
Allelic Variation of Glu-A1 and Glu-B1 Genes in Winter Durum Wheat and Its Effect on Quality Parameters
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Field Experiment
2.3. Grain Quality Evaluation
2.4. Pasta Quality Evaluation
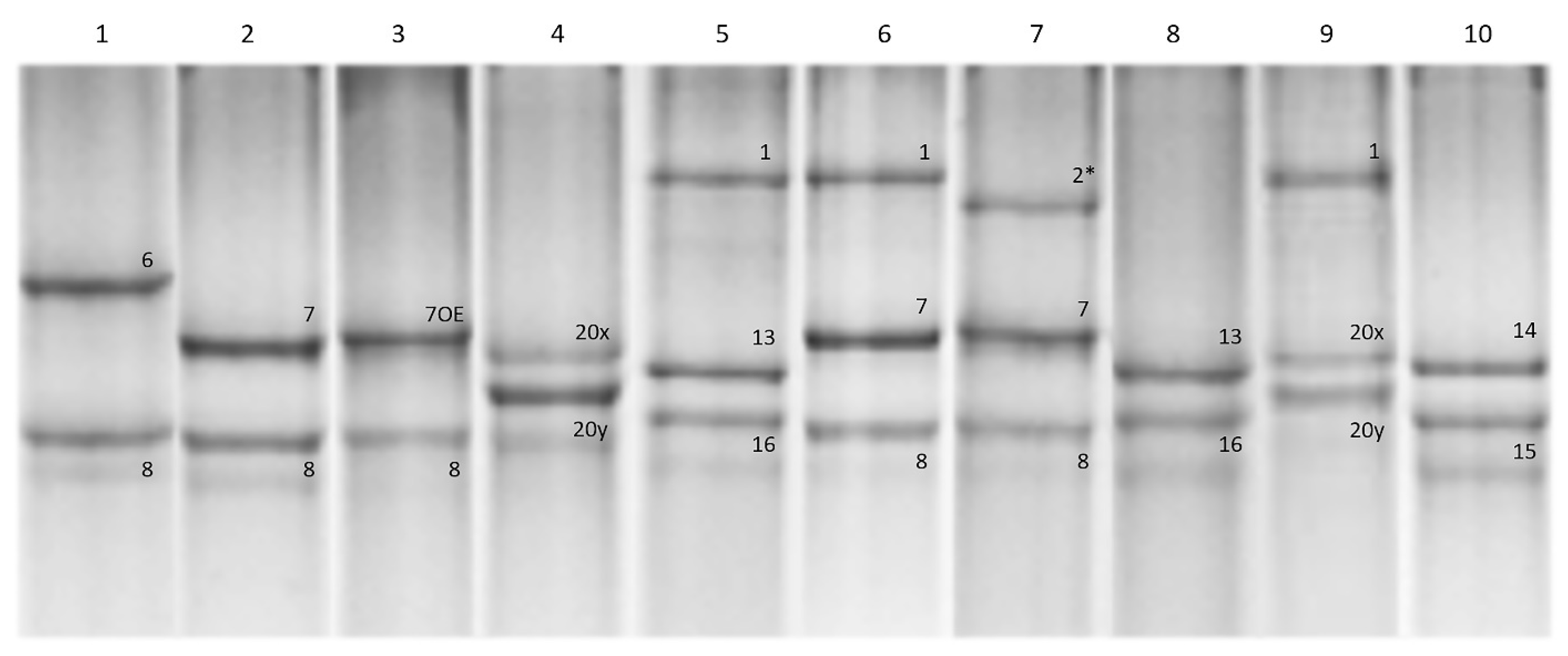
2.5. High Molecular Glutenin Characterisation
2.6. Glu-A1 and Glu-B1 Genotyping
2.7. Statistical Analyses
3. Results
3.1. Structure of the Collection
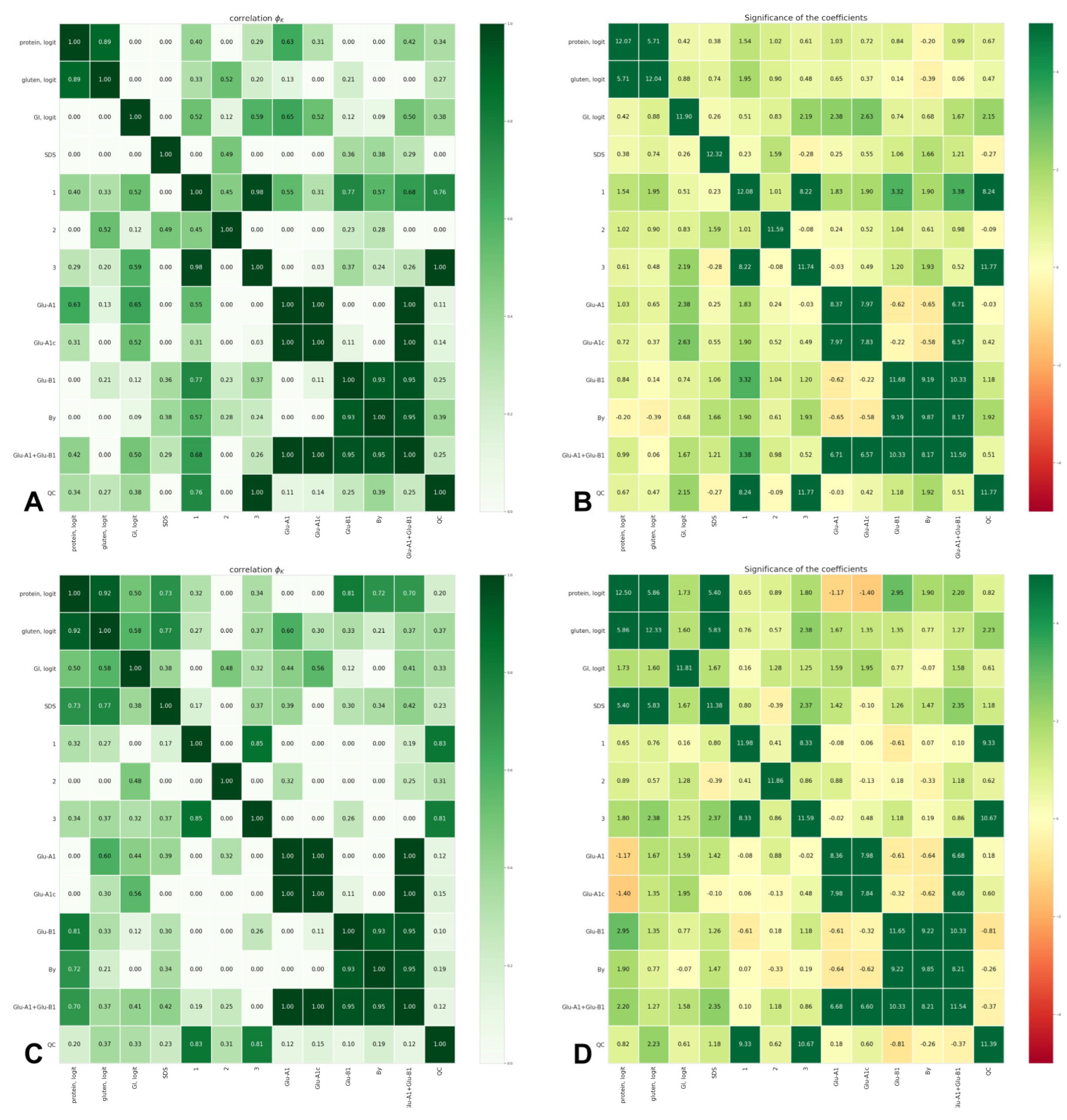
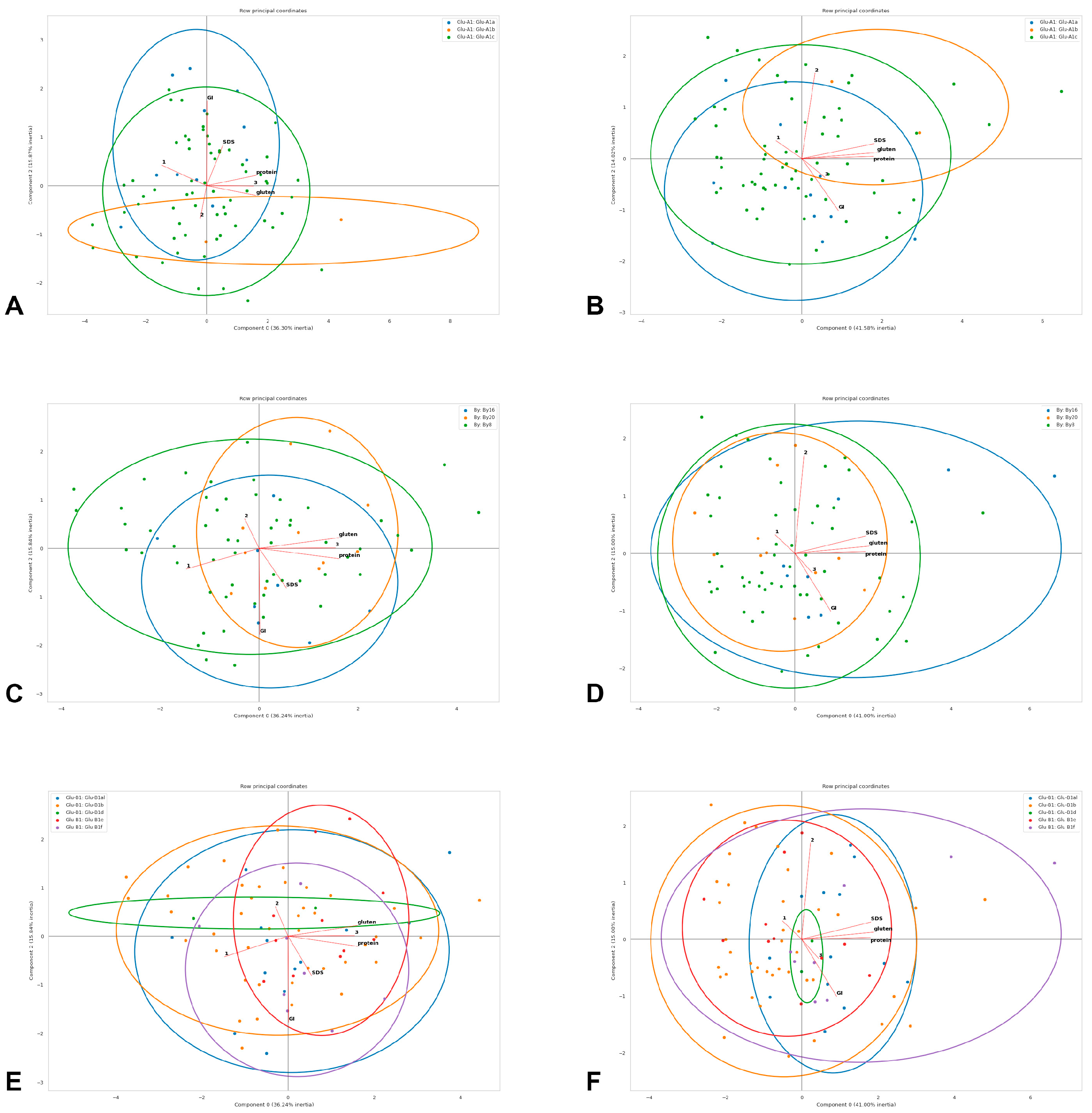
3.2. Associations between Quality Parameters
3.3. Gluten Index
3.4. Gluten Content
3.5. SDS Sedimentation Volume
3.6. Protein Content
3.7. Volume Increase Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Maximova, E.; Kulistikova, T. Ministry of Agriculture Expects to Increase Durum Wheat Production. Available online: https://www.agroinvestor.ru/markets/news/37398-minselkhoz-rasschityvaet-uvelichit-proizvodstvo-tverdoy-pshenitsy/ (accessed on 24 March 2023).
- Russian Pasta Market Analysis 2018–2022, Forecast for 2023–2027 under Sanctions. Available online: https://businesstat.ru/catalog/id8262/ (accessed on 24 March 2023).
- Statistics: World Pasta Production. Available online: https://www.pasta-unafpa.org/newt/unafpa/default.aspx?IDCONTENT=102 (accessed on 24 March 2023).
- Klimushina, M.V.; Divashuk, M.G.; Mokhammed, T.A.K.; Semenov, O.G.; Karlov, G.I. Analysis of Allelic State of Genes Responsible for Baking Properties in Allocytoplasmic Wheat Hybrids. Russ. J. Genet. 2013, 49, 530–538. [Google Scholar] [CrossRef]
- Mastrangelo, A.M.; Cattivelli, L. What Makes Bread and Durum Wheat Different? Trends Plant Sci. 2021, 26, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Padalino, L.; Mastromatteo, M.; Lecce, L.; Spinelli, S.; Contò, F.; Del Nobile, M.A. Effect of Durum Wheat Cultivars on Physico-Chemical and Sensory Properties of Spaghetti. J. Sci. Food Agric. 2014, 94, 2196–2204. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.X.; Wang, K.; Dupuis, B.; Taylor, D.; Nam, S. Kernel Vitreousness and Protein Content: Relationship, Interaction and Synergistic Effects on Durum Wheat Quality. J. Cereal Sci. 2018, 79, 210–217. [Google Scholar] [CrossRef]
- Kaplan Evlice, A. The Effect of Durum Wheat Genotypes on Cooking Quality of Pasta. Eur. Food Res. Technol. 2022, 248, 815–824. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.; Akhtar, H.; Zaheer, K.; Ali, R. Dietary Sources of Lutein and Zeaxanthin Carotenoids and Their Role in Eye Health. Nutrients 2013, 5, 1169–1185. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, M.; Prabhasankar, P. Health Based Pasta: Redefining the Concept of the Next Generation Convenience Food. Crit. Rev. Food Sci. Nutr. 2012, 52, 9–20. [Google Scholar] [CrossRef]
- Zhang, B.; Qiao, D.; Zhao, S.; Lin, Q.; Wang, J.; Xie, F. Starch-Based Food Matrices Containing Protein: Recent Understanding of Morphology, Structure, and Properties. Trends Food Sci. Technol. 2021, 114, 212–231. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, S.; Sun, B.; Wang, F.; Huang, J.; Wang, X.; Bao, Q. Effects of Thermal Properties and Behavior of Wheat Starch and Gluten on Their Interaction: A Review. Int. J. Biol. Macromol. 2021, 177, 474–484. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, F.; Sun, H.; Wang, J.; Duan, W.; Yan, Y. Wheat Glu-A1a Encoded 1Ax1 Subunit Enhances Gluten Physicochemical Properties and Molecular Structures That Confer Superior Breadmaking Quality. Int. J. Biol. Macromol. 2023, 225, 701–714. [Google Scholar] [CrossRef]
- Giraldo, P.; Ruiz, M.; Ibba, M.I.; Morris, C.F.; Labuschagne, M.T.; Igrejas, G. Durum Wheat Storage Protein Composition and the Role of LMW-GS in Quality. In Wheat Quality for Improving Processing and Human Health; Springer International Publishing: Cham, Switzerland, 2020; pp. 73–108. [Google Scholar]
- Martínez-Moreno, F.; Ammar, K.; Solís, I. Global Changes in Cultivated Area and Breeding Activities of Durum Wheat from 1800 to Date: A Historical Review. Agronomy 2022, 12, 1135. [Google Scholar] [CrossRef]
- Palamarchuk, A. Selection Strategies for Traits Relevant for Winter and Facultative Durum Wheat. In Durum Wheat Breeding: Current Approaches and Future Strategies; Royo, C., Nachit, M., Di Fonzo, N., Araus, J., Pfeiffer, W.H., Slafer, G.A., Eds.; Products Press: New York, NY, USA, 2005; pp. 599–644. ISBN 1-56022-333-2. [Google Scholar]
- Alabushev, A.V.; Makarova, T.S.; Samofalova, N.E.; Ilichkina, N.P.; Dubinina, O.A. Parameters of Adaptability and Stability of Winter Durum Wheat Varieties According to Various Forecrops in the Rostov Region. Agric. Sci. Euro-North-East 2019, 20, 557–566. [Google Scholar] [CrossRef]
- Longin, C.F.H.; Sieber, A.-N.; Reif, J.C. Combining Frost Tolerance, High Grain Yield and Good Pasta Quality in Durum Wheat. Plant Breed. 2013, 132, 353–358. [Google Scholar] [CrossRef]
- Samofalova, N.E.; Ilichkina, N.P.; Makarova, T.S.; Dubinina, O.A.; Kostylenko, O.A.; Kameneva, A.S.; Derova, T.G. Methods for the Development of the Initial Material in the Process of Winter Durum Wheat Breeding and Their Efciency. Grain Econ. Russ. 2020, 2, 54–60. [Google Scholar] [CrossRef]
- Kravchenko, N.S.; Samofalova, N.E.; Oldyreva, I.M.; Makarova, T.S. The Characteristics of the Winter Durum Wheat Varieties According to Grain Quality and Pasta Properties. Grain Econ. Russ. 2020, 3, 26–31. [Google Scholar] [CrossRef]
- Kameneva, A.S.; Ionova, E.V.; Marchenko, D.M.; Ilichkina, N.P.; Nekrasova, O.A. The Study of Collection Durum Winter Wheat Samples According to Grain Quality in the Rostov Region. Grain Econ. Russ. 2021, 1, 62–68. [Google Scholar] [CrossRef]
- Samofalova, N.E.; Ilichkina, N.P.; Bezuglaya, T.S.; Kravchenko, N.S.; Ivanisova, A.S.; Kabanova, N.V.; Dubinina, O.A. Correlation between Quality Traits of Kernels, Hard Semolina, Pasta of Durum Winter Wheat. Grain Econ. Russ. 2022, 4, 62–69. [Google Scholar] [CrossRef]
- Divashuk, M.G.; Bespalova, L.A.; Vasilyev, A.V.; Fesenko, I.A.; Puzyrnaya, O.Y.; Karlov, G.I. Reduced Height Genes and Their Importance in Winter Wheat Cultivars Grown in Southern Russia. Euphytica 2013, 190, 137–144. [Google Scholar] [CrossRef]
- Klimushina, M.V.; Gladkih, N.I.; Divashuk, M.G.; Bespalova, L.A.; Vasilyev, A.V.; Karlov, G.I. Distribution of Allelic Variants of Wx Genes in the Common Wheat Collection Made at the Krasnodar Lukyanenko Research Institute of Agriculture. Vavilov J. Genet. Breed. 2012, 16, 187–192. [Google Scholar]
- Bespalova, L.A.; Vasilyev, A.V.; Ablova, I.B.; Filobok, V.A.; Khudokormova, Z.N.; Davoyan, R.O.; Davoyan, E.R.; Karlov, G.I.; Soloviev, A.A.; Divashuk, M.G.; et al. Use of Molecular Markers in Wheat Breeding at the Krasnodar Lukyanenko Research Institute of Agriculture. Vavilov J. Genet. Breed. 2012, 16, 37–43. [Google Scholar]
- Bespalova, L.A.; Puchkov, Y.M.; Kolesnikov, F.A. Wheat Microevolution under Intensive Breeding Process in the Northern Caucasian Region. In Wheat Production in Stressed Environments; Springer: Dordrecht, The Netherlands; pp. 509–517.
- Bespalova, L.A. Adaptive Breeding of Winter Bread Wheat in Krasnodar. In Increasing Wheat Production in Central Asia through Science and International Cooperation, Proceedings of the First Central Asian Wheat Conference, Almaty, Kazakhstan, 10–13 June 2003; Morgunov, A., McNab, A., Camplbell, K.G., Paroda, R., Eds.; CIMMYT: Almaty, Kazakhstan, 2003; pp. 54–57. [Google Scholar]
- Romanenko, A.A.; Bespalova, L.A.; Mudrova, A.A.; Kudryashov, I.N.; Ablova, I.B.; Terpugova, N.I.; Samofalov, N.E.; Popov, A.S.; Revyakin, E.L. A Resource-Saving Technology of Winter Hard Wheat Growing; FGBNU “Rosinformagrotekh”: Moscow, Russia, 2013; ISBN 978-5-7367-0989-2. Available online: https://rosinformagrotech.ru/data/itpk/zernovye-kultury-i-kukuruza/send/17-zernovye-kultury-i-kukuruza/457-resursosberegayushchaya-tekhnologiya-proizvodstva-ozimoj-tverdoj-pshenitsy (accessed on 2 March 2023). (In Russian)
- Mudrova, A.A.; Yanovsky, A.S. The Results of Use of the Initial Material during Durum Winter Wheat Breeding in the Kuban Conditions. Grain Econ. Russ. 2016, 1, 24–27. [Google Scholar]
- Ablova, I.B. Resistance to Head Blight in Various Types of Wheat and Triticale in Krasnodar Environment. Cereal Res. Commun. 1997, 25, 715–720. [Google Scholar] [CrossRef]
- Chegdali, Y.; Ouabbou, H.; Essamadi, A.; Cervantes, F.; Ibba, M.I.; Guzmán, C. Assessment of the Glutenin Subunits Diversity in a Durum Wheat (T. turgidum ssp. durum) Collection from Morocco. Agronomy 2020, 10, 957. [Google Scholar] [CrossRef]
- Roncallo, P.F.; Guzmán, C.; Larsen, A.O.; Achilli, A.L.; Dreisigacker, S.; Molfese, E.; Astiz, V.; Echenique, V. Allelic Variation at Glutenin Loci (Glu-1, Glu-2 and Glu-3) in a Worldwide Durum Wheat Collection and Its Effect on Quality Attributes. Foods 2021, 10, 2845. [Google Scholar] [CrossRef] [PubMed]
- Dowell, F.E.; Maghirang, E.B.; Xie, F.; Lookhart, G.L.; Pierce, R.O.; Seabourn, B.W.; Bean, S.R.; Wilson, J.D.; Chung, O.K. Predicting Wheat Quality Characteristics and Functionality Using Near-Infrared Spectroscopy. Cereal Chem. J. 2006, 83, 529–536. [Google Scholar] [CrossRef]
- Sozinov, A.A.; Blokhin, N.I.; Vasilenko, I.I.; Sinitsyn, S.S.; Komarov, V.I.; Tarasenko, N.D.; Kravtsova, B.E. Methodological Guidelines for Assessing Grain Quality; Lenin All-Union Academy of Agricultural Sciences (VASKHNIL): Moscow, Russia, 1977. (In Russian) [Google Scholar]
- Singh, N.K.; Shepherd, K.W.; Cornish, G.B. A Simplified SDS—PAGE Procedure for Separating LMW Subunits of Glutenin. J. Cereal Sci. 1991, 14, 203–208. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid Isolation of High Molecular Weight Plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Rasheed, A.; Wen, W.; Gao, F.; Zhai, S.; Jin, H.; Liu, J.; Guo, Q.; Zhang, Y.; Dreisigacker, S.; Xia, X.; et al. Development and Validation of KASP Assays for Genes Underpinning Key Economic Traits in Bread Wheat. Theor. Appl. Genet. 2016, 129, 1843–1860. [Google Scholar] [CrossRef]
- Warton, D.I.; Hui, F.K.C. The Arcsine Is Asinine: The Analysis of Proportions in Ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Baak, M.; Koopman, R.; Snoek, H.; Klous, S. A New Correlation Coefficient between Categorical, Ordinal and Interval Variables with Pearson Characteristics. Comput. Stat. Data Anal. 2018, 152, 107043. [Google Scholar] [CrossRef]
- Sissons, M.J.; Ames, N.P.; Hare, R.A.; Clarke, J.M. Relationship between Glutenin Subunit Composition and Gluten Strength Measurements in Durum Wheat. J. Sci. Food Agric. 2005, 85, 2445–2452. [Google Scholar] [CrossRef]
- Nazco, R.; Peña, R.J.; Ammar, K.; Villegas, D.; Crossa, J.; Moragues, M.; Royo, C. Variability in Glutenin Subunit Composition of Mediterranean Durum Wheat Germplasm and Its Relationship with Gluten Strength. J. Agric. Sci. 2014, 152, 379–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Rudd, J.C.; Bai, G.; Haley, S.D.; Ibrahim, A.M.H.; Xue, Q.; Hays, D.B.; Graybosch, R.A.; Devkota, R.N.; St. Amand, P. Molecular Markers Linked to Important Genes in Hard Winter Wheat. Crop Sci. 2014, 54, 1304–1321. [Google Scholar] [CrossRef]
- Ragupathy, R.; Naeem, H.A.; Reimer, E.; Lukow, O.M.; Sapirstein, H.D.; Cloutier, S. Evolutionary Origin of the Segmental Duplication Encompassing the Wheat GLU-B1 Locus Encoding the Overexpressed Bx7 (Bx7OE) High Molecular Weight Glutenin Subunit. Theor. Appl. Genet. 2008, 116, 283–296. [Google Scholar] [CrossRef]
- Magallanes-López, A.M.; Ammar, K.; Morales-Dorantes, A.; González-Santoyo, H.; Crossa, J.; Guzmán, C. Grain Quality Traits of Commercial Durum Wheat Varieties and Their Relationships with Drought Stress and Glutenins Composition. J. Cereal Sci. 2017, 75, 1–9. [Google Scholar] [CrossRef]
- Cooper, J.K.; Stromberger, J.A.; Morris, C.F.; Bai, G.; Haley, S.D. End-Use Quality and Agronomic Characteristics Associated with the Glu-B1al High-Molecular-Weight Glutenin Allele in U.S. Hard Winter Wheat. Crop Sci. 2016, 56, 2348–2353. [Google Scholar] [CrossRef]
- Guzmán, C.; Crossa, J.; Mondal, S.; Govindan, V.; Huerta, J.; Crespo-Herrera, L.; Vargas, M.; Singh, R.P.; Ibba, M.I. Effects of Glutenins (Glu-1 and Glu-3) Allelic Variation on Dough Properties and Bread-Making Quality of CIMMYT Bread Wheat Breeding Lines. Field Crops Res. 2022, 284, 108585. [Google Scholar] [CrossRef]
- Giunta, F.; Bassu, S.; Mefleh, M.; Motzo, R. Is the Technological Quality of Old Durum Wheat Cultivars Superior to That of Modern Ones When Exposed to Moderately High Temperatures during Grain Filling? Foods 2020, 9, 778. [Google Scholar] [CrossRef]
- Graziano, S.; Marando, S.; Prandi, B.; Boukid, F.; Marmiroli, N.; Francia, E.; Pecchioni, N.; Sforza, S.; Visioli, G.; Gullì, M. Technological Quality and Nutritional Value of Two Durum Wheat Varieties Depend on Both Genetic and Environmental Factors. J. Agric. Food Chem. 2019, 67, 2384–2395. [Google Scholar] [CrossRef]





{kind=link}
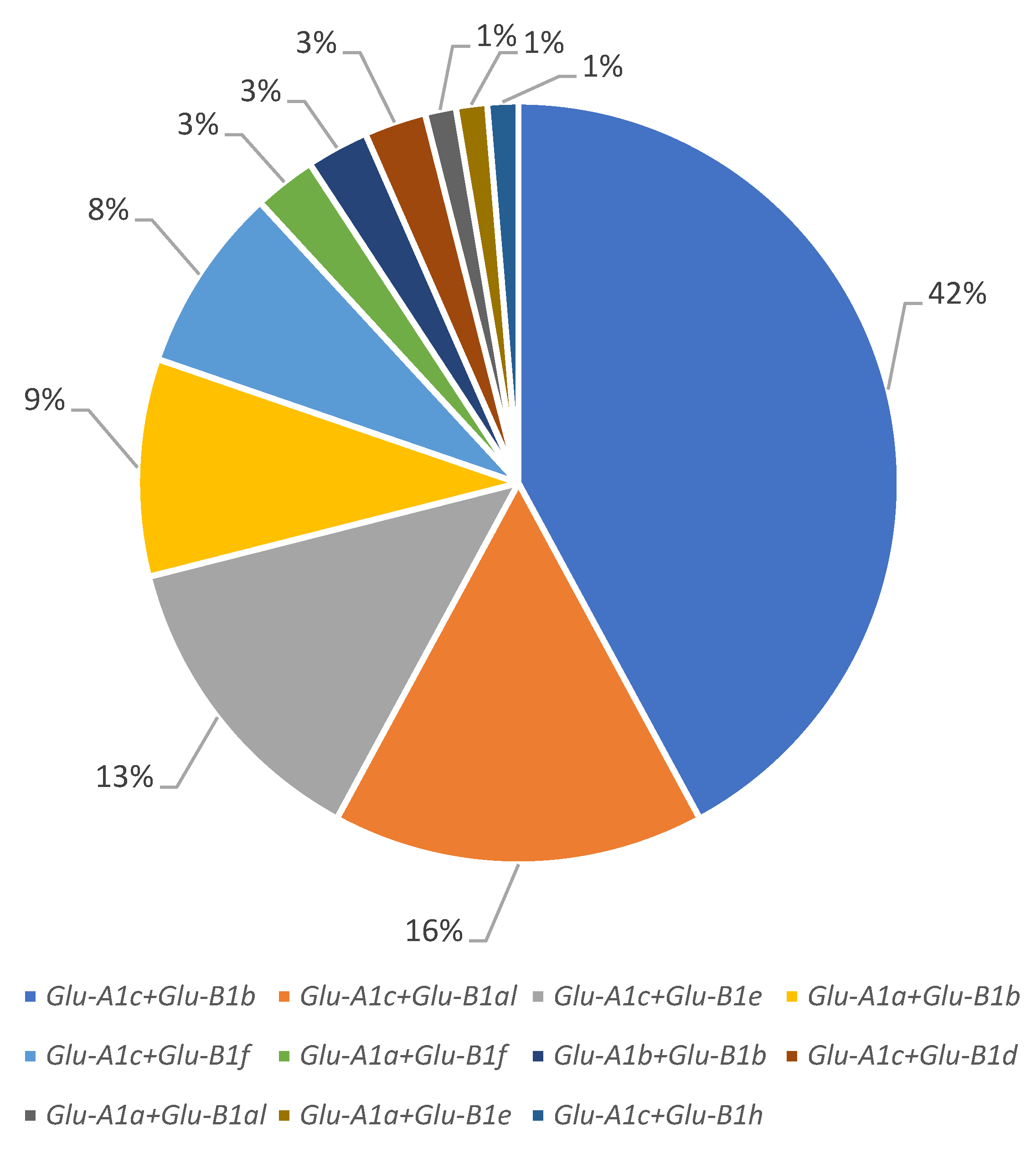{kind=link}
{kind=link}
{kind=link}
| Accession | Glu-A1 | Glu-B1 | GPE | Accession | Glu-A1 | Glu-B1 | GPE | ||
|---|---|---|---|---|---|---|---|---|---|
| 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | ||||||
| Afina | Glu-A1c | Glu-B1b | 4.5 | 4 | MV Pennedur | Glu-A1c | Glu-B1h | 4.5 | 4.5 |
| Agat Donskoy | Glu-A1c | Glu-B1b | 4 | 4 | MVTD 12-97 | Glu-A1c | Glu-B1al | 4.5 | 4 |
| Akveduk | Glu-A1c | Glu-B1b | 3.5 | 4 | MVTD 12-99 | Glu-A1c | Glu-B1e | 4.5 | 4.5 |
| Alena | Glu-A1c | Glu-B1b | 4.5 | 4.5 | MVTD 15-98 | Glu-A1c | Glu-B1e | 4.5 | 4.5 |
| Altana | Glu-A1c | Glu-B1b | 4 | 4 | Oniks | Glu-A1a | Glu-B1b | 4.5 | 4 |
| AMA 11855t85-90 | Glu-A1c | Glu-B1al | 3.5 | 3.5 | Pandur | Glu-A1c | Glu-B1b | 4 | 4.5 |
| AMA 15908ds1122-07 | Glu-A1c | Glu-B1f | 3.5 | 3.5 | Perlina odesskaya | Glu-A1c | Glu-B1e | 4.5 | 3.5 |
| Amazonka | Glu-A1c | Glu-B1f | 4 | 3.5 | Pributkova | Glu-A1c | Glu-B1b | 3.5 | 3.5 |
| Andromeda | Glu-A1c | Glu-B1b | 3.5 | 4 | Prikumchanka | Glu-A1c | Glu-B1b | 4 | 3.9 |
| Bagryanitsa (Italikum) | Glu-A1c | Glu-B1b | 4.5 | 3.5 | Rodur | Glu-A1c | Glu-B1al | 3.5 | 4 |
| Belgorodskaya yantarnaya | Glu-A1c | Glu-B1b | 3.5 | 4 | Rumyniya DF-00091-14-002 | Glu-A1c | Glu-B1al | 4 | 3.8 |
| Burshtyn | Glu-A1c | Glu-B1e | 4.5 | 4 | Rumyniya DF-01058-1-101 | Glu-A1c | Glu-B1al | 3.5 | 4 |
| Condur | Glu-A1c | Glu-B1al | 4 | 3 | Shulyndinka | Glu-A1c | Glu-B1b | 3.5 | 4 |
| DF-104/85 | Glu-A1c | Glu-B1b | 4.5 | 4 | Sin’ora | Glu-A1c | Glu-B1f | 4 | 3.8 |
| Diona | Glu-A1a | Glu-B1b | 4.5 | 4 | Stepnoy yantar’ | Glu-A1c | Glu-B1al | 3.5 | 3.5 |
| Dnepryana | Glu-A1c | Glu-B1b | 4 | 4 | Temnyy yantar’ | Glu-A1c | Glu-B1b | 3.5 | 4.5 |
| Dobryana | Glu-A1c | Glu-B1e | 4 | 4.5 | Teyya | Glu-A1c | Glu-B1e | 4.5 | 4.5 |
| Donchanka | Glu-A1a | Glu-B1e | 4 | 4 | Uniya | Glu-A1c | Glu-B1e | 4 | 4 |
| Elidur | Glu-A1c | Glu-B1al | 4 | 4 | Uslada | Glu-A1c | Glu-B1b | 4 | 3.9 |
| Eyrena | Glu-A1a | Glu-B1b | 4 | 3.5 | Wintergold | Glu-A1c | Glu-B1b | 3.5 | 4.5 |
| Khar’kovskaya 32 | Glu-A1a | Glu-B1b | 3.5 | 4 | Yagut | Glu-A1c | Glu-B1al | 4 | 4.3 |
| Kiprida | Glu-A1c | Glu-B1b | 4.5 | 3.5 | Yakhont (760/10) | Glu-A1a | Glu-B1f | 4 | 4 |
| Kontinent | Glu-A1c | Glu-B1b | 4 | 4.5 | Yubilyarka (330/10) | Glu-A1a | Glu-B1b | 4 | 3.9 |
| Kordon | Glu-A1c | Glu-B1f | 4 | 4 | Zhivitsa | Glu-A1c | Glu-B1e | 4 | 4 |
| Koshelevskaya | Glu-A1c | Glu-B1b | 3.5 | 4.5 | Zolotko | Glu-A1c | Glu-B1b | 4.5 | 4 |
| Kristall 2 | Glu-A1b | Glu-B1b | 4.5 | 4 | Zoloto Dona | Glu-A1c | Glu-B1b | 4 | 4.4 |
| Kristella | Glu-A1a | Glu-B1al | 4 | 4 | 533/14 | Glu-A1c | Glu-B1e | 4 | 4 |
| Krucha | Glu-A1c | Glu-B1b | 4 | 4 | 589/13 | Glu-A1a | Glu-B1b | 4 | 4.5 |
| Krupinka | Glu-A1c | Glu-B1b | 4 | 4 | 655/13 | Glu-A1c | Glu-B1b | 4 | 4 |
| Kurant | Glu-A1a | Glu-B1b | 4 | 3.9 | 737/11 | Glu-A1a | Glu-B1f | 4 | 4.5 |
| Laska | Glu-A1c | Glu-B1e | 4 | 3.5 | 796/11 | Glu-A1c | Glu-B1b | 4 | 4.5 |
| Lazurit | Glu-A1c | Glu-B1b | 4 | 3.5 | 21545t | Glu-A1c | Glu-B1f | 4 | 4 |
| Leukurum 21 | Glu-A1c | Glu-B1b | 4.5 | 4 | 3336h6-13-162 | Glu-A1c | Glu-B1b | 4 | 4 |
| Leukurum 36 | Glu-A1c | Glu-B1b | 4 | 4 | 3561h28 | Glu-A1b | Glu-B1b | 4 | 4 |
| Makar | Glu-A1c | Glu-B1b | 3.5 | 4.5 | 3591h96 | Glu-A1c | Glu-B1b | 4 | 4.5 |
| Martondur 1 | Glu-A1c | Glu-B1al | 4 | 3.8 | 3622h46 | Glu-A1c | Glu-B1d | 4 | 4.5 |
| MV Hundur | Glu-A1c | Glu-B1al | 4.5 | 4.5 | 3680h62 | Glu-A1c | Glu-B1d | 4.5 | 4 |
| MV Makaroni | Glu-A1c | Glu-B1al | 4.5 | 4.5 | 3796h33 | Glu-A1c | Glu-B1f | 3.5 | 3.8 |
| Locus/ Protein | Allele/ Subunit | Number of Accessions | Volume Increase Index | Breaking Strength | General Pasta Estimation | Gluten Index, % | SDS | Gluten Content, % | Protein Content, % | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | 2019–2020 | 2020–2021 | |||
| Glu-A1 | Glu-A1a | 11 | 3.88 | 4.02 | 804 | 788 | 4.05 | 4.03 | 77.7 ᵃ | 93.8 | 39.7 | 34.4 | 22.4 | 22.4 | 13.6 | 12.9 |
| Glu-A1b | 2 | 3.55 | 3.95 | 865 | 871 | 4.25 | 4.00 | 47.2 aᵇ | 93.8 | 41.1 | 39.8 | 25.0 | 24.9 | 14.6 | 13.5 | |
| Glu-A1c | 63 | 3.89 | 3.97 | 818 | 820 | 4.02 | 4.03 | 51.6 ᵇ | 83.3 | 42.0 | 35.1 | 23.2 | 22.4 | 13.7 | 13.0 | |
| Glu-A1c | Glu-A1a + Glu-A1b | 13 | 3.83 | 4.01 | 813 | 801 | 4.08 | 4.02 | 73.9 a | 93.8 | 39.9 ᵇ | 35.3 | 22.8 | 22.7 | 13.7 | 13.0 |
| Glu-A1c | 63 | 3.89 | 3.97 | 818 | 820 | 4.02 | 4.03 | 51.6 b | 83.3 | 42.0 ᵃ | 35.1 | 23.2 | 22.4 | 13.7 | 13.0 | |
| By | By16 | 8 | 3.97 ᵃ | 4.04 | 840 | 812 | 3.88 | 3.89 | 67.9 | 92.4 | 43.4 | 38.2 ᵃ | 23.7 | 24.7 ᵃ | 14.1 | 13.6 ᵃ |
| By20 | 11 | 3.67 ᵇ | 3.95 | 802 | 813 | 4.23 | 4.09 | 47.5 | 63.5 | 42.8 | 33.7 ᵇ | 23.2 | 22.2 ᵇ | 13.6 | 13.0 ᵃᵇ | |
| By8 | 56 | 3.92 ᵃ | 3.98 | 816 | 817 | 4.00 | 4.03 | 55.5 | 87.3 | 41.2 | 34.9 ᵇ | 23.0 | 22.1 ᵇ | 13.6 | 12.9 ᵇ | |
| Glu-B1 | Glu-B1al | 13 | 3.95 a | 4.02 | 818 | 824 | 3.96 | 3.92 | 63.5 | 95.3 ᵃ | 41.1 | 36.1 | 23.6 | 23.2 ᵃᵇ | 13.9 | 13.4 ᵃ |
| Glu-B1b | 41 | 3.91 a | 3.97 | 815 | 814 | 4.00 | 4.06 | 53.3 | 82.2 aᵇ | 41.3 | 34.6 | 22.9 | 21.8 ᵇ | 13.5 | 12.7 ᵇ | |
| Glu-B1d | 2 | 3.85 ᵃᵇ | 3.90 | 825 | 832 | 4.25 | 4.25 | 45.4 | 95.8 ᵃᵇ | 41.3 | 33.8 | 20.8 | 22.5 ᵃᵇ | 13.1 | 12.9 ᵃᵇ | |
| Glu-B1e | 11 | 3.67 b | 3.95 | 802 | 813 | 4.23 | 4.09 | 47.5 | 63.5 ᵇ | 42.8 | 33.7 | 23.2 | 22.2 aᵇ | 13.6 | 13.0 ᵃᵇ | |
| Glu-B1f | 8 | 3.97 a | 4.04 | 840 | 812 | 3.88 | 3.89 | 67.9 | 92.4 ᵃᵇ | 43.4 | 38.2 | 23.7 | 24.7 ᵃ | 14.1 | 13.6 ᵃ | |
| Glu-A1 + Glu-B1 | Glu-A1a Glu-B1b | 7 | 3.89 ᵃᵇ | 4.07 | 819 | 783 | 4.07 | 3.97 | 73.7 ᵃᵇ | 82.8 ᵃᵇ | 39.2 | 34.4 ᵃᵇ | 22.2 | 22.0 ᵃᵇ | 13.4 | 12.7 ab |
| Glu-A1a Glu-B1f | 2 | 3.85 ᵃᵇ | 3.80 | 782 | 804 | 4.00 | 4.25 | 89.2 ᵃ | 99.2 a | 39.4 | 34.0 ᵃᵇ | 23.6 | 23.2 ᵃᵇ | 14.1 | 12.9 ᵃᵇ | |
| Glu-A1b Glu-B1b | 2 | 3.55 ᵃᵇ | 3.95 | 865 | 871 | 4.25 | 4.00 | 47.2 aᵇᶜ | 93.8 ᵃᵇ | 41.1 | 39.8 ᵃᵇ | 25.0 | 24.9 ᵃᵇ | 14.6 | 13.5 ᵃᵇ | |
| Glu-A1c Glu-B1al | 12 | 3.96 a | 4.02 | 822 | 830 | 3.96 | 3.91 | 60.0 aᵇᶜ | 94.7 a | 41.1 | 36.1 ᵃᵇ | 23.8 | 23.2 ᵃᵇ | 13.9 | 13.5 a | |
| Glu-A1c Glu-B1b | 32 | 3.94 a | 3.94 | 810 | 817 | 3.97 | 4.08 | 48.8 ᶜ | 80.9 ᵃᵇ | 41.7 | 34.3 ᵇ | 22.9 | 21.5 ᵇ | 13.5 | 12.6 b | |
| Glu-A1c Glu-B1d | 2 | 3.85 ᵃᵇ | 3.90 | 825 | 832 | 4.25 | 4.25 | 45.4 aᵇᶜ | 95.8 ᵃᵇ | 41.3 | 33.8 ᵃᵇ | 20.8 | 22.5 ᵃᵇ | 13.1 | 12.9 ᵃᵇ | |
| Glu-A1c Glu-B1e | 10 | 3.65 b | 3.93 | 806 | 812 | 4.25 | 4.10 | 47.4 bᶜ | 53.7 b | 42.8 | 33.6 ᵇ | 23.3 | 22.2 aᵇ | 13.6 | 13.0 ᵃᵇ | |
| Glu-A1c Glu-B1f | 6 | 4.02 a | 4.12 | 860 | 815 | 3.83 | 3.77 | 57.3 ᵃᵇc | 84.6 ᵃᵇ | 44.8 | 39.6 ᵃ | 23.7 | 25.2 a | 14.1 | 13.9 a | |
| QC | 3.0–3.5 (LQ) | 16/12 | 4.12 a | 4.30 a | 843 | 787 | 3.50 ᶜ | 3.46 ᶜ | 42.6 ᵇ | 80.9 | 40.5 | 33.9 | 22.4 ᵇ | 22.1 | 13.3 ᵇ | 12.9 |
| 3.8–4.1 (MQ) | 40/43 | 3.93 ᵇ | 4.02 ᵇ | 812 | 824 | 4.00 ᵇ | 3.97 ᵇ | 64.0 ᵃ | 87.1 | 42.3 | 35.1 | 22.9 ᵇ | 22.3 | 13.7 ᵃ | 13.0 | |
| 4.3–4.5 (HQ) | 20/21 | 3.60 c | 3.70 c | 809 | 821 | 4.50 ᵃ | 4.49 ᵃ | 49.0 ᵇ | 85.6 | 41.4 | 35.9 | 24.1 ᵃ | 22.9 | 13.9 ᵃ | 13.1 | |
| Accession | 2019–2020 | 2020–2021 | ||||||
|---|---|---|---|---|---|---|---|---|
| Gluten Index, % | Gluten Content, % | General Pasta Estimation | Grain Yield, kg per 4.5 m2 Plot | Gluten Index, % | Gluten Content, % | General Pasta Estimation | Grain Yield, kg per 4.5 m2 Plot | |
| Krucha | 56.4 | 22.1 | 4.0 | 4.1 | 79.1 | 22.9 | 4.0 | 3.2 |
| Krupinka | 69.2 | 21.7 | 4.0 | 3.9 | 87.8 | 22.9 | 4.0 | 3.3 |
| 737/11 | 90.2 | 25.2 | 4.0 | 4.6 | 99.4 | 22.9 | 4.5 | 3.5 |
| 21545t | 80.0 | 26.6 | 4.0 | 3.1 | 92.3 | 30.5 | 4.0 | 2.4 |
| 589/13 | 75.7 | 25.5 | 4.0 | 4.6 | 99.7 | 25.4 | 4.5 | 2.9 |
| 3336h6-13-162 | 53.7 | 26.6 | 4.0 | 3.7 | 99.0 | 27.8 | 4.0 | 3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kroupina, A.Y.; Yanovsky, A.S.; Korobkova, V.A.; Bespalova, L.A.; Arkhipov, A.V.; Bukreeva, G.I.; Voropaeva, A.D.; Kroupin, P.Y.; Litvinov, D.Y.; Mudrova, A.A.; et al. Allelic Variation of Glu-A1 and Glu-B1 Genes in Winter Durum Wheat and Its Effect on Quality Parameters. Foods 2023, 12, 1436. https://doi.org/10.3390/foods12071436
Kroupina AY, Yanovsky AS, Korobkova VA, Bespalova LA, Arkhipov AV, Bukreeva GI, Voropaeva AD, Kroupin PY, Litvinov DY, Mudrova AA, et al. Allelic Variation of Glu-A1 and Glu-B1 Genes in Winter Durum Wheat and Its Effect on Quality Parameters. Foods. 2023; 12(7):1436. https://doi.org/10.3390/foods12071436
Chicago/Turabian StyleKroupina, Aleksandra Yu., Aleksey S. Yanovsky, Varvara A. Korobkova, Ludmila A. Bespalova, Andrey V. Arkhipov, Galina I. Bukreeva, Anastasiya D. Voropaeva, Pavel Yu. Kroupin, Dmitry Y. Litvinov, Aleksandra A. Mudrova, and et al. 2023. "Allelic Variation of Glu-A1 and Glu-B1 Genes in Winter Durum Wheat and Its Effect on Quality Parameters" Foods 12, no. 7: 1436. https://doi.org/10.3390/foods12071436