
iTRAQ-Based Proteomic Analyses of Regulation of Isothiocyanate and Endogenous Selenium Metabolism in Broccoli Sprouts by Exogenous Sodium Selenite

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Experimental Design
2.2. Determination of Sprout Length, Fresh Weight, Malondialdehyde Content, and Total Phenolics Content
2.3. Determination of ITCs Content, Myrosinase Activity, Glucosinolates Content, and Sulforaphane Content
2.4. Determination of Inorganic Selenium and Organic Selenium Content
2.5. RNA Extraction and Quantitative Real-Time PCR Analysis
2.6. Protein Extraction, Digestion, and iTRAQ Labeling
2.7. LC-MS/MS and Data Analysis
2.8. Statistical Analyses
3. Results
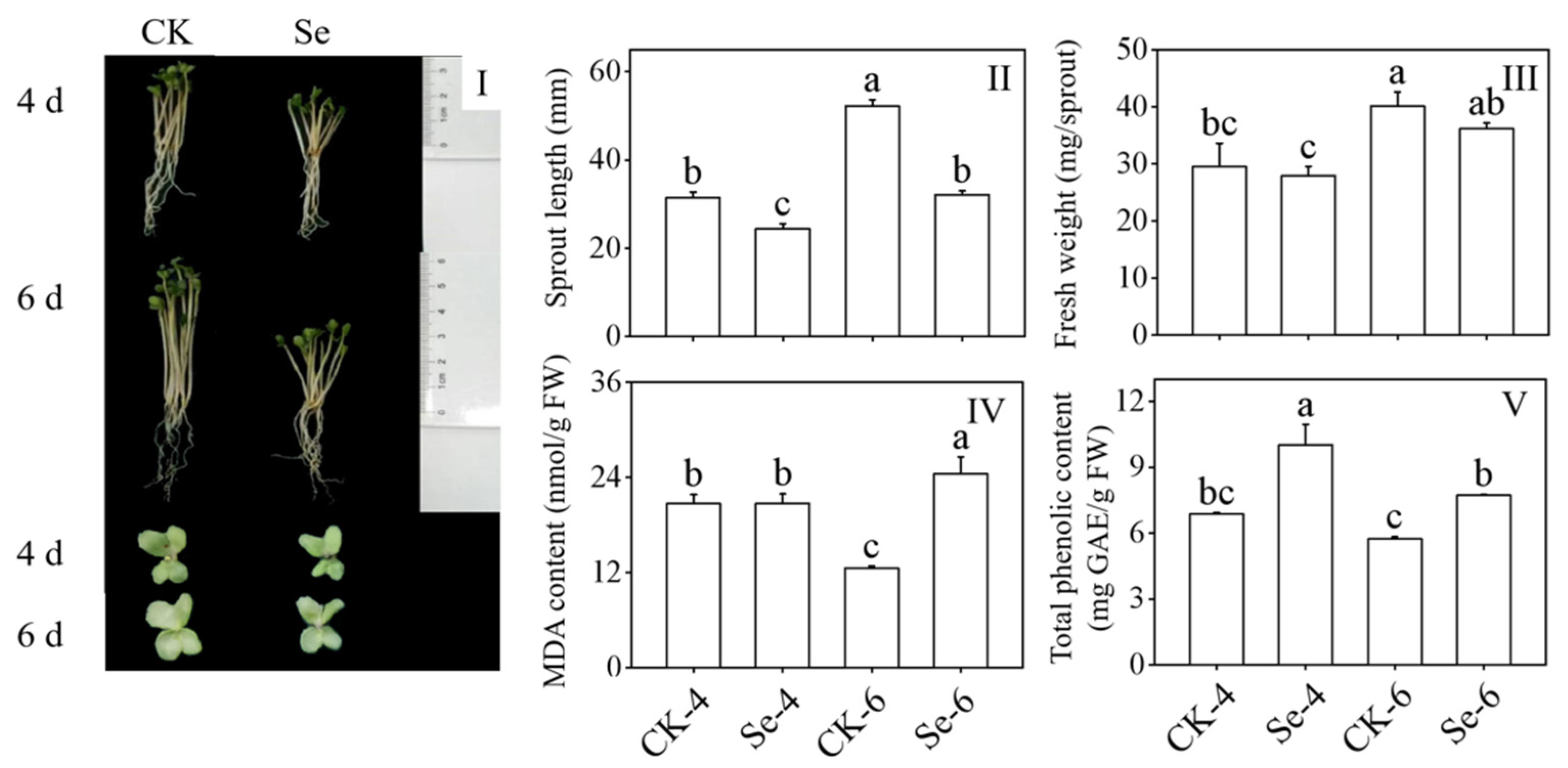
3.1. Effect of Selenium on Growth Performance, Sprout Length, Fresh Weight, Malondialdehyde Content, and Total Phenolic Content of Broccoli Sprouts
3.2. Effect of Selenium on ITCs Content, Myrosinase Activity, Glucosinolates Content, and Sulforaphane Content of Broccoli Sprouts
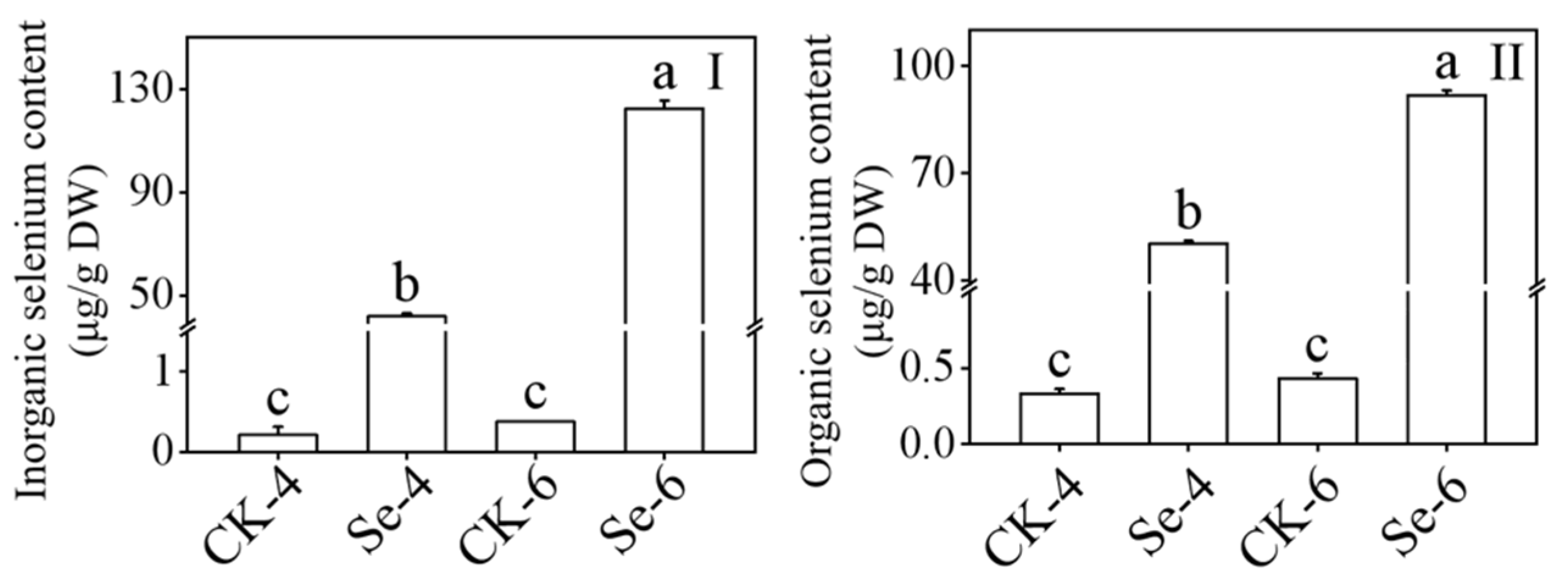
3.3. Effect of Selenium on Inorganic Selenium Content and Organic Selenium Content of Broccoli Sprouts
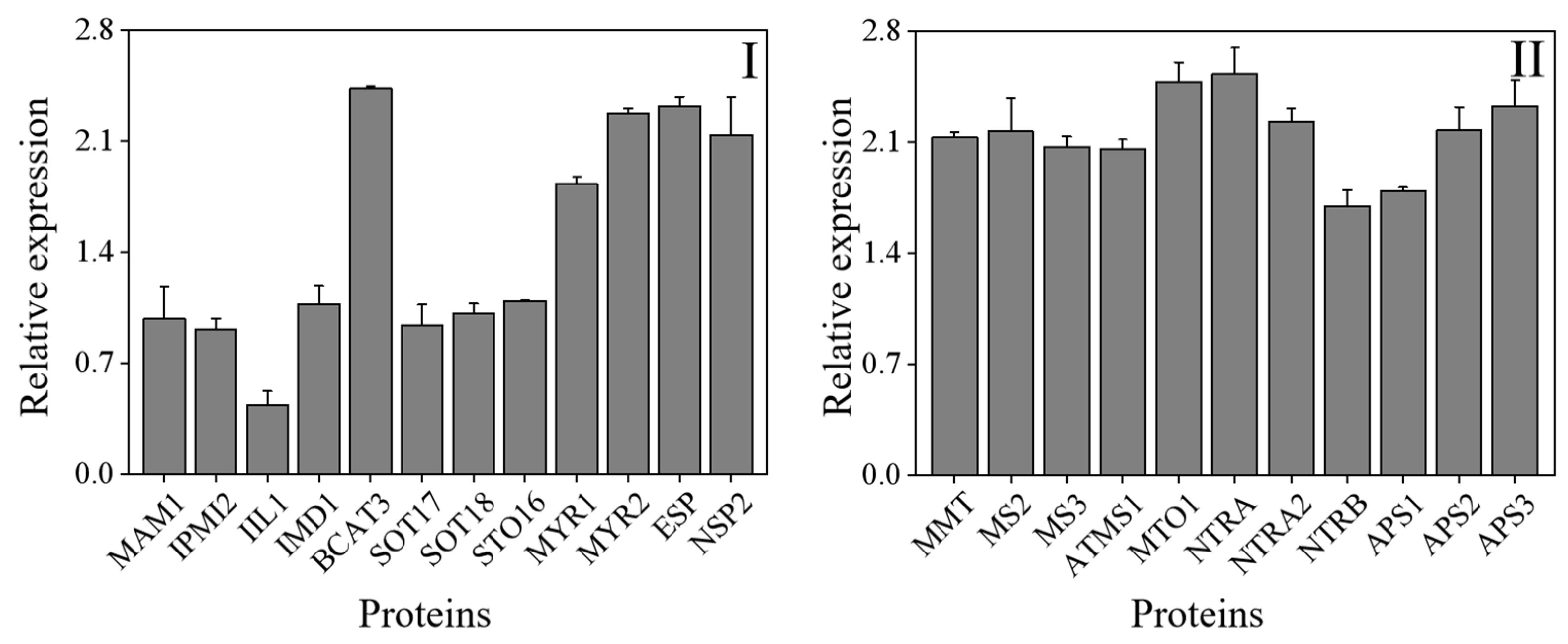
3.4. Changes in Gene Expression of ITCs and Selenium Metabolism Key Enzyme in Broccoli Sprouts
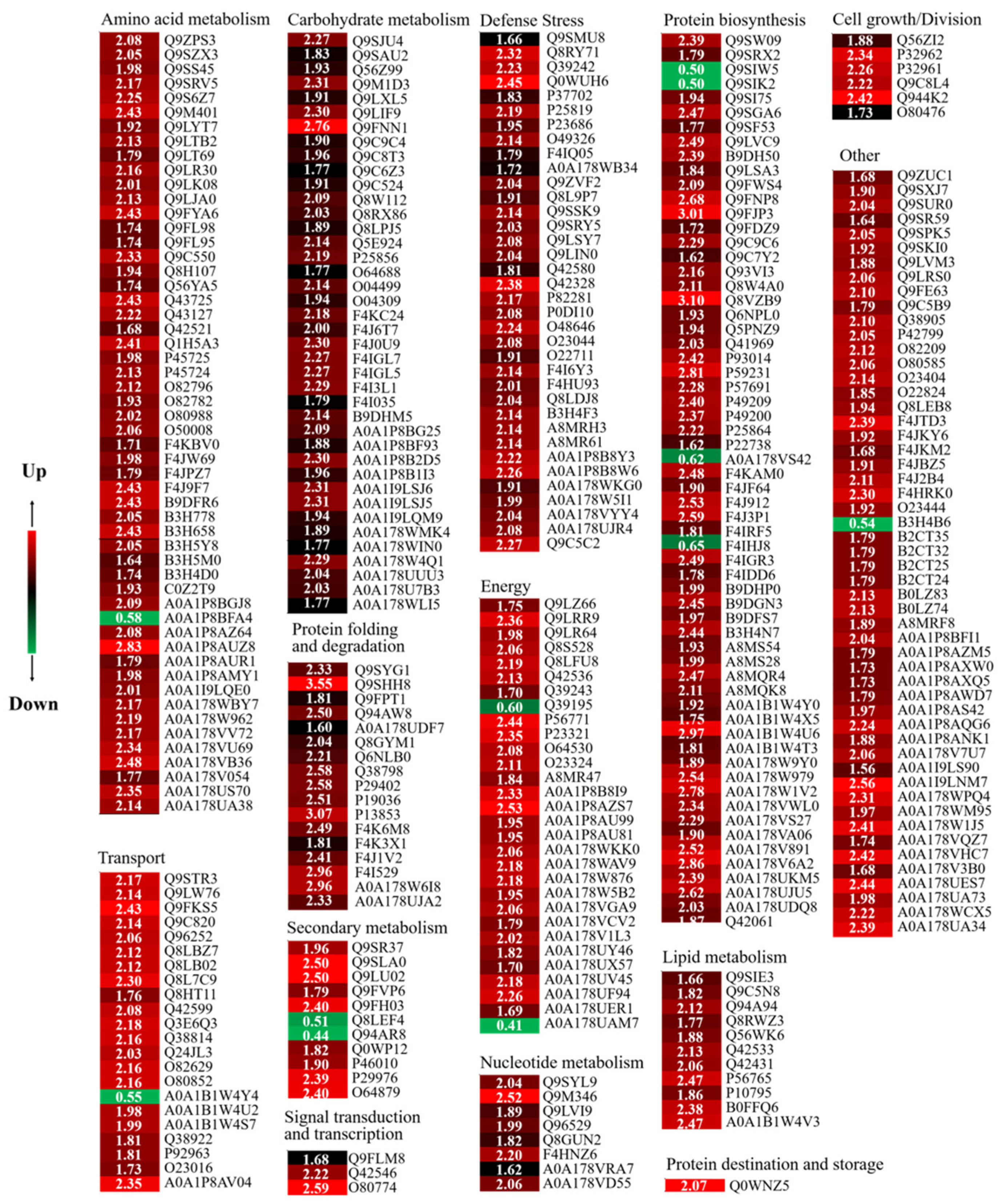
3.5. iTRAQ Analysis and Identification of Differentially Abundant Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Sun, Y.; Liu, H.; Song, S. The increase of antioxidant capacity of broccoli sprouts subject to slightly acidic electrolyzed water. Food Bio. 2022, 49, 101856. [Google Scholar] [CrossRef]
- Matusheski, N.V.; Juvik, J.A.; Jeffery, E.H. Heating decreases epithiospecifier protein activity and increases sulforaphane formation in broccoli. Phytochemistry 2004, 65, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Vanduchova, A.; Anzenbacher, P.; Anzenbacherova, E. Isothiocyanate from Broccoli, Sulforaphane, and Its Properties. J. Med. Food 2019, 22, 121–126. [Google Scholar] [CrossRef]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: Metabolism and excretion in humans. Cancer Epidemiol. Biomark. Prev. 2001, 10, 501–508. [Google Scholar]
- Jiao, D.; Yu, M.C.; Hankin, J.H.; Low, S.-H.; Chung, F.-L. Total isothiocyanate contents in cooked vegetables frequently consumed in Singapore. J. Agric. Food Chem. 1998, 46, 1055–1058. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, X.; Zhao, S.; Ma, C.; Cui, J.; Zheng, Y. Sulforaphane protects against cardiovascular disease via Nrf2 activation. Oxid. Med. Cell Longev. 2015, 2015, 407580. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Fahey, J.W.; Holtzclaw, W.D.; Wehage, S.L.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Sulforaphane bioavailability from glucoraphanin-rich broccoli: Control by active endogenous myrosinase. PLoS ONE 2015, 10, e0140963. [Google Scholar] [CrossRef]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Zoumpourlis, V.; Amery, T.; Galanis, A.; Pappa, A.; et al. The role of isothiocyanates as cancer chemo-preventive, chemo-therapeutic and anti-melanoma agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef]
- Burčul, F.; Generalić Mekinić, I.; Radan, M.; Rollin, P.; Blažević, I. Isothiocyanates: Cholinesterase inhibiting, antioxidant, and anti-inflammatory activity. J. Enzym. Inhib. Med. Chem. 2018, 33, 577–582. [Google Scholar] [CrossRef]
- Mohanty, S.; Sahoo, A.K.; Konkimalla, V.B.; Pal, A.; Si, S.C. Naringin in combination with isothiocyanates as liposomal formulations potentiates the anti-inflammatory activity in different acute and chronic animal models of rheumatoid arthritis. ACS Omega 2020, 5, 28319–28332. [Google Scholar] [CrossRef]
- Nieto, J.A.; Hellín, P.; Pérez, B.; Viadel, B.; Alapont, A.; Agudelo, A. Fresh Brassicaceae sprouting broccoli (Bimi®) glucosinolates profile characterization and bioaccessibility through an in vitro dynamic digestion study. J. Food Compos. Anal. 2023, 115, 104941. [Google Scholar] [CrossRef]
- Guo, Q.; Guo, L.; Wang, Z.; Zhuang, Y.; Gu, Z. Response surface optimization and identification of isothiocyanates produced from broccoli sprouts. Food Chem. 2013, 141, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, Y.; Cheng, C.; Yang, Z.; Luo, Z.; Fang, W. iTRAQ-based proteomic and physiological analyses of broccoli sprouts in response to exogenous melatonin with ZnSO4 stress. RSC Adv. 2021, 11, 12336–12347. [Google Scholar] [CrossRef]
- Stuss, M.; Michalska-Kasiczak, M.; Sewerynek, E. The role of selenium in thyroid gland pathophysiology. Endokrynol. Pol. 2017, 68, 440–465. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Sarkar, S.; Ahmad, Z.; Vemuri, H.; Garai, S.; Mondal, M.; Bhatt, R.; et al. Selenium biofortification: Roles, mechanisms, responses and prospects. Molecules 2021, 26, 881. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, J.; Huang, H.; Mao, S.; Wu, Q.; Huang, K. Exogenous selenium treatment promotes glucosinolate and glucoraphanin accumulation in broccoli by activating their biosynthesis and transport pathways. Appl. Sci. 2022, 12, 4101. [Google Scholar] [CrossRef]
- Tian, M.; Xu, X.; Liu, Y.; Xie, L.; Pan, S. Effect of Se treatment on glucosinolate metabolism and health-promoting compounds in the broccoli sprouts of three cultivars. Food Chem. 2016, 190, 374–380. [Google Scholar] [CrossRef]
- Tian, M.; Xu, X.; Liu, F.; Fan, X.; Pan, S. Untargeted metabolomics reveals predominant alterations in primary metabolites of broccoli sprouts in response to pre-harvest selenium treatment. Food Res. Int. 2018, 111, 205–211. [Google Scholar] [CrossRef]
- Zhuang, L.; Xu, K.; Zhu, Y.; Wang, F.; Xiao, J.; Guo, L. Calcium affects glucoraphanin metabolism in broccoli sprouts under ZnSO4 stress. Food Chem. 2021, 334, 127520. [Google Scholar] [CrossRef]
- Mencin, M.; Jamnik, P.; Mikulič Petkovšek, M.; Veberič, R.; Terpinc, P. Enzymatic treatments of raw, germinated and fermented spelt (Triticum spelta L.) seeds improve the accessibility and antioxidant activity of their phenolics. LWT 2022, 169, 114046. [Google Scholar] [CrossRef]
- Burow, M.; Losansky, A.; Muller, R.; Plock, A.; Kliebenstein, D.J.; Wittstock, U. The genetic basis of constitutive and herbivore-induced ESP-independent nitrile formation in Arabidopsis. Plant Physiol. 2009, 149, 561–574. [Google Scholar] [CrossRef]
- Guo, L.; Yang, R.; Wang, Z.; Guo, Q.; Gu, Z. Glucoraphanin, sulforaphane and myrosinase activity in germinating broccoli sprouts as affected by growth temperature and plant organs. J. Funct. Foods 2014, 9, 70–77. [Google Scholar] [CrossRef]
- Guo, L.; Gu, Z.; Jin, X.; Yang, R. iTRAQ—Based proteomic and physiological analyses of broccoli sprouts in response to the stresses of heat, hypoxia and heat plus hypoxia. Plant Soil 2016, 414, 355–377. [Google Scholar] [CrossRef]
- Sun, M.; Liu, G.; Wu, Q. Speciation of organic and inorganic selenium in selenium-enriched rice by graphite furnace atomic absorption spectrometry after cloud point extraction. Food Chem. 2013, 141, 66–71. [Google Scholar] [CrossRef]
- Lyi, S.M.; Heller, L.I.; Rutzke, M.; Welch, R.M.; Kochian, L.V.; Li, L. Molecular and biochemical characterization of the selenocysteine Se-methyltransferase gene and Se-methylselenocysteine synthesis in broccoli. Plant Physiol. 2005, 138, 409–420. [Google Scholar] [CrossRef]
- Cheng, C.; Liu, Y.; Fang, W.; Tao, J.; Yang, Z.; Yin, Y. iTRAQ-based proteomic and physiological analyses of mustard sprouts in response to heat stress. RSC Adv. 2020, 10, 6052–6062. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Qi, F.; Gao, L.; Rao, S.; Yang, Z.; Fang, W. iTRAQ-based quantitative proteomic analysis of dark-germinated soybeans in response to salt stress. RSC Adv. 2018, 8, 17905–17913. [Google Scholar] [CrossRef]
- Jin, X.; Yang, R.; Guo, L.; Wang, X.; Yan, X.; Gu, Z. iTRAQ analysis of low-phytate mung bean sprouts treated with sodium citrate, sodium acetate and sodium tartrate. Food Chem. 2017, 218, 285–293. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Guo, L.; Jin, X.; Shen, C.; Zhou, Y.; Gu, Z. Enhancement of glucosinolate and sulforaphane formation of broccoli sprouts by zinc sulphate via its stress effect. J. Funct. Foods 2015, 13, 345–349. [Google Scholar] [CrossRef]
- Ríos, J.J.; Rosales, M.A.; Blasco, B.; Cervilla, L.M.; Romero, L.; Ruiz, J.M. Biofortification of Se and induction of the antioxidant capacity in lettuce plants. Sci. Hortic. 2008, 116, 248–255. [Google Scholar] [CrossRef]
- Williams, D.J.; Critchley, C.; Pun, S.; Nottingham, S.; O’Hare, T.J. Epithiospecifier protein activity in broccoli: The link between terminal alkenyl glucosinolates and sulphoraphane nitrile. Phytochemistry 2008, 69, 2765–2773. [Google Scholar] [CrossRef] [PubMed]
- Gui, J.-Y.; Rao, S.; Huang, X.; Liu, X.; Cheng, S.; Xu, F. Interaction between selenium and essential micronutrient elements in plants: A systematic review. Sci. Total Environ. 2022, 853, 158673. [Google Scholar] [CrossRef]
- Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K.; Wu, Q. Effects of sulfur and selenium on glucosinolate biosynthesis in cabbage. Plant Mol. Biol. Rep. 2019, 38, 62–74. [Google Scholar] [CrossRef]
- McKenzie, M.J.; Chen, R.K.Y.; Leung, S.; Joshi, S.; Rippon, P.E.; Joyce, N.I.; McManus, M.T. Selenium treatment differentially affects sulfur metabolism in high and low glucosinolate producing cultivars of broccoli (Brassica oleracea L.). Plant Physiol. BioChem. 2017, 121, 176–186. [Google Scholar] [CrossRef]
- Avila, F.W.; Faquin, V.; Yang, Y.; Ramos, S.J.; Guilherme, L.R.; Thannhauser, T.W.; Li, L. Assessment of the anticancer compounds Se-methylselenocysteine and glucosinolates in Se-biofortified broccoli (Brassica oleracea L. var. italica) sprouts and florets. J. Agric. Food Chem. 2013, 61, 6216–6223. [Google Scholar] [CrossRef]
- Zuluaga, D.L.; Graham, N.S.; Klinder, A.; van Ommen Kloeke, A.E.E.; Marcotrigiano, A.R.; Wagstaff, C.; Verkerk, R.; Sonnante, G.; Aarts, M.G.M. Overexpression of the MYB29 transcription factor affects aliphatic glucosinolate synthesis in Brassica oleracea. Plant Mol. Biol. 2019, 101, 65–79. [Google Scholar] [CrossRef]
- El Mehdawi, A.F.; Jiang, Y.; Guignardi, Z.S.; Esmat, A.; Pilon, M.; Pilon-Smits, E.A.H.; Schiavon, M. Influence of sulfate supply on selenium uptake dynamics and expression of sulfate/selenate transporters in selenium hyperaccumulator and nonhyperaccumulator Brassicaceae. New Phytol. 2018, 217, 194–205. [Google Scholar] [CrossRef]
- Zhou, X.; Yuan, Y.; Yang, Y.; Rutzke, M.; Thannhauser, T.W.; Kochian, L.V.; Li, L. Involvement of a broccoli COQ5 methyltransferase in the production of volatile selenium compounds. Plant Physiol. 2009, 151, 528–540. [Google Scholar] [CrossRef]
- Ramos, S.J.; Yuan, Y.; Faquin, V.; Guilherme, L.R.; Li, L. Evaluation of genotypic variation of broccoli (Brassica oleracea var. italic) in response to selenium treatment. J. Agric. Food Chem. 2011, 59, 3657–3665. [Google Scholar] [CrossRef]
- Sonderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of glucosinolates—Gene discovery and beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.J.; Rutzke, M.A.; Hayes, R.J.; Faquin, V.; Guilherme, L.R.; Li, L. Selenium accumulation in lettuce germplasm. Planta 2011, 233, 649–660. [Google Scholar] [CrossRef] [PubMed]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Actin | CTGTTCCAATCTACGAGGGTTTCT | GCTCGGCTGTGGTGGTGAA |
| MYB28 | AGACTGCGATGGACTAACTACCTAAA | CCGACCACTTGTTTCCACGA |
| UGT74B1 | CAAAGACGATAAAGGCTACGGC | TCCCAAAGGAACCAAACGAA |
| ST5b | CCGACACTACCTTACCGAACCA | CGTGAGGAAAAGAGGCGATG |
| OX-1 | GTGGACATTATACCGAACCTTACG | TGTGGACTTCTTTGGCGACCT |
| MYR | AAGGTCATCAGGGAGAAGGGTG | TGTTTGGCAGGGTTCTTAGTGG |
| ESP-F | ACATTTGGGACCAGGGACG | TTTCCATACACGGTGGCAGTC |
| BoSultr1;1 | GATTCTGCTGCAAGTGACGA | ACGCGAATGATCAAGATTCC |
| BoSAT1;1 | ATATCCATCCAGCAGCGAAG | CTGTCTCCGCAAGCTTTACC |
| BoHMT1 | TTCAGGAATGCCTTGAAACC | TTAGCTTTTCCGTCCCACAC |
| BoSMT | GATCAACTGTACCCCTCCAAG | TCCCAACTCCTGTGTTTTCC |
| BoCOQ5-2 | AAGGAAAGACTCGTTGGGAAG | TCCTAAACGCAACATCACCC |
| Pathway ID | Pathway | Input Number | p-Value |
|---|---|---|---|
| Se/C | Se/C | ||
| ath00195 | Photosynthesis | 12 | 0.0007 |
| ath00250 | Alanine, aspartate and glutamate metabolism | 8 | 0.0061 |
| ath00260 | Glycine, serine and threonine metabolism | 10 | 0.0056 |
| ath00270 | Cysteine and methionine metabolism | 12 | 0.0057 |
| ath00290 | Valine, leucine and isoleucine biosynthesis | 5 | 0.0198 |
| ath00380 | Tryptophan metabolism | 8 | 0.0048 |
| ath00450 | Selenocompound metabolism | 10 | 2.82 × 10−8 |
| ath00460 | Cyanoamino acid metabolism | 10 | 0.0016 |
| ath00480 | Glutathione metabolism | 10 | 0.0273 |
| ath00620 | Pyruvate metabolism | 9 | 0.0395 |
| ath00630 | Glyoxylate and dicarboxylate metabolism | 14 | 2.69 × 10−5 |
| ath00710 | Carbon fixation in photosynthetic organisms | 11 | 0.0011 |
| ath00920 | Sulfur metabolism | 15 | 7.49 × 10−10 |
| ath00940 | Phenylpropanoid biosynthesis | 19 | 0.0003 |
| ath01100 | Metabolic pathways | 132 | 9.41 × 10−11 |
| ath01110 | Biosynthesis of secondary metabolites | 81 | 1.48 × 10−7 |
| ath01130 | Biosynthesis of antibiotics | 52 | 2.01 × 10−10 |
| ath01200 | Carbon metabolism | 45 | 1.23 × 10−14 |
| ath01210 | 2-Oxocarboxylic acid metabolism | 9 | 0.0202 |
| ath01230 | Biosynthesis of amino acids | 37 | 6.95 × 10−10 |
| ath03010 | Ribosome | 44 | 4.03 × 10−9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, X.; Cheng, Y.; Yang, Z.; Yang, J.; Fang, W.; Yin, Y. iTRAQ-Based Proteomic Analyses of Regulation of Isothiocyanate and Endogenous Selenium Metabolism in Broccoli Sprouts by Exogenous Sodium Selenite. Foods 2023, 12, 1397. https://doi.org/10.3390/foods12071397
Quan X, Cheng Y, Yang Z, Yang J, Fang W, Yin Y. iTRAQ-Based Proteomic Analyses of Regulation of Isothiocyanate and Endogenous Selenium Metabolism in Broccoli Sprouts by Exogenous Sodium Selenite. Foods. 2023; 12(7):1397. https://doi.org/10.3390/foods12071397
Chicago/Turabian StyleQuan, Xiaolan, Yuwei Cheng, Zhengfei Yang, Jia Yang, Weiming Fang, and Yongqi Yin. 2023. "iTRAQ-Based Proteomic Analyses of Regulation of Isothiocyanate and Endogenous Selenium Metabolism in Broccoli Sprouts by Exogenous Sodium Selenite" Foods 12, no. 7: 1397. https://doi.org/10.3390/foods12071397