
Can Glyphosate and Temperature Affect the Nutritional Lipid Quality in the Mussel Mytilus galloprovincialis?

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Mussels and Exposure to GLY and Temperature (T°C)
2.3. Moisture, Lipid and Fatty Acid Analyses
2.4. Lipid Nutritional Quality Indices (LNQIs)
2.5. Statistical Analyses
3. Results and Discussion
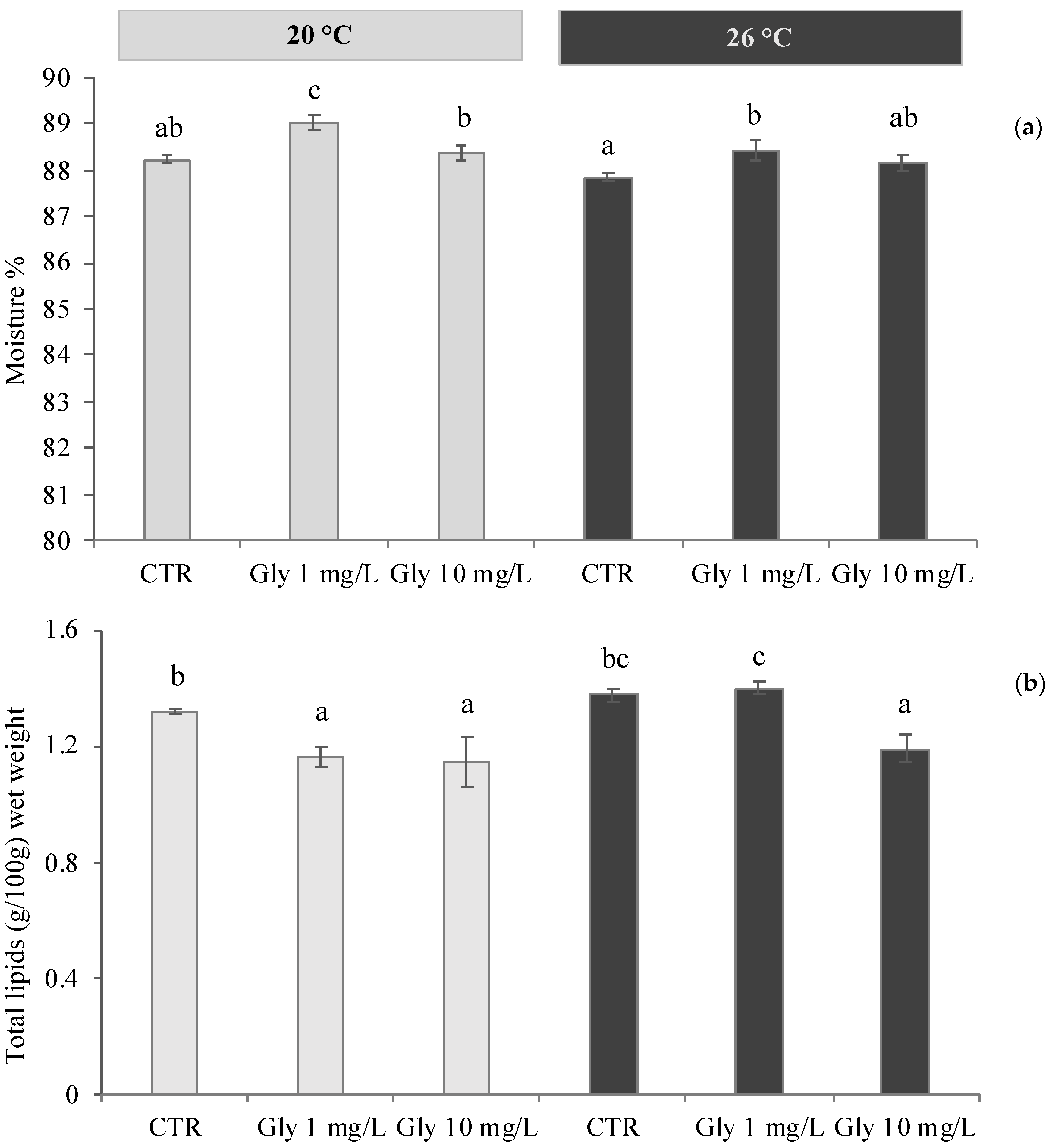
3.1. Effects of Temperature and Glyphosate on Moisture and Lipid Content of M. galloprovincialis
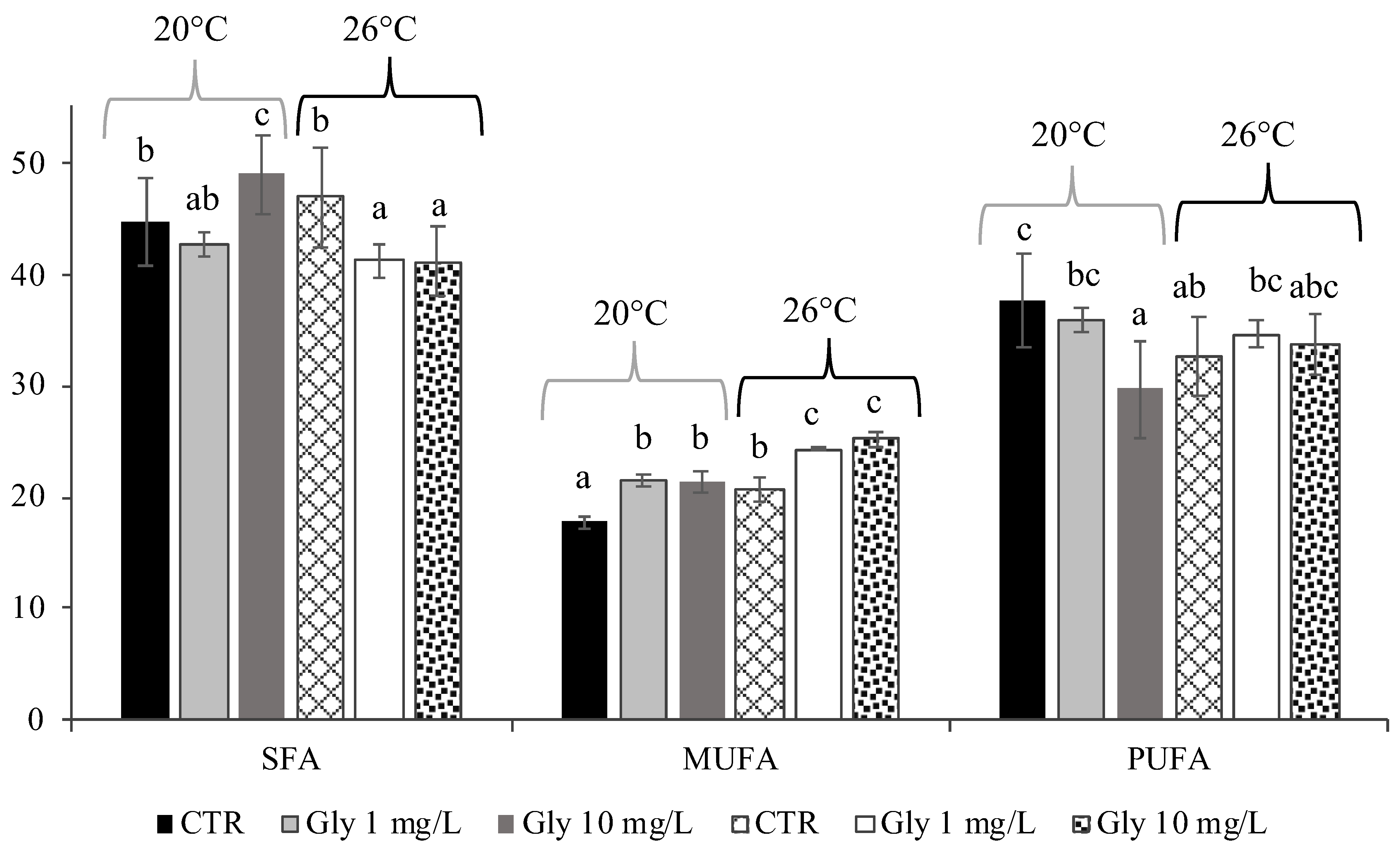
3.2. Effects of Temperature and Glyphosate on Fatty Acids of M. galloprovincialis
3.3. Lipid Nutritional Quality Indices
EPA + DHA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yaghubi, E.; Carboni, S.; Snipe, R.M.J.; Shaw, C.S.; Fyfe, J.J.; Smith, C.M.; Kaur, G.; Tan, S.-Y.; Hamilton, D.L. Farmed Mussels: A Nutritive Protein Source, Rich in Omega-3 Fatty Acids, with a Low Environmental Footprint. Nutrients 2021, 13, 1124. [Google Scholar] [CrossRef]
- European Market Observatory for Fisheries and Aquaculture Products (EUMOFA). Fresh Mussel in the EU; Case study; Publications Office of the European Union: Luxembourg, 2019. [Google Scholar]
- Biandolino, F.; Di Leo, A.; Parlapiano, I.; Papa, L.; Giandomenico, S.; Spada, L.; Prato, E. Nutritional quality of edible marine bivalves from the southern coast of Italy, Mediterranean Sea. Pol. J. Food Nutr. Sci. 2019, 69, 71–81. [Google Scholar] [CrossRef]
- Biandolino, F.; Parlapiano, I.; Denti, G.; Di Nardo, V.; Prato, E. Effect of different Cooking methods on lipid content and fatty acid profiles of Mytilus galloprovincialis. Foods 2021, 10, 416. [Google Scholar] [CrossRef] [PubMed]
- Prato, E.; Biandolino, F.; Parlapiano, I.; Giandomenico, S.; Denti, G.; Calò, M.; Di Leo, A. Proximate, fatty acids and metals in edible marine bivalves from Italian market: Beneficial and risk for consumers health. Sci. Total Environ. 2019, 648, 153–163. [Google Scholar] [CrossRef]
- Prato, E.; Fanelli, G.; Parlapiano, I.; Biandolino, F. Bioactive fatty acids in seafood from Ionian Sea and relation to dietary recommendations. Int. J. Food Sci. 2020, 71, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Merdzhanova, A.; Panayotova, V.; Dobreva, D.A.; Peycheva, K. Can fish and shellfish species from the Black Sea supply health beneficial amounts of bioactive fatty acids? Biomolecules 2021, 11, 1661. [Google Scholar] [CrossRef] [PubMed]
- Peycheva, K.; Panayotova, V.; Stancheva, R.; Makedonski, L.; Merdzhanova, A.; Cammilleri, G.; Ferrantelli, V.; Calabrese, V.; Cicero, N.; Fazio, F. Effect of steaming on chemical composition of Mediterranean mussel (Mytilus galloprovincialis): Evaluation of potential risk associated with human consumption. Food Sci. Nutr. 2022, 10, 3052–3061. [Google Scholar] [CrossRef]
- Budge, S.M.; Parrish, C.C.; McKenzie, C.H. Fatty acid composition of phytoplankton, settling particulate matter and sediments at a sheltered bivalve aquaculture site. Mar. Chem. 2001, 76, 285–303. [Google Scholar] [CrossRef]
- Jónasdóttir, S.H. Fatty acid profiles and production in marine phytoplankton. Mar. Drugs 2019, 17, 151. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Omega-3 fatty acids in inflammation an autoimmune diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet: The Omega-6/Omega-3 ratio and the brain. Mol. Neurobiol. 2011, 44, 203–215. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An Increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Bienias, J.L.; Tangney, C.C.; Bennett, D.A.; Wilson, R.S.; Aggarwal, N.; Schneider, J. Consumption of fish and n-3 fatty acids and risk of incident Alzheimer disease. Arch. Neurol. 2003, 60, 940–946. [Google Scholar] [CrossRef]
- Burdge, G.C.; Jones, A.E.; Wootton, S.A. Eicosapentaenoic and docosapentaenoic acids are the principal products of alpha-linolenic acid metabolism in young men. Br. J. Nutr. 2002, 88, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Childs, C.E.; Kew, S.; Finnegan, Y.E.; Minihane, A.M.; Leigh-Firbank, E.C.; Williams, C.M.; Calder, P.C. Increased dietary α-linolenic acid has sex-specific effects upon eicosapentaenoic acid status in humans: Re-examination of data from a randomised, placebo-controlled, parallel study. Nutr. J. 2014, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Harnack, K.; Andersen, G.; Somoza, V. Quantitation of alpha-linolenic acid elongation to eicosapentaenoic and docosahexaenoic acid as affected by the ratio of n6/n3 fatty acids. Nutr. Metab. 2009, 6, 8. [Google Scholar] [CrossRef] [PubMed]
- Plourde, M.; Cunnane, S.C. Extremely limited synthesis of long chain polyunsaturates in adults: Implications for their dietary essentiality and use as supplements. Appl. Physiol. Nutr. Metab. 2007, 32, 619–634. [Google Scholar] [CrossRef] [PubMed]
- Brenna, J.T.; Salem, N.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Carboni, S.; Kaur, G.; Pryce, A.; McKee, K.; Desbois, A.P.; Dick, J.R.; Galloway, S.D.R.; Hamilton, D.L. Mussel consumption as a ‘food first’ approach to improve omega-3 Status. Nutrients 2019, 11, 1381. [Google Scholar] [CrossRef] [PubMed]
- Baumard, P.; Budzinski, H.; Garrigues, P.; Narbonne, J.F.; Burgeot, T.; Michel, X.; Bellocq, J. Polycyclic aromatic hydrocarbon (PAH) burden of mussels (Mytilus sp.) indifferent marine environments in relation with sediment PAH contamination, and bioavailability. Mar. Environ. Res. 1999, 47, 415–439. [Google Scholar] [CrossRef]
- Brunner, E.J.; Jones, P.J.; Friel, S.; Bartley, M. Fish, human health and marine ecosystem health: Policies in collision. Int. J. Epidemiol. 2009, 38, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Karacik, B.; Okay, O.S.; Henkelmann, B.; Bernhöft, S.; Schramm, K.-W. Polycyclic aromatic hydrocarbons and effects on marine organisms in the Istanbul Strait. Environ. Int. 2009, 35, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Anacleto, P.; Maulvault, A.L.; Nunes, M.L.; Carvallo, M.L.; Rosa, R.; Marques, A. Effects of depuration on metal levels and health status of bivalve mollusks. Food Control 2015, 47, 493–501. [Google Scholar] [CrossRef]
- Kasiotis, K.M.; Emmanouil, C.; Anastasiadou, P.; Papadi-Psyllou, A.; Papadopoulos, A.; Okay, O.; Machera, K. Organic pollution and its effects in the marine mussel Mytilus galloprovincialis in Eastern Mediterranean coasts. Chemosphere 2015, 119, S145–S152. [Google Scholar] [CrossRef]
- Domingo, J.L. Nutrients and chemical pollutants in fish and shellfish: Balancing health benefits and risks of regular fish consumption. Crit. Rev. Food Sci. Nut. 2016, 56, 979–988. [Google Scholar] [CrossRef]
- Mititelu, M.; Neacs, S.M.; Oprea, E.; Dumitrescu, D.-E.; Nedelescu, M.; Drăgănescu, D.; Nicolescu, T.O.; Roşca, A.C.; Ghica, M. Black Sea mussels qualitative and quantitative chemical analysis: Nutritional benefits and possible risks through consumption. Nutrients 2022, 14, 964. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in Glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef]
- Matozzo, V.; Fabrello, J.; Marin, M.G. The effects of Glyphosate and its commercial formulations to marine invertebrates: A review. J. Mar. Sci. Eng. 2020, 8, 399. [Google Scholar] [CrossRef]
- Córdova López, A.M.; Almeida Sarmento, R.; de Souza Saraiva, A.; Ramos Pereira, R.; Soares, A.M.V.M.; Pestana, J.L.T. Exposure to Roundup® affects behaviour, head regeneration and reproduction of the freshwater planarian Girardia tigrina. Sci. Total Environ. 2019, 675, 453–461. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Mottier, A.; Séguin, A.; Devos, A.; Le Pabic, C.; Voiseux, C.; Lebel, J.M.; Serpentini, A.; Fiévet, B.; Costil, K. Effects of subchronic exposure to Glyphosate in juvenile oysters (Crassostrea gigas): From molecular to individual levels. Mar. Pollut. Bull. 2015, 95, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Iummato, M.M.; Sabatini, S.E.; Cacciatore, L.C.; Cochón, A.C.; Cataldo, D.; de Molina, M.D.; Juárez, Á.B. Biochemical responses of the golden mussel Limnoperna fortunei under dietary Glyphosate exposure. Ecotoxicol. Environ. Saf. 2018, 163, 69–75. [Google Scholar] [CrossRef]
- Filimonova, V.; Gonçalves, F.; Marques, J.C.; De Troch, M.; Goncalves, A.M. Fatty acid profiling as bioindicator of chemical stress in marine organisms: A review. Ecol. Indic. 2016, 67, 657–672. [Google Scholar] [CrossRef]
- Fouzai, C.; Trabelsi, W.; Bejaoui, S.; Telahigue, K.; Rabeh, I.; Nechi, S.; Chelbi, E.; El Cafsi, M.; Soudani, N. Cellular toxicity mechanisms of lambda-cyhalothrin in Venus verrucosa as revealed by fatty acid composition, redox status and histopathological changes. Ecol. Indic. 2020, 108, 105690. [Google Scholar] [CrossRef]
- Fadhlaoui, M.; Lavoie, I. Effects of temperature and Glyphosate on fatty acid composition, antioxidant capacity, and lipid peroxidation in the gastropod Lymneae sp. Water 2021, 13, 1039. [Google Scholar] [CrossRef]
- Telahigue, K.; Rabeh, I.; Hajji, T.; Trabelsi, W.; Fouzai, C.; Nechi, S.; Chelbi, E.; El Cafsi, M.; Soudani, N. Assessment of the impacts of glyphosate and its commercial formulation Roundup® on the respiratory tree of the sea cucumber Holothuria forskali using a multivariate biomarker approach. Chemosphere 2021, 269, 129376. [Google Scholar] [CrossRef] [PubMed]
- Telahigue, K.; Rabeh, I.; Mhadhbi, L.; Nechi, S.; Chelbi, E.; Ben Ali, M.; Hedfi, A.; Al-Harbi, M.S.; Hajji, T. Glyphosate exposure modulates lipid composition, histo-architecture and oxidative stress status and induces neurotoxicity in the smooth scallop Flexopecten glaber. Pestic. Biochem. Physiol. 2022, 184, 105099. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.M.; Greeley, M.S. Ecotoxicological indicators of water quality: Using multiresponse indicators to assess the health of aquatic ecosystems. Wat. Air Soil Poll. 2000, 123, 103–115. [Google Scholar] [CrossRef]
- Prato, E.; Danieli, A.; Maffia, M.; Biandolino, F. Lipid and fatty acid compositions of Mytilus galloprovincialis cultured in the Mar Grande of Taranto (Southern Italy): Feeding strategies and trophic relationships. Zool. Stud. 2010, 49, 211–219. [Google Scholar]
- Bongiorno, T.; Iacumin, L.; Tubaro, F.; Marcuzzo, E.; Sensidoni, A.; Tulli, F. Seasonal changes in technological and nutritional quality of Mytilus galloprovincialis from suspended culture in the Gulf of Trieste (North Adriatic Sea). Food Chem. 2015, 173, 355–362. [Google Scholar] [CrossRef]
- Azpeitia, K.; Ferrer, L.; Revilla, M.; Pagaldai, J.; Mendiola, D. Growth, biochemical profile, and fatty acid composition of mussel (Mytilus galloprovincialis Lmk.) cultured in the open ocean of the Bay of Biscay (northern Spain). Aquaculture 2016, 454, 95–108. [Google Scholar] [CrossRef]
- Rousch, J.; Bingham, S.; Sommerfeld, M. Changes in fatty acid profiles of thermo-intolerant and thermo-tolerant marine diatoms during temperature stress. J. Exp. Mar. Biol. Ecol. 2003, 295, 145–156. [Google Scholar] [CrossRef]
- Fuschino, J.; Guschina, I.A.; Dobson, G.; Yan, N.D.; Harwood, J.L.; Arts, M.T. Rising water temperatures alter lipid dynamics and reduce n-3 essential fatty acid concentrations in Scenedesmus obliquus (Chlorophyta). J. Phycol. 2011, 47, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.R.; Vetter, W.; Gaus, C.; Shaw, G.R.; Müller, J.F. Trace organic compounds in the marine environment. Mar. Pollut. Bull. 2002, 45, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Amoatey, P.; Baawain, M.S. Effects of pollution on freshwater aquatic organisms. Water Environ. Res. 2019, 91, 1272–1287. [Google Scholar] [CrossRef] [PubMed]
- Buccolieri, A.; Buccolieri, G.; Cardellicchio, N.; Dell’Atti, A.; Di Leo, A.; Maci, A.; Petronio, B.M. Distribution and speciation of metals in surface sediments of Taranto gulf (Ionian sea, Southern Italy). Ann. Chim. 2004, 94, 469–478. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Prato, L.; Chiantore, M.; Kelly, M.; Hughes, A.D.; James, P.; Ferranti, M.P.; Biandolino, F.; Parlapiano, I.; Sicuro, B.; Fanelli, G. Effect of formulated diets on the proximate composition and fatty acid profiles of sea urchin Paracentrotus lividus gonad. Aquac. Int. 2018, 26, 185–202. [Google Scholar] [CrossRef]
- Ulbricht, T.L.; Southgate, D.A.T. Coronary heart disease: Seven dietary factors. Lancet 1991, 338, 985–992. [Google Scholar] [CrossRef]
- Santos-Silva, J.; Bessa, R.J.B.; Santos-Silva, F. Effects of genotype, feeding system and slaughter weight on the quality of light lambs. II. Fatty acid composition of meat. Livest. Prod. Sci. 2002, 77, 187–194. [Google Scholar] [CrossRef]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A mini-review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]
- Pirestani, S.; Sahari, A.; Barzegar, M. Fatty acids changes during frozen storage in several fish species from South Caspian Sea. J. Agric. Sci. Technol. 2010, 12, 321–329. [Google Scholar]
- Frontera, J.L.; Vatnick, I.; Chaulet, A.; Rodríguez, E.M. Effects of glyphosate and polyoxyethylenamine on growth and energetic reserves in the freshwater crayfish Cherax quadricarinatus (Decapoda, Parastacidae). Arch. Environ. Contam. Toxicol. 2011, 61, 590–598. [Google Scholar] [CrossRef]
- Avigliano, L.; Fassiano, A.V.; Medesani, D.A.; Ríos De Molina, M.C.; Rodríguez, E.M. Effects of glyphosate on growth rate, metabolic rate and energy reserves of early juvenile crayfish, Cherax quadricarinatus M. Bull. Environ. Contam. Toxicol. 2014, 92, 631–635. [Google Scholar] [CrossRef]
- Rahman, M.S.; Rahman, M.S. Effects of elevated temperature on prooxidant-antioxidant homeostasis and redox status in the American oyster: Signaling pathways of cellular apoptosis during heat stress. Environ. Res. 2021, 196, 110428. [Google Scholar] [CrossRef]
- Noyola, J.; Mascarób, M.; Caamal-Monsreal, C.; Norena-Barroso, E.; Díaz, F.; Re, D.; Sánchez, A.; Rosas, C. Effect of temperature on energetic balance and fatty acid composition of early juveniles of Octopus maya. J. Exp. Mar. Biol. Ecol. 2013, 445, 156–165. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, C.; Gao, Q.; Dong, S.; Ye, Z.; Tian, X. Impact of water temperature on the growth and fatty acid profiles of juvenile sea cucumber Apostichopus japonicus (Selenka). J. Therm. Biol. 2016, 60, 155–161. [Google Scholar] [CrossRef]
- Cohen, Z.; Norman, H.A.; Heimer, Y.M. Potential use of substituted pyridazinones for selecting polyunsaturated fatty acid overproducing cell lines of algae. Phytochemistry 1993, 32, 259–264. [Google Scholar] [CrossRef]
- Piretti, M.V.; Zuppa, F.; Paglinca, G.; Taioli, F. Investigation of the seasonal variations of fatty acid. Constituents in selected tissue of the bivalve mollusc, Scapharca inaequivalvis (Bruguiere). Comp. Biochem. Physiol. 1988, 89, 183–187. [Google Scholar] [CrossRef]
- Thompson, P.A.; Guo, M.-X.; Harrison, P.J.; Whyte, J.N.C. Effects of variation in temperature. II. On the fatty acid composition of eight species of marine phytoplankton. J. Phycol. 1992, 28, 488–497. [Google Scholar]
- Dridi, S.; Romdhane, M.S.; El Cafsi, M. Nutritional quality in terms of lipid content and fatty acid composition of neural and polar lipids in the adductor muscle of the oyster Crassostrea gigas (Thunberg, 1794) farmed in the Bizert lagoon (Tunisia) in relation with sexual cycle and environmental settings. Egypt. J. Aquat. Res. 2017, 43, 329–336. [Google Scholar]
- Ezgeta-Balic, D.; Najdek, M.; Peharda, M.; Blazina, M. Seasonal fatty acid profile analysis to trace origin of food sources of four commercially important bivalves. Aquaculture 2012, 334–337, 89–100. [Google Scholar] [CrossRef]
- Fokina, N.N.; Ruokolainen, T.R.; Nemova, N.N.; Bakhmet, I.N. Changes of blue mussels Mytilus edulis L. lipid composition under cadmium and copper toxic effect. Biol. Trace Elem. Res. 2013, 154, 217–225. [Google Scholar] [CrossRef]
- Arts, M.T.; Ackman, R.G.; Holub, B.J. ‘Essential fatty acids’ in aquatic ecosystems: A crucial link between diet and human health and evolution. Can. J. Fish. Aquat. Sci. 2001, 58, 122–137. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). Climate Change: Implications for Food Safety; FAO: Rome, Italy, 2008. Available online: http://www.fao.org/docrep/010/i0195e/i0195e00.HTM. (accessed on 15 December 2022).
- Bayir, M.; Sirkeciogl, A.N.; Bayir, A.; Aras, M. Alterations in Fatty Acids of Polar Lipids in Salmo trutta on Long-term Exposure to a Glyphosate-Based Herbicide (Roundup ®). Pak. J. Biol. Sci. 2013, 16, 1194–1198. [Google Scholar] [CrossRef]
- Mariamenatu, A.H.; Abdu, E.M. Overconsumption of omega-6 polyunsaturated fatty acids (PUFAs) versus deficiency of omega-3 PUFAs in modern-day diets: The disturbing factor for their “balanced antagonistic metabolic functions” in the human body. J. Lipids 2021, 2021, 8848161. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the ω-6/ω-3 Balance in Health and Disease: Evolutionary Aspects of Diet. In Healthy Agriculture, Healthy Nutrition, Healthy People; Simopoulos, A.P., Ed.; Karger: Basel, Switzerland, 2011; Volume 102, pp. 10–21. [Google Scholar]
- HMSO. Nutritional Aspects of Cardiovascular Disease Report on Health and Social Subjects; HMSO: London, UK, 1994. [Google Scholar]
- Fernandes, C.E.; da Silva Vasconcelos, M.A.; de Almeida Ribeiro, M.; Sarubbo, L.A.; Andrade, S.A.C.; de Molo Filho, A.B. Nutritional and lipid profiles in marine fish species from Brasil. Food Chem. 2014, 160, 67–71. [Google Scholar] [CrossRef]
- Araujo, P.; Truzzi, C.; Belghit, I.; Antonucci, M. The impact of seawater warming on fatty acid composition and nutritional quality indices of Trematomus bernacchii from the Antarctic region. Food Chem. 2021, 365, 130500. [Google Scholar] [CrossRef]
- Łuczýnska, J.; Paszczyk, B.; Nowosad, J.; Łuczýnski, M.J. Mercury, fatty acids content and lipid quality indexes in muscles of freshwater and marine fish on the Polish market, risk assessment of fish consumption. Int. J. Environ. Res. Public Health 2017, 14, 1120. [Google Scholar] [CrossRef]
- Küçükgülmez, A.; Yanar, Y.; Çelik, M.; Ersor, B. Fatty acids profile, atherogenic, thrombogenic, and polyene lipid indices in golden grey mullet (Liza aurata) and gold band goatfish (Upeneus moluccensis) from Mediterranean Sea. J. Aquat. Food Prod. Technol. 2018, 27, 912–918. [Google Scholar] [CrossRef]
- Martin, A. Apports Nutritionnels Conseilles Pour la Population Française, 3rd ed.; Tec et Doc Lavoisier: Paris, France, 2001; p. 608. [Google Scholar]
- Health Council of the Netherlands. Guidelines to a Healthy Diet 2006; Health Council of the Netherlands: The Hague, The Netherlands, 2006.
- Superior Health Council of Belgium. Advisory Report: Recommendations and Claims Made on Omega-3 Fatty Acids; Advisory Report; Superior Health Council of Belgium: Brussel, Belgium, 2004; pp. 1–8.
- FAO/WHO. Diet, Nutrition, and the Prevention of Chronic Diseases; WHO Technical Reper Series; WHO: Geneva, Switzerland, 2003; Volume 916. [Google Scholar]



{kind=link}
{kind=link}
| 20 °C | 26 °C | |||||
|---|---|---|---|---|---|---|
| CTR | Gly 1 mg/L | Gly 10 mg/L | CTR | Gly 1 mg/L | Gly 10 mg/L | |
| C14:0 | 6.52 ± 1.28 | 5.59 ± 0.58 | 6.86 ± 0.30 | 7.74 ± 1.32 | 6.85 ± 0.27 | 5.52 ± 0.78 |
| C15:0 | 1.59 ± 0.18 | 1.18 ± 0.17 | 1.43 ± 0.12 | 1.20 ± 0.18 | 1.21 ± 0.12 | 1.32 ± 0.48 |
| C16:0 | 28.5 ± 2.9 | 27.5 ± 2.4 | 31.8 ± 0.6 | 30.9 ± 3.2 | 27.1 ± 1.4 | 26.1 ± 3.3 |
| C17:0 | 1.72 ± 0.25 ab | 1.96 ± 0.31 bc | 2.17 ± 0.13 c | 1.61 ± 0.03 ab | 1.52 ± 0.08 a | 1.92 ± 0.24 bc |
| C18:0 | 6.30 ± 0.35 c | 6.40 ± 0.23 c | 6.68 ± 0.218 c | 5.43 ± 0.29 b | 4.48 ± 0.21 a | 6.29 ± 0.26 c |
| ∑SFA | 44.6 ± 2.9 b | 42.6 ± 3.5 ab | 48.9 ± 1.0 c | 46.9 ± 4 b | 41.2 ± 1.5 a | 41.1 ± 4.1 a |
| C14:1 | 2.25 ± 0.19 b | 2.57 ± 0.58 b | 4.36 ± 0.34 c | 0.91 ± 0.12 a | 2.09 ± 0.32 b | 2.27 ± 0.22 b |
| C15:1 | 2.36 ± 0.38 b | 2.67 ± 0.57 b | 3.54 ± 0.46 c | 1.48 ± 0.32 a | 2.10 ± 0.11 ab | 1.94 ± 0.37 ab |
| C16:1 | 6.30 ± 0.80 a | 5.64 ± 0.15 a | 6.12 ± 0.38 a | 8.09 ± 0.61 b | 8.73 ± 0.326 b | 6.13 ± 0.98 a |
| C17:1 | 0.59 ± 0.12 ab | 0.61 ± 0.17 ab | 0.58 ± 0.10 ab | 0.81 ± 0.07 bc | 0.86 ± 0.11 c | 0.49 ± 0.03 a |
| C18:1 n-7 | 3.17 ± 0.06 ab | 3.41 ± 0.23 c | 3.40 ± 0.16 c | 3.04 ± 0.01 a | 2.98 ± 0.09 a | 3.29 ± 0.05 bc |
| C18:1 n-9 | 1.18 ± 0.36 a | 4.44 ± 0.88 b | 1.91 ± 0.35 a | 4.57 ± 1.41 b | 5.34 ± 0.91 b | 8.80 ± 1.29 c |
| C20:1 n-9 | 1.98 ± 0.25 bc | 2.11 ± 0.17 c | 1.52 ± 0.03 a | 1.70 ± 0.21 ab | 2.18 ± 0.13 c | 2.31 ± 0.25 c |
| ∑MUFA | 17.8 ± 0.5 a | 21.5 ± 0.9 b | 21.4 ± 0.51 b | 20.6 ± 1.1 b | 24.3 ± 0.7 c | 25.2 ± 1.6 c |
| C18:2 n-6 | 2.28 ± 0.06 ab | 2.50 ± 0.13 c | 2.18 ± 0.10 a | 2.13 ± 0.09 a | 2.37 ± 0.07 bc | 2.12 ± 0.08 a |
| C18:3 n-3 | 4.10 ± 0.37 ab | 3.85 ± 0.32 a | 3.69 ± 0.46 a | 3.98 ± 0.08 a | 4.58 ± 0.25 b | 3.79 ± 0.11 a |
| C18:4 n-3 | 2.05 ± 0.31 | 2.19 ± 0.10 | 1.74 ± 0.08 | 2.16 ± 0.08 | 1.77 ± 0.15 | 2.14 ± 0.45 |
| C22:0 + 20:3 n-6 | 0.54 ± 0.10 bc | 0.62 ± 0.09 c | 0.40 ± 0.02 ab | 0.32 ± 0.02 a | 0.66 ± 0.13 c | 0.56 ± 0.11 bc |
| C20:4 n-6 (ARA) | 3.24 ± 0.18 bc | 3.41 ± 0.40 c | 2.76 ± 0.16 ab | 2.58 ± 0.27 a | 3.01 ± 0.06 bc | 3.27 ± 0.53 bc |
| C20:5 n-3 (EPA) | 14.6 ± 1.1 c | 14 ± 1.4 b | 12.0 ± 0.7 a | 13 ± 1.4 b | 13.4 ± 0.4 b | 11.7 ± 0.9 a |
| C22:5 n-3 | 0.68 ± 0.11 a | 0.62 ± 0.14 a | 0.50 ± 0.03 a | 0.60 ± 0.10 a | 0.66 ± 0.07 a | 0.93 ± 0.12 b |
| C22:6n3 (DHA) | 10.0 ± 1.8 c | 8.73 ± 1.45 bc | 6.36 ± 0.85 a | 7.77 ± 0.87 ab | 8.10 ± 0.77 ab | 9.13 ± 1.38 bc |
| ∑ PUFA | 37.5 ± 3.1 c | 35.9 ± 3.4 bc | 29.7 ± 1.0 a | 32.5 ± 2.4 ab | 34.5 ± 1.5 bc | 33.6 ± 3.0 abc |
| n-3 | 31.4 ± 2.8 b | 29.4 ± 3.1 b | 24.3 ± 1.2 a | 27.5 ± 2.3 ab | 28.5 ± 1.4 b | 27.7 ± 2.3 ab |
| n-6 | 6.07 ± 0.33 c | 6.53 ± 0.30 c | 5.34 ± 0.17 ab | 5.03 ± 0.20 a | 6.04 ± 0.17 c | 5.95 ± 0.61 bc |
| 20 °C | 26 °C | |||||
|---|---|---|---|---|---|---|
| CTR | Gly 1 mg/L | Gly 10 mg/L | CTR | Gly 1 mg/L | Gly 10 mg/L | |
| C14:0 | 60.2 ± 11.2 bc | 44.8 ± 4.6 a | 55.9 ± 2.5 abc | 74.6 ± 9.5 d | 67.1 ± 3.1 cd | 46.1 ± 3.4 ab |
| C15:0 | 14.7 ± 1.6 | 9.50 ± 1.12 | 11.7 ± 1.0 | 11.6 ± 1.4 | 11.9 ± 0.9 | 11.0 ± 1.2 |
| C16:0 | 263 ± 27 c | 220 ± 19.5 ab | 259 ± 4.6 bc | 297 ± 14.8 c | 266 ± 11.6 c | 218 ± 12.7 a |
| C17:0 | 15.9 ± 2.3 | 15.7 ± 1.5 | 17.7 ± 1.1 | 15.5 ± 0.9 | 14.9 ± 0.6 | 16.0 ± 1.1 |
| C18:0 | 58.2 ± 3.2 c | 51.3 ± 1.8 b | 54.4 ± 1.4 bc | 53.3 ± 2.1 b | 44 ± 1.8 a | 52.4 ± 1.7 b |
| ∑SFA | 412 ± 36 b | 342 ± 28 a | 398 ± 8.3 b | 451 ± 16.8 b | 403 ± 18.7 b | 343 ± 17.8 a |
| C14:1 | 20.8 ± 1.7 b | 20.6 ± 3.1 b | 35.5 ± 2.7 c | 8.79 ± 0.84 a | 20.5 ± 1.8 b | 19.0 ± 1.8 b |
| C15:1 | 21.8 ± 3.5 b | 21.4 ± 2.5 b | 28.9 ± 3.7 c | 14.3 ± 0.7 a | 20.6 ± 1.5 b | 16.2 ± 1.9 ab |
| C16:1 | 58.2 ± 7.3 b | 45.2 ± 1.2 a | 49.9 ± 3.1 ab | 78 ± 4.9 c | 85.6 ± 1.9 c | 51.1 ± 3.1 ab |
| C17:1 | 5.44 ± 1.12 a | 4.89 ± 1.07 a | 4.75 ± 0.83 a | 7.83 ± 0.53 b | 8.42 ± 0.76 b | 4.11 ± 0.36 a |
| C18:1n7 | 29.3 ± 0.6 | 27.3 ± 1.8 | 27.7 ± 1.3 | 29.3 ± 0.4 | 29.2 ± 1.3 | 27.4 ± 1.2 |
| C18:1n9c | 10.9 ± 2.2 a | 35.6 ± 5.7 bc | 15.6 ± 2.1 ab | 44 ± 6.8 c | 52.3 ± 2.8 c | 73.3 ± 8.3 d |
| C20:1n9 | 18.3 ± 2.2 bc | 16.9 ± 1.4 b | 12.4 ± 0.3 a | 16.4 ± 1.5 b | 21.3 ± 1.5 c | 19.2 ± 1.4 bc |
| ∑MUFA | 164 ± 4.9 a | 172 ± 7.2 a | 174 ± 4.1 a | 199 ± 8.9 b | 238 ± 7.2 c | 210 ± 11.7 b |
| C18:2n6c | 21.1 ± 0.6 b | 20.0 ± 1.1 b | 17.8 ± 0.8 a | 20.5 ± 0.5 b | 23.2 ± 0.6 c | 17.7 ± 0.9 a |
| C18:3n3 | 37.8 ± 3.5 b | 30.9 ± 2.5 a | 30.1 ± 3.2 a | 38.3 ± 1.3 b | 44.9 ± 3.1 c | 31.6 ± 1.3 a |
| C18:4n3 | 19.9 ± 3.7 | 17.5 ± 0.8 | 14.2 ± 0.6 | 20.9 ± 1.1 | 17.3 ± 1.5 | 17.9 ± 1.2 |
| C22:0 + 20:3n6 | 5.04 ± 0.98 cd | 5.00 ± 1.01 cd | 3.2 ± 0.2 ab | 3.11 ± 0.31 a | 6.53 ± 0.58 d | 4.71 ± 0.83 bc |
| C20:4n6 | 30 ± 2.7 c | 27.3 ± 3.2 abc | 22.5 ± 1.3 a | 24.9 ± 1.6 ab | 29.5 ± 1.2 bc | 27.3 ± 2.8 abc |
| C20:5n3 (EPA) | 135 ± 11.7 c | 112 ± 11.3 ab | 98.0 ± 6.0 a | 125 ± 9.6 bc | 131 ± 3.8 bc | 97.3 ± 8.4 a |
| C22:5n3 | 6.32 ± 1.67 bc | 4.94 ± 0.94 ab | 4.07 ± 0.29 a | 5.78 ± 0.74 abc | 6.46 ± 0.51 bc | 7.76 ± 0.62 c |
| C22:6n3 (DHA) | 92.7 ± 11.3 c | 70.0 ± 10.1 b | 51.8 ± 6.1 a | 74.9 ± 8.3 bc | 79.4 ± 5.4 bc | 76.2 ± 9.6 bc |
| ∑ PUFA | 346 ± 31.7 d | 288 ± 24.7 abc | 242 ± 8.2 d | 314 ± 11.9 bcd | 339 ± 12.6 cd | 280 ± 11.3 ab |
| n-3 | 291 ± 21.3 d | 235 ± 14.6 ab | 198 ± 9.5 a | 265 ± 18.6 cd | 279 ± 11.7 cd | 231 ± 21.1 ab |
| n-6 | 56.1 ± 3.1 cd | 52.4 ± 2.3 bc | 43.5 ± 1.3 a | 48.5 ± 2.1 ab | 59.2 ± 2.3 d | 49.7 ± 3.7 b |
| 20 °C | 26 °C | |||||
|---|---|---|---|---|---|---|
| CTR | Gly 1 mg/L | Gly 10 mg/L | CTR | Gly 1 mg/L | Gly 10 mg/L | |
| EPA + DHA % | 24.6 ± 2.4 c | 22.7 ± 2.3 bc | 18.4 ± 1.5 a | 20.8 ± 2.3 ab | 21.5 ± 1.1 abc | 20.8 ± 2.0 ab |
| n-3/n-6 | 5.17 ± 0.47 bc | 4.48 ± 0.32 a | 4.56 ± 0.25 a | 5.45 ± 0.45 c | 4.71 ± 0.14 ab | 4.63 ± 0.13 a |
| n-6/n-3 | 0.19 ± 0.02 ab | 0.22 ± 0.02 bc | 0.22 ± 0.02 bc | 0.18 ± 0.02 a | 0.21 ± 0.01 bc | 0.21 ± 0.01 bc |
| PUFA/SFA | 0.85 ± 0.12 b | 0.85 ± 0.10 b | 0.61 ± 0.03 a | 0.70 ± 0.11 ab | 0.84 ± 0.10 b | 0.83 ± 0.09 b |
| UNS/SFA | 1.25 ± 0.16 abc | 1.35 ± 0.11 bc | 1.04 ± 0.04 a | 1.15 ± 0.16 ab | 1.43 ± 0.09 c | 1.44 ± 0.15 c |
| ARA/DHA | 0.33 ± 0.03 a | 0.40 ± 0.06 b | 0.44 ± 0.08 b | 0.34 ± 0.05 a | 0.37 ± 0.03 b | 0.36 ± 0.02 ab |
| ARA/EPA | 0.22 ± 0.02 ab | 0.24 ± 0.00 b | 0.23 ± 0.03 b | 0.20 ± 0.00 a | 0.22 ± 0.01 ab | 0.28 ± 0.01 c |
| AI | 0.99 ± 0.11 ab | 0.87 ± 0.13 a | 1.16 ± 0.06 b | 1.18 ± 0.15 b | 0.93 ± 0.07 a | 0.83 ± 0.16 a |
| TI | 0.38 ± 0.06 a | 0.38 ± 0.05 a | 0.50 ± 0.03 b | 0.44 ± 0.04 ab | 0.37 ± 0.03 a | 0.37 ± 0.08 a |
| h/H | 1.05 ± 0.15 bc | 1.15 ± 0.14 bc | 0.76 ± 0.05 a | 0.92 ± 0.14 ab | 1.11 ± 0.1 bc | 1.28 ± 0.18 c |
| HPI | 1.04 ± 0.14 abc | 1.16 ± 0.09 c | 0.86 ± 0.04 a | 0.88 ± 0.13 ab | 1.08 ± 0.0 bc | 1.24 ± 0.16 c |
| UI | 186 ± 11.7 c | 179 ± 17.2 bc | 150 ± 6.5 a | 163 ± 14.8 ab | 176 ± 7.5 bc | 173 ± 12.8 bc |
| FLQ | 24.6 ± 2.4 c | 22.7 ± 1.8 bc | 18.4 ± 1.6 a | 20.8 ± 2.3 ab | 21.5 ± 1.2 abc | 20.8 ± 2.0 ab |
| PI | 0.88 ± 0.06 c | 0.84 ± 0.07 c | 0.58 ± 0.06 a | 0.68 ± 0.108 ab | 0.80 ± 0.08 bc | 0.82 ± 0.06 c |
| T°C | Gly | EPA +DHA (mg/100 g ww) | Mussel Portion (g) Containing Recommended EPA + DHA | |
|---|---|---|---|---|
| 250 mg | 500 mg | |||
| 20 °C | CTR | 227 ± 23.7 c | 110 | 220 |
| 1 mg/L | 182 ± 13.1 b | 137 | 275 | |
| 10 mg/L | 150 ± 12.9 a | 167 | 334 | |
| 26 °C | CTR | 200 ± 15.6 bc | 125 | 250 |
| 1 mg/L | 211 ± 8.6 bc | 119 | 238 | |
| 10 mg/L | 173 ± 17.3 b | 144 | 288 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biandolino, F.; Prato, E.; Grattagliano, A.; Parlapiano, I. Can Glyphosate and Temperature Affect the Nutritional Lipid Quality in the Mussel Mytilus galloprovincialis? Foods 2023, 12, 1595. https://doi.org/10.3390/foods12081595
Biandolino F, Prato E, Grattagliano A, Parlapiano I. Can Glyphosate and Temperature Affect the Nutritional Lipid Quality in the Mussel Mytilus galloprovincialis? Foods. 2023; 12(8):1595. https://doi.org/10.3390/foods12081595
Chicago/Turabian StyleBiandolino, Francesca, Ermelinda Prato, Asia Grattagliano, and Isabella Parlapiano. 2023. "Can Glyphosate and Temperature Affect the Nutritional Lipid Quality in the Mussel Mytilus galloprovincialis?" Foods 12, no. 8: 1595. https://doi.org/10.3390/foods12081595