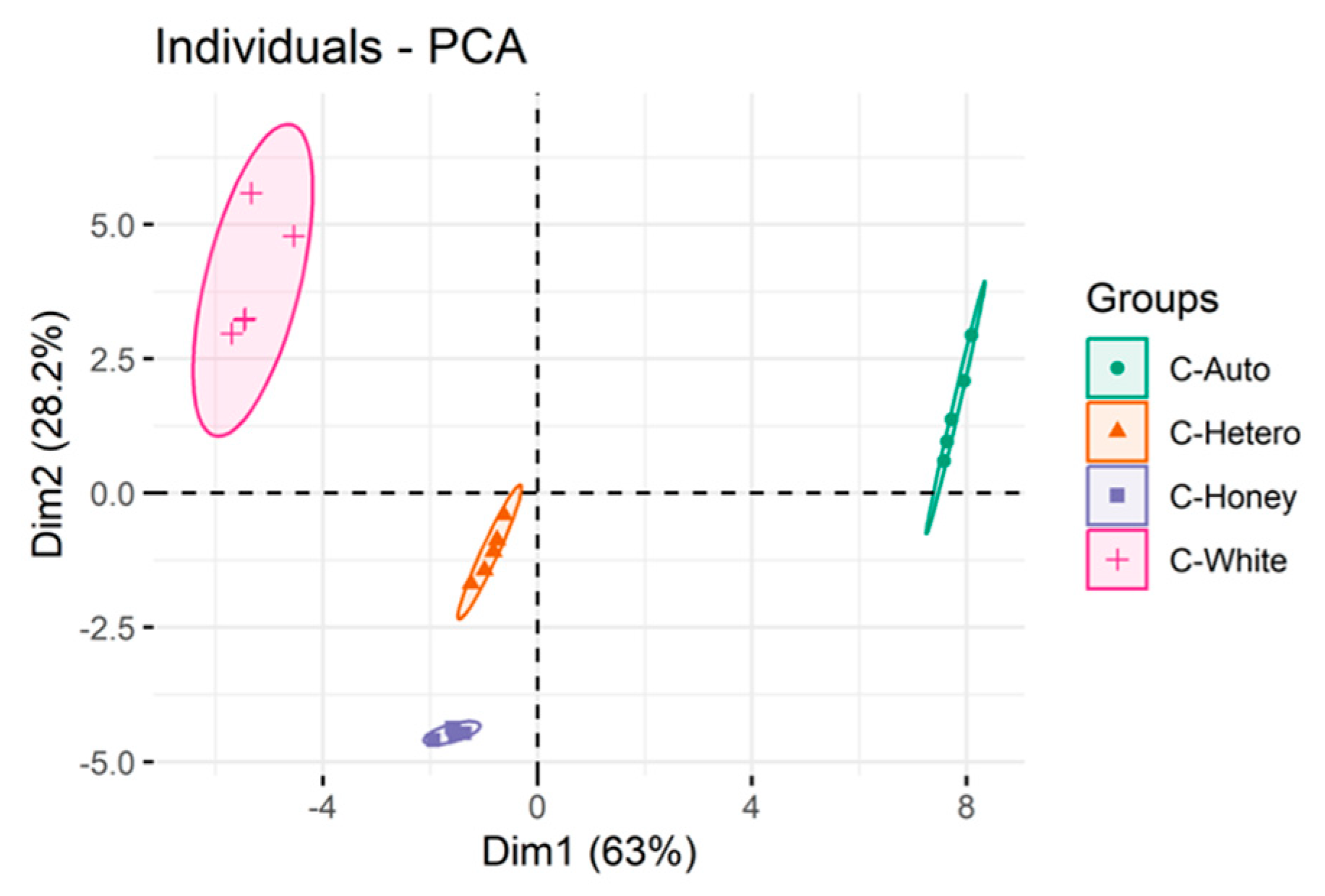
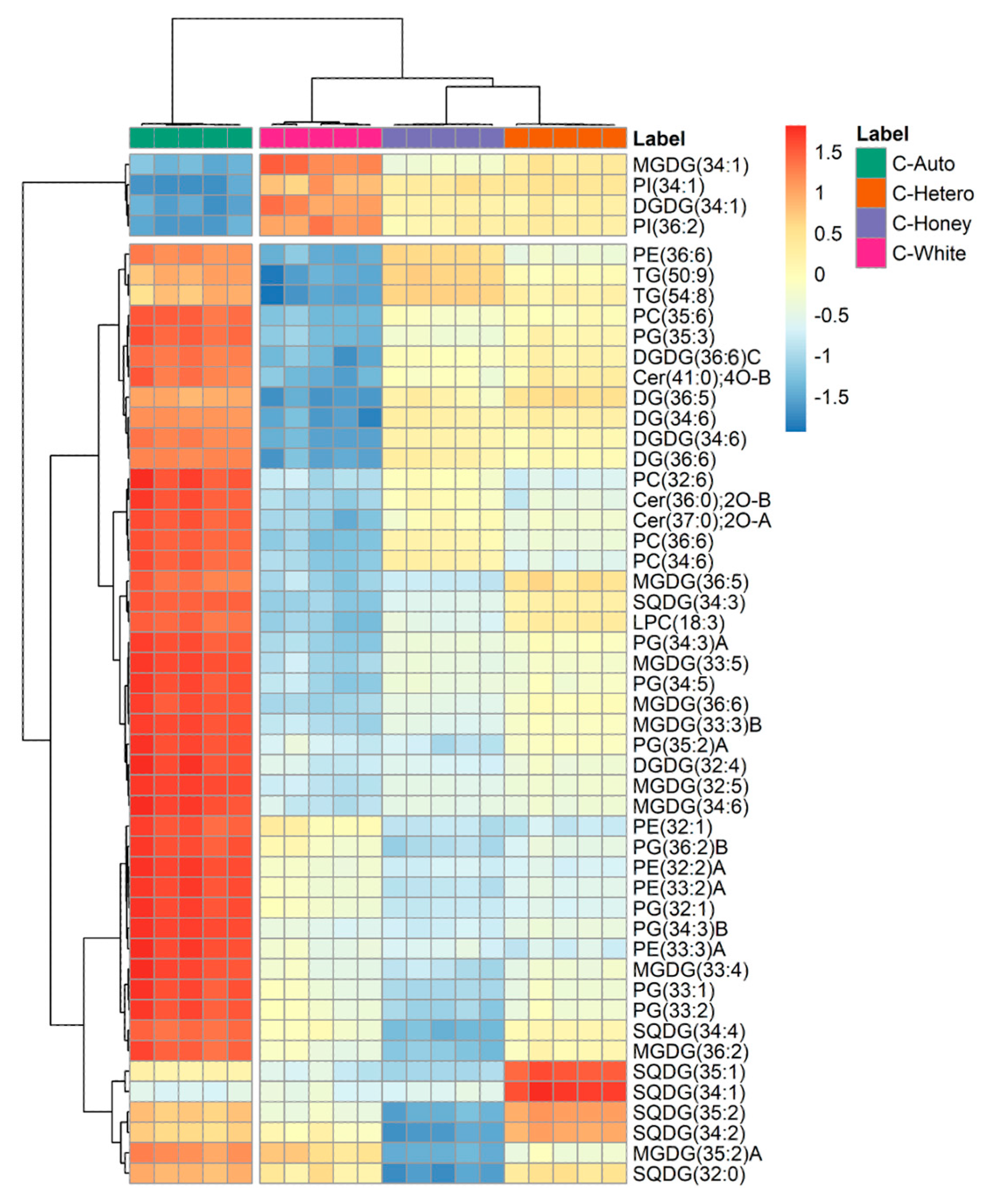
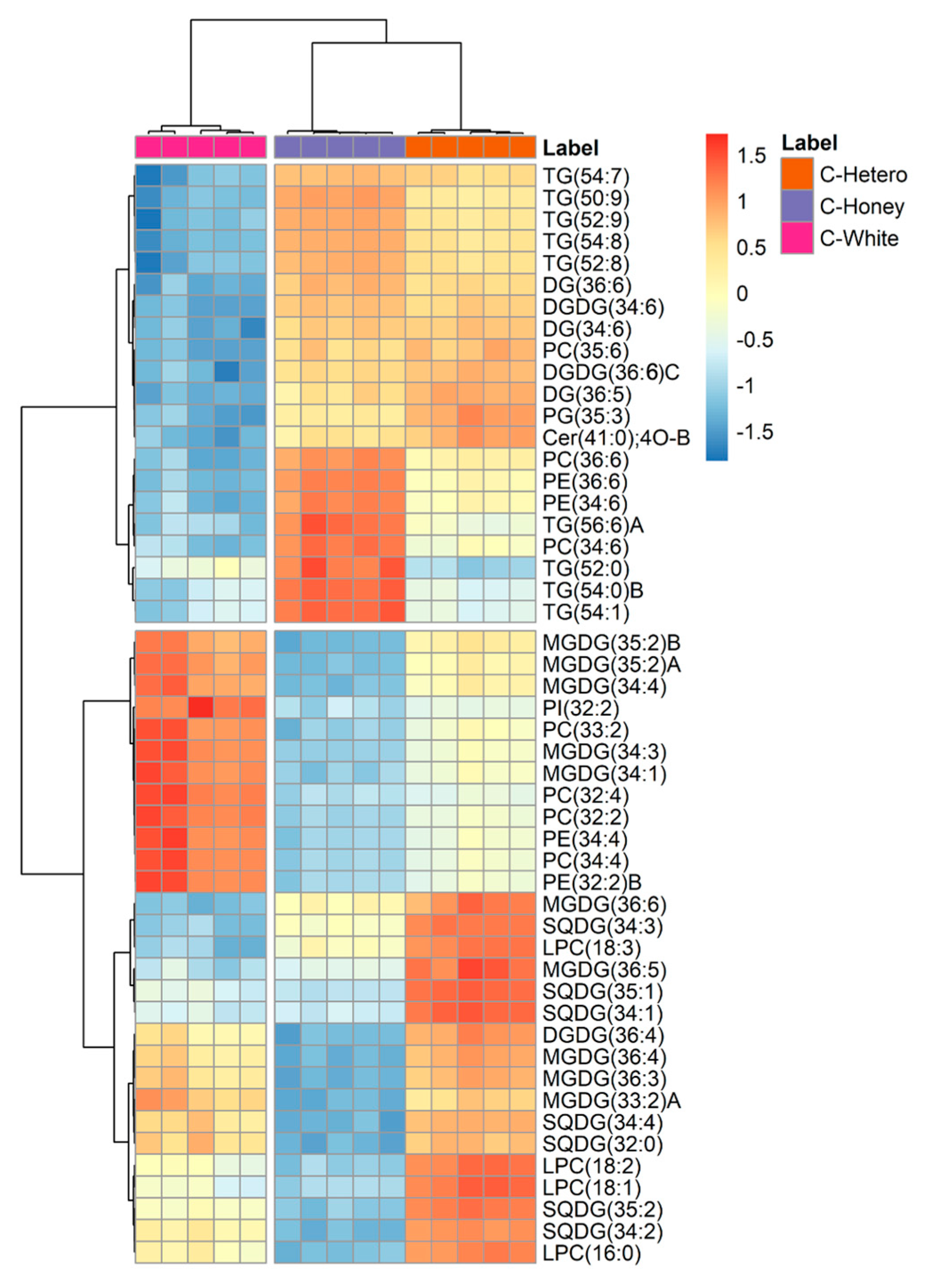
3.2. Lipidomics Profile of Chlorella vulgaris Strains
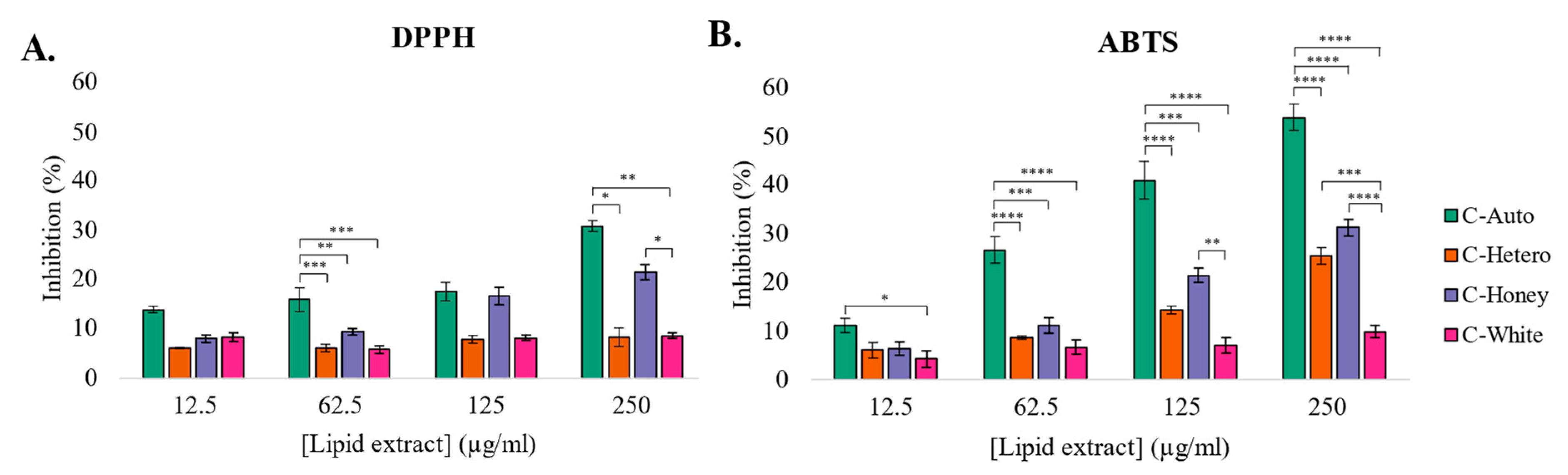
3.3. Antioxidant Activity of Lipid Extracts from C. vulgaris Strains
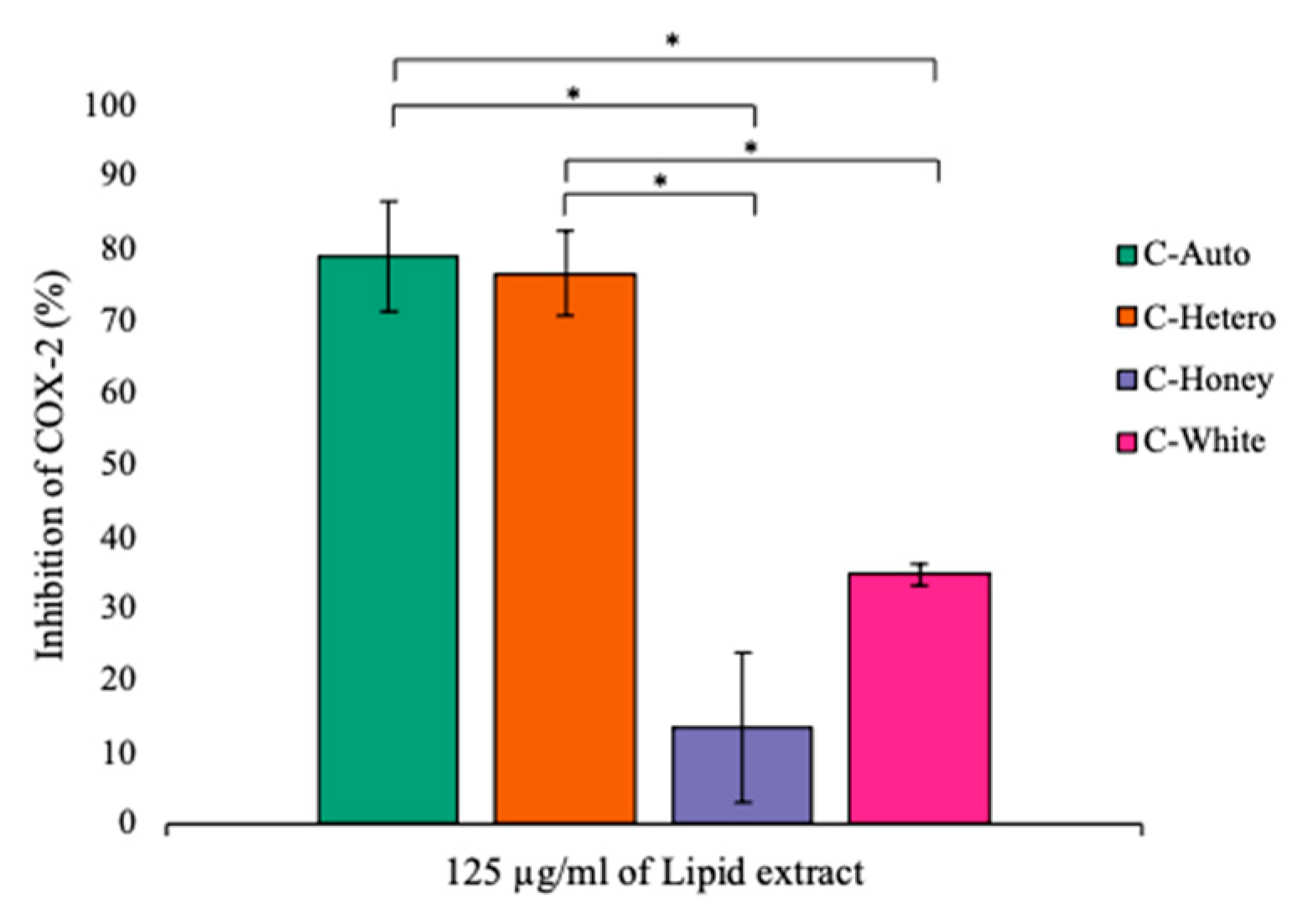
3.4. Anti-Inflammatory Activity of Lipid Extracts of C. vulgaris Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levasseur, W.; Perré, P.; Pozzobon, V. A Review of High Value-Added Molecules Production by Microalgae in Light of the Classification. Biotechnol. Adv. 2020, 41, 107545. [Google Scholar] [CrossRef] [PubMed]
- Ru, I.T.K.; Sung, Y.Y.; Jusoh, M.; Wahid, M.E.A.; Nagappan, T. Chlorella Vulgaris: A Perspective on Its Potential for Combining High Biomass with High Value Bioproducts. Appl. Phycol. 2020, 1, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Barros, A.; Pereira, H.; Campos, J.; Marques, A.; Varela, J.; Silva, J. Heterotrophy as a Tool to Overcome the Long and Costly Autotrophic Scale-up Process for Large Scale Production of Microalgae. Sci. Rep. 2019, 9, 13935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elalami, D.; Oukarroum, A.; Barakat, A. Anaerobic Digestion and Agronomic Applications of Microalgae for Its Sustainable Valorization. RSC Adv. 2021, 11, 26444–26462. [Google Scholar] [CrossRef]
- Lafarga, T. Effect of Microalgal Biomass Incorporation into Foods: Nutritional and Sensorial Attributes of the End Products. Algal Res. 2019, 41, 101566. [Google Scholar] [CrossRef]
- Schüler, L.; Greque de Morais, E.; Trovão, M.; Machado, A.; Carvalho, B.; Carneiro, M.; Maia, I.; Soares, M.; Duarte, P.; Barros, A.; et al. Isolation and Characterization of Novel Chlorella Vulgaris Mutants with Low Chlorophyll and Improved Protein Contents for Food Applications. Front. Bioeng. Biotechnol. 2020, 8, 469. [Google Scholar] [CrossRef]
- Couto, D.; Melo, T.; Conde, T.A.; Costa, M.; Silva, J.; Domingues, M.R.M.; Domingues, P. Chemoplasticity of the Polar Lipid Profile of the Microalgae Chlorella Vulgaris Grown under Heterotrophic and Autotrophic Conditions. Algal Res. 2021, 53, 102128. [Google Scholar] [CrossRef]
- Yao, L.; Gerde, J.A.; Lee, S.-L.; Wang, T.; Harrata, K.A. Microalgae Lipid Characterization. J. Agric. Food Chem. 2015, 63, 1773–1787. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Sun, B.; She, X.; Zhao, F.; Cao, Y.; Ren, D.; Lu, J. Lipid Production and Composition of Fatty Acids in Chlorella Vulgaris Cultured Using Different Methods: Photoautotrophic, Heterotrophic, and Pure and Mixed Conditions. Ann. Microbiol. 2014, 64, 1239–1246. [Google Scholar] [CrossRef]
- Djuricic, I.; Calder, P.C. Beneficial Outcomes of Omega-6 and Omega-3 Polyunsaturated Fatty Acids on Human Health: An Update for 2021. Nutrients 2021, 13, 2421. [Google Scholar] [CrossRef]
- Lordan, R.; Tsoupras, A.; Zabetakis, I. Phospholipids of Animal and Marine Origin: Structure, Function, and Anti-Inflammatory Properties. Mol. J. Synth. Chem. Nat. Prod. Chem. 2017, 22, 1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lordan, R.; Redfern, S.; Tsoupras, A.; Zabetakis, I. Inflammation and Cardiovascular Disease: Are Marine Phospholipids the Answer? Food Funct. 2020, 11, 2861–2885. [Google Scholar] [CrossRef] [PubMed]
- Couto, D.; Melo, T.; Conde, T.A.; Moreira, A.S.P.; Ferreira, P.; Costa, M.; Silva, J.; Domingues, R.; Domingues, P. Food Grade Extraction of Chlorella Vulgaris Polar Lipids: A Comparative Lipidomic Study. Food Chem. 2022, 375, 131685. [Google Scholar] [CrossRef] [PubMed]
- Breil, C.; Abert Vian, M.; Zemb, T.; Kunz, W.; Chemat, F. “Bligh and Dyer” and Folch Methods for Solid–Liquid–Liquid Extraction of Lipids from Microorganisms. Comprehension of Solvatation Mechanisms and towards Substitution with Alternative Solvents. Int. J. Mol. Sci. 2017, 18, 708. [Google Scholar] [CrossRef] [Green Version]
- Couto, D.; Conde, T.A.; Melo, T.; Neves, B.; Costa, M.; Cunha, P.; Guerra, I.; Correia, N.; Silva, J.T.; Pereira, H.; et al. Effects of Outdoor and Indoor Cultivation on the Polar Lipid Composition and Antioxidant Activity of Nannochloropsis Oceanica and Nannochloropsis Limnetica: A Lipidomics Perspective. Algal Res. 2022, 64, 102718. [Google Scholar] [CrossRef]
- Melo, T.; Alves, E.; Azevedo, V.; Martins, A.S.; Neves, B.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Lipidomics as a New Approach for the Bioprospecting of Marine Macroalgae—Unraveling the Polar Lipid and Fatty Acid Composition of Chondrus Crispus. Algal Res. 2015, 8, 181–191. [Google Scholar] [CrossRef]
- Ulbricht, T.L.V.; Southgate, D.A.T. Coronary Heart Disease: Seven Dietary Factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Hulbert, A.J.; Pamplona, R.; Buffenstein, R.; Buttemer, W.A. Life and Death: Metabolic Rate, Membrane Composition, and Life Span of Animals. Physiol. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef]
- Lai, Z.; Tsugawa, H.; Wohlgemuth, G.; Mehta, S.; Mueller, M.; Zheng, Y.; Ogiwara, A.; Meissen, J.; Showalter, M.; Takeuchi, K.; et al. Identifying Epimetabolites by Integrating Metabolome Databases with Mass Spectrometry Cheminformatics. Nat. Methods 2018, 15, 53–56. [Google Scholar] [CrossRef]
- Tsugawa, H.; Ikeda, K.; Takahashi, M.; Satoh, A.; Mori, Y.; Uchino, H.; Okahashi, N.; Yamada, Y.; Tada, I.; Bonini, P.; et al. A Lipidome Atlas in MS-DIAL 4. Nat. Biotechnol. 2020, 38, 1159–1163. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Allaire, J.J. RStudio: Integrated Development Environment for R; PBC: Boston, MA, USA, 2016; Available online: http://www.Rstudio.com/ (accessed on 24 February 2023).
- Alboukadel Kassambara Rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R Package Version 0.7.2. 2023. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 24 February 2023).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Package Version 1.0.7. R Packag Version. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 24 February 2023).
- Kolde, R. Pheatmap: Pretty Heatmaps, R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 24 February 2023).
- Šimat, V.; Hamed, I.; Petričević, S.; Bogdanović, T. Seasonal Changes in Free Amino Acid and Fatty Acid Compositions of Sardines, Sardina Pilchardus (Walbaum, 1792): Implications for Nutrition. Foods 2020, 9, 867. [Google Scholar] [CrossRef] [PubMed]
- Conde, T.A.; Neves, B.F.; Couto, D.; Melo, T.; Neves, B.; Costa, M.; Silva, J.; Domingues, P.; Domingues, M.R. Microalgae as Sustainable Bio-Factories of Healthy Lipids: Evaluating Fatty Acid Content and Antioxidant Activity. Mar. Drugs 2021, 19, 357. [Google Scholar] [CrossRef]
- Matos, Â.P.; Feller, R.; Moecke, E.H.S.; de Oliveira, J.V.; Junior, A.F.; Derner, R.B.; Sant’Anna, E.S. Chemical Characterization of Six Microalgae with Potential Utility for Food Application. J. Am. Oil Chem. Soc. 2016, 93, 963–972. [Google Scholar] [CrossRef]
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of Temperature and Nitrogen Concentration on the Growth and Lipid Content of Nannochloropsis Oculata and Chlorella Vulgaris for Biodiesel Production. Chem. Eng. Process. Process Intensif. 2009, 48, 1146–1151. [Google Scholar] [CrossRef]
- Parichehreh, R.; Gheshlaghi, R.; Mahdavi, M.A.; Elkamel, A. Optimization of Lipid Production in Chlorella Vulgaris for Biodiesel Production Using Flux Balance Analysis. Biochem. Eng. J. 2019, 141, 131–145. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Omega-3 and Omega-6 Polyunsaturated Fatty Acids: Dietary Sources, Metabolism, and Significance—A Review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Ramesh Kumar, B.; Deviram, G.; Mathimani, T.; Duc, P.A.; Pugazhendhi, A. Microalgae as Rich Source of Polyunsaturated Fatty Acids. Biocatal. Agric. Biotechnol. 2019, 17, 583–588. [Google Scholar] [CrossRef]
- Farag, M.A.; Gad, M.Z. Omega-9 Fatty Acids: Potential Roles in Inflammation and Cancer Management. J. Genet. Eng. Biotechnol. 2022, 20, 48. [Google Scholar] [CrossRef]
- Santa-María, C.; López-Enríquez, S.; Montserrat-de la Paz, S.; Geniz, I.; Reyes-Quiroz, M.E.; Moreno, M.; Palomares, F.; Sobrino, F.; Alba, G. Update on Anti-Inflammatory Molecular Mechanisms Induced by Oleic Acid. Nutrients 2023, 15, 224. [Google Scholar] [CrossRef] [PubMed]
- Santos-Silva, J.; Bessa, R.J.B.; Santos-Silva, F. Effect of Genotype, Feeding System and Slaughter Weight on the Quality of Light Lambs II. Fatty Acid Composition of Meat. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Yang, Y.; Ge, S.; Chen, Q.; Lin, S.; Zeng, S.; Tan, B.K.; Hu, J. Chlorella Unsaturated Fatty Acids Suppress High-Fat Diet-Induced Obesity in C57/BL6J Mice. J. Food Sci. 2022, 87, 3644–3658. [Google Scholar] [CrossRef]
- Regueiras, A.; Huguet, Á.; Conde, T.; Couto, D.; Domingues, P.; Domingues, M.R.; Costa, A.M.; da Silva, J.L.; Vasconcelos, V.; Urbatzka, R. Potential Anti-Obesity, Anti-Steatosis, and Anti-Inflammatory Properties of Extracts from the Microalgae Chlorella Vulgaris and Chlorococcum Amblystomatis under Different Growth Conditions. Mar. Drugs 2022, 20, 9. [Google Scholar] [CrossRef]
- Da Costa, E.; Melo, T.; Reis, M.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Polar Lipids Composition, Antioxidant and Anti-Inflammatory Activities of the Atlantic Red Seaweed Grateloupia Turuturu. Mar. Drugs 2021, 19, 414. [Google Scholar] [CrossRef] [PubMed]
- Indelicato, S.; Bongiorno, D.; Pitonzo, R.; Di Stefano, V.; Calabrese, V.; Indelicato, S.; Avellone, G. Triacylglycerols in Edible Oils: Determination, Characterization, Quantitation, Chemometric Approach and Evaluation of Adulterations. J. Chromatogr. A 2017, 1515, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lenihan-Geels, G.; Bishop, K.; Ferguson, L. Alternative Sources of Omega-3 Fats: Can We Find a Sustainable Substitute for Fish? Nutrients 2013, 5, 1301–1315. [Google Scholar] [CrossRef]
- Santigosa, E.; Brambilla, F.; Milanese, L. Microalgae Oil as an Effective Alternative Source of EPA and DHA for Gilthead Seabream (Sparus Aurata) Aquaculture. Animals 2021, 11, 971. [Google Scholar] [CrossRef]
- Ytrestøyl, T.; Aas, T.S.; Åsgård, T. Utilisation of Feed Resources in Production of Atlantic Salmon (Salmo Salar) in Norway. Aquaculture 2015, 448, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, E.; Silva, J.; Mendonça, S.; Abreu, M.; Domingues, M. Lipidomic Approaches towards Deciphering Glycolipids from Microalgae as a Reservoir of Bioactive Lipids. Mar. Drugs 2016, 14, 101. [Google Scholar] [CrossRef] [Green Version]
- Alhajj, M.J.; Montero, N.; Yarce, C.J.; Salamanca, C.H. Lecithins from Vegetable, Land, and Marine Animal Sources and Their Potential Applications for Cosmetic, Food, and Pharmaceutical Sectors. Cosmetics 2020, 7, 87. [Google Scholar] [CrossRef]
- Ma, A.-C.; Chen, Z.; Wang, T.; Song, N.; Yan, Q.; Fang, Y.-C.; Guan, H.-S.; Liu, H.-B. Isolation of the Molecular Species of Monogalactosyldiacylglycerols from Brown Edible Seaweed Sargassum Horneri and Their Inhibitory Effects on Triglyceride Accumulation in 3T3-L1 Adipocytes. J. Agric. Food Chem. 2014, 62, 11157–11162. [Google Scholar] [CrossRef] [PubMed]
- Makhlouf Brahmi, M.; Portmann, C.; D’Ambrosio, D.; Woods, T.M.; Banfi, D.; Reichenbach, P.; Da Silva, L.; Baudat, E.; Turcatti, G.; Lingner, J.; et al. Telomerase Inhibitors from Cyanobacteria: Isolation and Synthesis of Sulfoquinovosyl Diacylglycerols from Microcystis Aeruguinosa PCC 7806. Chem.-Eur. J. 2013, 19, 4596–4601. [Google Scholar] [CrossRef] [PubMed]
- Mizushina, Y.; Sugiyama, Y.; Yoshida, H.; Hanashima, S.; Yamazaki, T.; Kamisuki, S.; Ohta, K.; Takemura, M.; Yamaguchi, T.; Matsukage, A.; et al. Galactosyldiacylglycerol, a Mammalian DNA Polymerase Alpha-Specific Inhibitor from a Sea Alga, Petalonia Bingbamiae. Biol. Pharm. Bull. 2001, 24, 982–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New Acylated Sulfoglycolipids and Digalactolipids and Related Known Glycolipids from Cyanobacteria with a Potential to Inhibit the Reverse Transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar] [CrossRef]
- Da Costa, E.; Amaro, H.M.; Melo, T.; Guedes, A.C.; Domingues, M.R. Screening for Polar Lipids, Antioxidant, and Anti-Inflammatory Activities of Gloeothece Sp. Lipid Extracts Pursuing New Phytochemicals from Cyanobacteria. J. Appl. Phycol. 2020, 32, 3015–3030. [Google Scholar] [CrossRef]
- Pagels, F.; Amaro, H.M.; Tavares, T.G.; Amil, B.F.; Guedes, A.C. Potential of Microalgae Extracts for Food and Feed Supplementation—A Promising Source of Antioxidant and Anti-Inflammatory Compounds. Life 2022, 12, 1901. [Google Scholar] [CrossRef]
- Conde, T.A.; Zabetakis, I.; Tsoupras, A.; Medina, I.; Costa, M.; Silva, J.; Neves, B.; Domingues, P.; Domingues, M.R. Microalgal Lipid Extracts Have Potential to Modulate the Inflammatory Response: A Critical Review. Int. J. Mol. Sci. 2021, 22, 9825. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | C-Auto | C-Hetero | C-Honey | C-White |
|---|---|---|---|---|
| C14:0 | 0.2 ± 0.0 a,b | 0.2 ± 0.0 a | 0.2 ± 0.0 | 0.2 ± 0.0 b |
| C15:0 | 0.4 ± 0.0 a,b,c | 0.1 ± 0.0 a | 0.1 ± 0.0 b | 0.1 ± 0.0 c |
| C16:0 | 15.6 ± 0.3 a,b | 23.7 ± 0.4 c | 24.7 ± 0.4 a | 26.3 ± 0.6 b,c |
| C16:1(n−9) | 1.1 ± 0.0 a,b | 2.1 ± 0.0 | 2.7 ± 0.1 a | 2.8 ± 0.2 b |
| C16:1(n−7) | 2.9 ± 0.1 a | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.1 ± 0.0 a |
| C16:2(n−6) | 7.2 ± 0.2 | 7.1 ± 0.2 | 3.8 ± 0.2 a | 8.6 ± 0.6 a |
| C17:0 | 0.7 ± 0.0 a | 0.3 ± 0.0 | 0.8 ± 1.0 | 0.2 ± 0.0 a |
| C16:3(n−3) | 14.8 ± 0.5 a,b | 3.4 ± 0.1 a | 4.7 ± 0.3 c | 1.0 ± 0.1 b,c |
| C18:0 | 5.5 ± 2.4 a | 5.6 ± 2.2 b | 9.6 ± 0.4 | 12.7 ± 3.4 a,b |
| C18:1(n−9) | 4.5 ± 0.4 a,b | 16.6 ± 0.7 a | 18.0 ± 0.2 b,c | 15.3 ± 0.8 c |
| C18:1(n−7) | 2.7 ± 0.1 a,b | 0.8 ± 0.1 c | 0.4 ± 0.0 b | 0.3 ± 0.1 a,c |
| C18:2(n−6) | 18.0 ± 0.5 a,b | 32.7 ± 0.7 a,c | 25.5 ± 0.3 c | 30.5 ± 1.8 b |
| C18:3(n−3) | 26.3 ± 0.8 a,b | 6.9 ± 0.1 a | 9.0 ± 0.2 c | 1.8 ± 0.1 b,c |
| C20:0 | 0.1 ± 0.0 a | 0.2 ± 0.0 b | 0.4 ± 0.0 a,b | 0.2 ± 0.1 |
| Σ SFA | 22.5 ± 1.5 a,b,c | 30.1 ± 1.8 a,d | 35.8 ± 1.1 b | 39.7 ± 3.4 c,d |
| Σ MUFA | 11.2 ± 0.4 a,b | 19.7 ± 0.8 a | 21.3 ± 0.3 b,c | 18.5 ± 1.0 c |
| Σ PUFA | 66.3 ± 1.5 a,b | 50.1 ± 1.0 | 43.0 ± 0.9 a | 41.9 ± 2.5 b |
| Σ (n−3) | 41.1 ± 1.3 a,b | 10.3 ± 0.2 a | 13.7 ± 0.5 c | 2.8 ± 0.3 b,c |
| Σ (n−6) | 25.2 ± 0.6 a,b | 39.8 ± 0.9 a | 29.3 ± 0.5 | 39.1 ± 2.3 b |
| Indexes | C-Auto | C-Hetero | C-Honey | C-White |
|---|---|---|---|---|
| n−6/n−3 | 0.6 ± 0.0 a,b | 3.9 ± 0.0 a | 2.1 ± 0.0 c | 13.9 ± 0.6 b,c |
| AI | 0.2 ± 0.0 a,b | 0.4 ± 0.0 c | 0.4 ± 0.0 a | 0.5 ± 0.0 b,c |
| TI | 0.2 ± 0.0 a,b | 0.5 ± 0.0 c | 0.5 ± 0.0 a | 1.1 ± 0.2 b,c |
| h/H | 4.7 ± 0.2 a | 2.8 ± 0.0 b | 2.5 ± 0.0 | 2.2 ± 0.1 a,b |
| PoxI | 107.8 ± 2.7 a | 61.1 ± 1.2 | 58.9 ± 1.4 | 45.1 ± 2.7 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurício, T.; Couto, D.; Lopes, D.; Conde, T.; Pais, R.; Batista, J.; Melo, T.; Pinho, M.; Moreira, A.S.P.; Trovão, M.; et al. Differences and Similarities in Lipid Composition, Nutritional Value, and Bioactive Potential of Four Edible Chlorella vulgaris Strains. Foods 2023, 12, 1625. https://doi.org/10.3390/foods12081625
Maurício T, Couto D, Lopes D, Conde T, Pais R, Batista J, Melo T, Pinho M, Moreira ASP, Trovão M, et al. Differences and Similarities in Lipid Composition, Nutritional Value, and Bioactive Potential of Four Edible Chlorella vulgaris Strains. Foods. 2023; 12(8):1625. https://doi.org/10.3390/foods12081625
Chicago/Turabian StyleMaurício, Tatiana, Daniela Couto, Diana Lopes, Tiago Conde, Rita Pais, Joana Batista, Tânia Melo, Marisa Pinho, Ana S. P. Moreira, Mafalda Trovão, and et al. 2023. "Differences and Similarities in Lipid Composition, Nutritional Value, and Bioactive Potential of Four Edible Chlorella vulgaris Strains" Foods 12, no. 8: 1625. https://doi.org/10.3390/foods12081625