
A Narrative Mini Review on Current Status of Hypoallergenic Wheat Development for IgE-Mediated Wheat Allergy, Wheat-Dependent Exercise-Induced Anaphylaxis
 , ,
, ,
Abstract
:1. Introduction
2. WDEIA and the Responsible Allergens
3. Approaches to Develop Hypoallergnic Wheat
3.1. Wheat Lines with a Reduced Allergenicity
3.2. Hypoallergenic wheat by Enzymic Degradation/Ion Exchanger Deamidation
3.3. Production of Hypoallergenic Wheat by Thioredoxin Treatment
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Cianferoni, A. Wheat allergy: Diagnosis and management. J. Asthma Allergy 2016, 9, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Sicherer, S.H.; Warren, C.M.; Dant, C.; Gupta, R.S.; Nadeau, K.C. Food Allergy from Infancy through Adulthood. J. Allergy Clin. Immunol. Pract. 2020, 8, 1854–1864. [Google Scholar] [CrossRef] [PubMed]
- Keet, C.A.; Matsui, E.C.; Dhillon, G.; Lenehan, P.; Paterakis, M.; Wood, R.A. The natural history of wheat allergy. Ann. Allergy Asthma Immunol. 2009, 102, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Kunie, K.; Matsuo, H. Food-dependent exercise-induced anaphylaxis. J. Dermatol. Sci. 2007, 47, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Ansley, L.; Bonini, M.; Delgado, L.; Del Giacco, S.; Du Toit, G.; Khaitov, M.; Kurowski, M.; Hull, J.H.; Moreira, A.; Robson-Ansley, P.J. Pathophysiological mechanisms of exercise-induced anaphylaxis: An EAACI position statement. Allergy 2015, 70, 1212–1221. [Google Scholar] [CrossRef]
- Scherf, K.A.; Brockow, K.; Biedermann, T.; Koehler, P.; Wieser, H. Wheat-dependent exercise-induced anaphylaxis. Clin. Exp. Allergy 2015, 46, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Chinuki, Y.; Fukutomi, Y.; Nakatani, E.; Yagami, A.; Matsunaga, K.; Oda, Y.; Fukunaga, A.; Adachi, A.; Hiragun, M.; et al. Long-term dynamics of omega-5 gliadin-specific IgE levels in patients with adult-onset wheat allergy. J. Allergy Clin. Immunol. Pract. 2019, 8, 1149–1151.e3. [Google Scholar] [CrossRef]
- Gupta, R.S.; Warren, C.M.; Smith, B.M.; Jiang, J.; Blumenstock, J.A.; Davis, M.M.; Schleimer, R.P.; Nadeau, K.C. Prevalence and Severity of Food Allergies among US Adults. JAMA Netw. Open 2019, 2, e185630. [Google Scholar] [CrossRef] [Green Version]
- Morita, E.; Chinuki, Y.; Takahashi, H.; Nabika, T.; Yamasaki, M.; Shiwaku, K. Prevalence of Wheat Allergy in Japanese Adults. Allergol. Int. 2012, 61, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Dhami, S.; Nurmatov, U.; Arasi, S.; Khan, T.; Asaria, M.; Zaman, H.; Agarwal, A.; Netuveli, G.; Roberts, G.; Pfaar, O.; et al. Allergen immunotherapy for allergic rhinoconjunctivitis: A systematic review and meta-analysis. Allergy 2017, 72, 1597–1631. [Google Scholar] [CrossRef] [Green Version]
- Babaie, D.; Ebisawa, M.; Soheili, H.; Ghasemi, R.; Zandieh, F.; Sahragard, M.; Seifi, H.; Fallahi, M.; Khoshmirsafa, M.; Darougar, S.; et al. Oral Wheat Immunotherapy: Long-Term Follow-Up in Children with Wheat Anaphylaxis. Int. Arch. Allergy Immunol. 2021, 183, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Węgrzyn, A.; Wood, R.A.; Nadeau, K.C.; Pongracic, J.A.; Henning, A.K.; Lindblad, R.W.; Beyer, K.; Sampson, H.A. Multicenter, randomized, double-blind, placebo-controlled clinical trial of vital wheat gluten oral immunotherapy. J. Allergy Clin. Immunol. 2019, 143, 651–661.e9. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Tatham, A.S.; Shewry, P.R. Allergy to wheat and related cereals. Clin. Exp. Allergy 2008, 38, 1712–1726. [Google Scholar] [CrossRef]
- Matsuo, H.; Yokooji, T.; Taogoshi, T. Common food allergens and their IgE-binding epitopes. Allergol. Int. 2015, 64, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Palosuo, K.; Alenius, H.; Varjonen, E.; Koivuluhta, M.; Mikkola, J.; Keskinen, H.; Kalkkinen, N.; Reunala, T. A novel wheat gliadin as a cause of exercise-induced anaphylaxis. J. Allergy Clin. Immunol. 1999, 103, 912–917. [Google Scholar] [CrossRef]
- Palosuo, K.; Varjonen, E.; Kekki, O.-M.; Klemola, T.; Kalkkinen, N.; Alenius, H.; Reunala, T. Wheat ω-5 gliadin is a major allergen in children with immediate allergy to ingested wheat. J. Allergy Clin. Immunol. 2001, 108, 634–638. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Matsuo, H.; Mihara, S.; Morimoto, K.; Savage, A.; Tatham, A. Fast ω-gliadin is a major allergen in wheat-dependent exercise-induced anaphylaxis. J. Dermatol. Sci. 2003, 33, 99–104. [Google Scholar] [CrossRef]
- Matsuo, H.; Morita, E.; Tatham, A.S.; Morimoto, K.; Horikawa, T.; Osuna, H.; Ikezawa, Z.; Kaneko, S.; Kohno, K.; Dekio, S. Identification of the IgE-binding Epitope in ω-5 Gliadin, a Major Allergen in Wheat-dependent Exercise-induced Anaphylaxis. J. Biol. Chem. 2004, 279, 12135–12140. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, H.; Kohno, K.; Niihara, H.; Morita, E. Specific IgE Determination to Epitope Peptides of ω-5 Gliadin and High Molecular Weight Glutenin Subunit Is a Useful Tool for Diagnosis of Wheat-Dependent Exercise-Induced Anaphylaxis. J. Immunol. 2005, 175, 8116–8122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, H.; Kohno, K.; Morita, E. Molecular cloning, recombinant expression and IgE-binding epitope of omega-5 gliadin, a major allergen in wheat-dependent exercise-induced anaphylaxis. FEBS J. 2005, 272, 4431–4438. [Google Scholar] [CrossRef] [PubMed]
- Battais, F.; Mothes, T.; Moneret-Vautrin, D.A.; Pineau, F.; Kanny, G.; Popineau, Y.; Bodinier, M.; Denery-Papini, S. Identification of IgE-binding epitopes on gliadins for patients with food allergy to wheat. Allergy 2005, 60, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Matsuo, H.; Chinuki, Y.; Takahashi, H.; Dahlström, J.; Tanaka, A. Food-Dependent Exercise-Induced Anaphylaxis—Importance of Omega-5 Gliadin and HMW-Glutenin as Causative Antigens for Wheat-Dependent Exercise-Induced Anaphylaxis. Allergol. Int. 2009, 58, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennard, L.; Thomas, I.; Rutkowski, K.; Azzu, V.; Yong, P.F.; Kasternow, B.; Hunter, H.; Cabdi, N.M.; Nakonechna, A.; Wagner, A. A Multicenter Evaluation of Diagnosis and Management of Omega-5 Gliadin Allergy (Also Known as Wheat-Dependent Exercise-Induced Anaphylaxis) in 132 Adults. J. Allergy Clin. Immunol. Pract. 2018, 6, 1892–1897. [Google Scholar] [CrossRef]
- Bouchez-Mahiout, I.; Snégaroff, J.; Tylichova, M.; Pecquet, C.; Branlard, G.; Laurière, M. Low Molecular Weight Glutenins in Wheat-Dependant, Exercise-Induced Anaphylaxis: Allergenicity and Antigenic Relationships with Omega 5-Gliadins. Int. Arch. Allergy Immunol. 2010, 153, 35–45. [Google Scholar] [CrossRef]
- Yokooji, T.; Kurihara, S.; Murakami, T.; Chinuki, Y.; Takahashi, H.; Morita, E.; Harada, S.; Ishii, K.; Hiragun, M.; Hide, M.; et al. Characterization of Causative Allergens for Wheat-Dependent Exercise-Induced Anaphylaxis Sensitized with Hydrolyzed Wheat Proteins in Facial Soap. Allergol. Int. 2013, 62, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Ogino, R.; Chinuki, Y.; Yokooji, T.; Takizawa, D.; Matsuo, H.; Morita, E. Identification of peroxidase-1 and beta-glucosidase as cross-reactive wheat allergens in grass pollen-related wheat allergy. Allergol. Int. 2020, 70, 215–222. [Google Scholar] [CrossRef]
- Lexhaller, B.; Tompos, C.; Scherf, K.A. Fundamental study on reactivities of gluten protein types from wheat, rye and barley with five sandwich ELISA test kits. Food Chem. 2017, 237, 320–330. [Google Scholar] [CrossRef]
- Palosuo, K.; Alenius, H.; Varjonen, E.; Kalkkinen, N.; Reunala, T. Rye γ-70 and γ-35 secalins and barley γ-3 hordein cross-react with ω-5 gliadin, a major allergen in wheat-dependent, exercise-induced anaphylaxis. Clin. Exp. Allergy 2001, 31, 466–473. [Google Scholar] [CrossRef]
- Snégaroff, J.; Bouchez-Mahiout, I.; Pecquet, C.; Branlard, G.; Laurière, M. Study of IgE Antigenic Relationships in Hypersensitivity to Hydrolyzed Wheat Proteins and Wheat-Dependent Exercise-Induced Anaphylaxis. Int. Arch. Allergy Immunol. 2006, 139, 201–208. [Google Scholar] [CrossRef]
- Fukutomi, Y.; Itagaki, Y.; Taniguchi, M.; Saito, A.; Yasueda, H.; Nakazawa, T.; Hasegawa, M.; Nakamura, H.; Akiyama, K. Rhinoconjunctival sensitization to hydrolyzed wheat protein in facial soap can induce wheat-dependent exercise-induced anaphylaxis. J. Allergy Clin. Immunol. 2011, 127, 531–533.e3. [Google Scholar] [CrossRef]
- Chinuki, Y.; Morita, E. Wheat-Dependent Exercise-Induced Anaphylaxis Sensitized with Hydrolyzed Wheat Protein in Soap. Allergol. Int. 2012, 61, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagami, A.; Aihara, M.; Ikezawa, Z.; Hide, M.; Kishikawa, R.; Morita, E.; Chinuki, Y.; Fukutomi, Y.; Urisu, A.; Fukushima, A.; et al. Outbreak of immediate-type hydrolyzed wheat protein allergy due to a facial soap in Japan. J. Allergy Clin. Immunol. 2017, 140, 879–881.e7. [Google Scholar] [CrossRef] [Green Version]
- Denery-Papini, S.; Bodinier, M.; Larre, C.; Brossard, C.; Pineau, F.; Triballeau, S.; Pietri, M.; Battais, F.; Mothes, T.; Paty, E.; et al. Allergy to deamidated gluten in patients tolerant to wheat: Specific epitopes linked to deamidation. Allergy 2012, 67, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, N.; Nilsson, C.; Ekoff, H.; Wieser-Pahr, S.; Borres, M.P.; Valenta, R.; Hedlin, G.; Sjölander, S. Grass-Allergic Children Frequently Show Asymptomatic Low-Level IgE Co-Sensitization and Cross-Reactivity to Wheat. Int. Arch. Allergy Immunol. 2018, 177, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Denery-Papini, S.; Lauriére, M.; Branlard, G.; Morisset, M.; Pecquet, C.; Choudat, D.; Merlino, M.; Pineau, F.; Popineau, Y.; Boulenc, E.; et al. Influence of the Allelic Variants Encoded at the Gli-B1 Locus, Responsible for a Major Allergen of Wheat, on IgE Reactivity for Patients Suffering from Food Allergy to Wheat. J. Agric. Food Chem. 2007, 55, 799–805. [Google Scholar] [CrossRef]
- Gabler, A.M.; Gebhard, J.; Eberlein, B.; Biedermann, T.; Scherf, K.A.; Brockow, K. The basophil activation test differentiates between patients with wheat-dependent exercise-induced anaphylaxis and control subjects using gluten and isolated gluten protein types. Clin. Transl. Allergy 2021, 11, e12050. [Google Scholar] [CrossRef] [PubMed]
- Gabler, A.M.; Gebhard, J.; Norwig, M.-C.; Eberlein, B.; Biedermann, T.; Brockow, K.; Scherf, K.A. Basophil Activation to Gluten and Non-Gluten Proteins in Wheat-Dependent Exercise-Induced Anaphylaxis. Front. Allergy 2022, 3, 822554. [Google Scholar] [CrossRef]
- Lombardo, C.; Bolla, M.; Chignola, R.; Senna, G.; Rossin, G.; Caruso, B.; Tomelleri, C.; Cecconi, D.; Brandolini, A.; Zoccatelli, G. Study on the Immunoreactivity of Triticum monococcum (Einkorn) Wheat in Patients with Wheat-Dependent Exercise-Induced Anaphylaxis for the Production of Hypoallergenic Foods. J. Agric. Food Chem. 2015, 63, 8299–8306. [Google Scholar] [CrossRef]
- Waga, J.; Skoczowski, A. Development and characteristics of ω-gliadin-free wheat genotypes. Euphytica 2013, 195, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Skoczowski, A.; Obtułowicz, K.; Czarnobilska, E.; Dyga, W.; Mazur, M.; Stawoska, I.; Waga, J. Antibody reactivity in patients with IgE-mediated wheat allergy to various subunits and fractions of gluten and non-gluten proteins from ω-gliadin-free wheat genotypes. Ann. Agric. Environ. Med. 2017, 24, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Takahashi, H.; Endo, T.R.; Matsuo, H.; Shiwaku, K.; Morita, E. Characterization of a hypoallergenic wheat line lacking ω-5 gliadin. Allergol. Int. 2016, 65, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Yokooji, T.; Nouma, H.; Ogino, R.; Taogoshi, T.; Morita, E.; Matsuo, H. Quantification of the ω5- and γ-gliadin content in wheat flour and rat plasma with an enzyme-linked immunosorbent assay using antibodies specific to their IgE-binding epitopes. Allergol. Int. 2018, 68, 112–113. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Yokooji, T.; Ninomiya, N.; Taogoshi, T.; Morita, E.; Matsuo, H. Evaluation of the allergenicity of ω5-gliadin-deficient Hokushin wheat (1BS-18) in a wheat allergy rat model. Biochem. Biophys. Rep. 2019, 20, 100702. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Yokooji, T.; Kunimoto, K.; Inoguchi, K.; Ogino, R.; Taogoshi, T.; Morita, E.; Matsuo, H. Hypoallergenic Wheat Line (1BS-18H) Lacking ω5-Gliadin Induces Oral Tolerance to Wheat Gluten Proteins in a Rat Model of Wheat Allergy. Foods 2022, 11, 2181. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Kang, C.-S.; Beom, H.-R.; Jang, Y.-R.; Altenbach, S.B.; Lim, S.-H.; Kim, Y.-M.; Park, C.-S. Characterization of a wheat mutant missing low-molecular-weight glutenin subunits encoded by the B-genome. J. Cereal Sci. 2017, 73, 158–164. [Google Scholar] [CrossRef]
- Lee, J.; Kim, S.-R.; Park, J.H.; Park, K.-H.; Jeong, K.Y.; Lee, J.-H.; Kang, C.-S.; Kim, K.-H.; Park, J.-W. Evaluation of Allergenicity on a ω-5 Gliadin-Deficient Cultivar in Wheat-Dependent Exercise-Induced Anaphylaxis. Allergy Asthma Immunol. Res. 2022, 14, 379–392. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Chang, H.-C.; Simon-Buss, A.; Jang, Y.-R.; Denery-Papini, S.; Pineau, F.; Gu, Y.Q.; Huo, N.; Lim, S.-H.; Kang, C.-S.; et al. Towards reducing the immunogenic potential of wheat flour: Omega gliadins encoded by the D genome of hexaploid wheat may also harbor epitopes for the serious food allergy WDEIA. BMC Plant Biol. 2018, 18, 291. [Google Scholar] [CrossRef]
- Huo, N.; Zhang, S.; Zhu, T.; Dong, L.; Wang, Y.; Mohr, T.; Hu, T.; Liu, Z.; Dvorak, J.; Luo, M.-C.; et al. Gene Duplication and Evolution Dynamics in the Homeologous Regions Harboring Multiple Prolamin and Resistance Gene Families in Hexaploid Wheat. Front. Plant Sci. 2018, 9, 673. [Google Scholar] [CrossRef] [Green Version]
- Altenbach, S.B.; Allen, P.V. Transformation of the US bread wheat ‘Butte 86’ and silencing of omega-5 gliadin genes. GM Crop. 2011, 2, 66–73. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Tanaka, C.K.; Seabourn, B.W. Silencing of omega-5 gliadins in transgenic wheat eliminates a major source of environmental variability and improves dough mixing properties of flour. BMC Plant Biol. 2014, 14, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenbach, S.B.; Tanaka, C.K.; Pineau, F.; Lupi, R.; Drouet, M.; Beaudouin, E.; Morisset, M.; Denery-Papini, S. Assessment of the Allergenic Potential of Transgenic Wheat (Triticum aestivum) with Reduced Levels of ω5-Gliadins, the Major Sensitizing Allergen in Wheat-Dependent Exercise-Induced Anaphylaxis. J. Agric. Food Chem. 2015, 63, 9323–9332. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Watanabe, J.; Sonoyama, K.; Tanabe, S. Novel Method for Producing Hypoallergenic Wheat Flour by Enzymatic Fragmentation of the Constituent Allergens and Its Application to Food Processing. Biosci. Biotechnol. Biochem. 2000, 64, 2663–2667. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Tokue, C.; Ogiwara, H.; Kimura, H.; Arai, S. Chemical and Nutritional Properties of Hypoallergenic Wheat Flour. Biosci. Biotechnol. Biochem. 2001, 65, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Narasaka, S.; Endo, Y.; Fu, Z.; Moriyama, M.; Arai, S.; Abe, K.; Kato, H. Safety Evaluation of Hypoallergenic Wheat Flour by Using a DNA Microarray. Biosci. Biotechnol. Biochem. 2006, 70, 1464–1470. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Tanabe, S.; Watanabe, M.; Shinmoto, H.; Sonoyama, K. The production of hypoallergenic wheat flour and the analysis of its allergy suppressive effects. Biofactors 2004, 22, 295–297. [Google Scholar] [CrossRef]
- Tanabe, S. Analysis of Food Allergen Structures and Development of Foods for Allergic Patients. Biosci. Biotechnol. Biochem. 2008, 72, 649–659. [Google Scholar] [CrossRef]
- Kumagai, H.; Suda, A.; Sakurai, H.; Kumagai, H.; Arai, S.; Inomata, N.; Ikezawa, Z. Improvement of Digestibility, Reduction in Allergenicity, and Induction of Oral Tolerance of Wheat Gliadin by Deamidation. Biosci. Biotechnol. Biochem. 2007, 71, 977–985. [Google Scholar] [CrossRef] [Green Version]
- Abe, R.; Shimizu, S.; Yasuda, K.; Sugai, M.; Okada, Y.; Chiba, K.; Akao, M.; Kumagai, H.; Kumagai, H. Evaluation of Reduced Allergenicity of Deamidated Gliadin in a Mouse Model of Wheat-Gliadin Allergy Using an Antibody Prepared by a Peptide Containing Three Epitopes. J. Agric. Food Chem. 2014, 62, 2845–2852. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Adamidi, C.; Lozano, R.M.; Yee, B.C.; Momma, M.; Kobrehel, K.; Ermel, R.; Frick, O.L. Thioredoxin-linked mitigation of allergic responses to wheat. Proc. Natl. Acad. Sci. USA 1997, 94, 5372–5377. [Google Scholar] [CrossRef] [Green Version]
- Yano, H.; Wong, J.H.; Lee, Y.M.; Cho, M.-J.; Buchanan, B.B. A strategy for the identification of proteins targeted by thioredoxin. Proc. Natl. Acad. Sci. USA 2001, 98, 4794–4799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, H. Ongoing Applicative Studies of Plant Thioredoxins. Mol. Plant 2014, 7, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Waga, J.; Zientarski, J.; Obtułowicz, K.; Bilo, B.; Stachowicz, M. Gliadin Immunoreactivity and Dough Rheological Properties of Winter Wheat Genotypes Modified by Thioredoxin. Cereal Chem. 2008, 85, 488–494. [Google Scholar] [CrossRef]
- Matsumoto, T.; Shimada, Y.; Hirai, S. Mitigated binding of IgE to thioredoxin-treated salt-soluble wheat allergens in a child with Baker’s asthma. Ann. Allergy, Asthma Immunol. 2007, 98, 599–600. [Google Scholar] [CrossRef] [PubMed]
- Kasarda, D.D.; Bernardin, J.E.; Qualset, C.O. Relationship of gliadin protein components to chromosomes in hexaploid wheats (Triticum aestivum L.). Proc. Natl. Acad. Sci. USA 1976, 73, 3646–3650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DuPont, F.M.; Vensel, W.H.; Chan, R.; Kasarda, D.D. Characterization of the 1B-Type ω-Gliadins from Triticum aestivum Cultivar Butte. Cereal Chem. 2000, 77, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Wieser, H.; Kieffer, R.; Lelley, T. The influence of 1B/1R chromosome translocation on gluten protein composition and technological properties of bread wheat. J. Sci. Food. Agric. 2000, 80, 1640–1647. [Google Scholar] [CrossRef]
- Corbellini, M.; Empilli, S.; Vaccino, P.; Brandolini, A.; Borghi, B.; Heun, M.; Salamini, F. Einkorn Characterization for Bread and Cookie Production in Relation to Protein Subunit Composition. Cereal Chem. 1999, 76, 727–733. [Google Scholar] [CrossRef]
- Tanabe, S.; Arai, S.; Yanagihara, Y.; Mita, H.; Takahashi, K.; Watanabe, M. A Major Wheat Allergen Has a Gln-Gln-Gln-Pro-Pro Motif Identified as an IgE-Binding Epitope. Biochem. Biophys. Res. Commun. 1996, 219, 290–293. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Conti, A.; Pravettoni, V.; Bonomi, S.; Iametti, S.; Fortunato, D.; Scibilia, J.; Bindslev-Jensen, C.; Ballmer-Weber, B.; et al. Wheat IgE-mediated food allergy in European patients: Alpha-amylase inhibitors, lipid transfer proteins and low-molecular-weight glutenins. Allergenic molecules recognized by double-blind, placebo-controlled food challenge. Int. Arch. Allergy Immunol. 2007, 144, 10–22. [Google Scholar] [CrossRef]
- Du Toit, G.; Sampson, H.A.; Plaut, M.; Burks, A.W.; Akdis, C.A.; Lack, G. Food allergy: Update on prevention and tolerance. J. Allergy Clin. Immunol. 2017, 141, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brough, H.A.; Nadeau, K.C.; Sindher, S.B.; Alkotob, S.; Chan, S.; Bahnson, H.T.; Leung, D.Y.M.; Lack, G. Epicutaneous sensitization in the development of food allergy: What is the evidence and how can this be prevented? Allergy 2020, 75, 2185–2205. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto-Hanada, K.; Kobayashi, T.; Williams, H.C.; Mikami, M.; Saito-Abe, M.; Morita, K.; Natsume, O.; Sato, M.; Iwama, M.; Miyaji, Y.; et al. Early aggressive intervention for infantile atopic dermatitis to prevent development of food allergy: A multicenter, investigator-blinded, randomized, parallel group controlled trial (PACI Study)—Protocol for a randomized controlled trial. Clin. Transl. Allergy 2018, 8, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaji, Y.; Yang, L.; Yamamoto-Hanada, K.; Narita, M.; Saito, H.; Ohya, Y. Earlier aggressive treatment to shorten the duration of eczema in infants resulted in fewer food allergies at 2 years of age. J. Allergy Clin. Immunol. Pract. 2019, 8, 1721–1724.e6. [Google Scholar] [CrossRef] [PubMed]
- Abe, R.; Matsukaze, N.; Kobayashi, H.; Yamaguchi, Y.; Uto-Kondo, H.; Kumagai, H.; Kumagai, H. Allergenicity of Deamidated and/or Peptide-Bond-Hydrolyzed Wheat Gliadin by Transdermal Administration. Foods 2020, 9, 635. [Google Scholar] [CrossRef]
- Sun, N.; Liu, Y.; Liu, K.; Wang, S.; Liu, Q.; Lin, S. Gastrointestinal fate of food allergens and its relationship with allergenicity. Compr. Rev. Food Sci. Food Saf. 2022, 21, 3376–3404. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-C.; Ren, J.-P.; Cho, M.-J.; Zhou, S.-M.; Kim, Y.-B.; Guo, H.-X.; Wong, J.H.; Niu, H.-B.; Kim, H.-K.; Morigasaki, S.; et al. The Level of Expression of Thioredoxin is Linked to Fundamental Properties and Applications of Wheat Seeds. Mol. Plant 2009, 2, 430–441. [Google Scholar] [CrossRef]
- Stawoska, I.; Wesełucha-Birczyńska, A.; Skoczowski, A.; Dziurka, M.; Waga, J. FT-Raman Spectroscopy as a Tool to Study the Secondary Structures of Wheat Gliadin Proteins. Molecules 2021, 26, 5388. [Google Scholar] [CrossRef]
- Weichel, M.; Glaser, A.G.; Ballmer-Weber, B.K.; Schmid-Grendelmeier, P.; Crameri, R. Wheat and maize thioredoxins: A novel cross-reactive cereal allergen family related to baker’s asthma. J. Allergy Clin. Immunol. 2006, 117, 676–681. [Google Scholar] [CrossRef]
- Brant, A. Baker’s asthma. Curr. Opin. Allergy Clin. Immunol. 2007, 7, 152–155. [Google Scholar] [CrossRef]
- Bianco, M.; Ventura, G.; Calvano, C.D.; Losito, I.; Cataldi, T.R. A new paradigm to search for allergenic proteins in novel foods by integrating proteomics analysis and in silico sequence homology prediction: Focus on spirulina and chlorella microalgae. Talanta 2021, 240, 123188. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Kayrala, N.; Trapeau, M.; Aoun, M.; Bordenave, N. The clean label trend: An ineffective heuristic that disserves both consumers and the food industry? Compr. Rev. Food Sci. Food Saf. 2022, 21, 4921–4938. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Chinuki, Y.; Hamada, Y.; Fukutomi, Y.; Sugiyama, A.; Kishikawa, R.; Fukunaga, A.; Oda, Y.; Ugajin, T.; Yokozeki, H.; et al. Genome-wide association study reveals an association between the HLA-DPB1∗02:01:02 allele and wheat-dependent exercise-induced anaphylaxis. Am. J. Hum. Genet. 2021, 108, 1540–1548. [Google Scholar] [CrossRef] [PubMed]


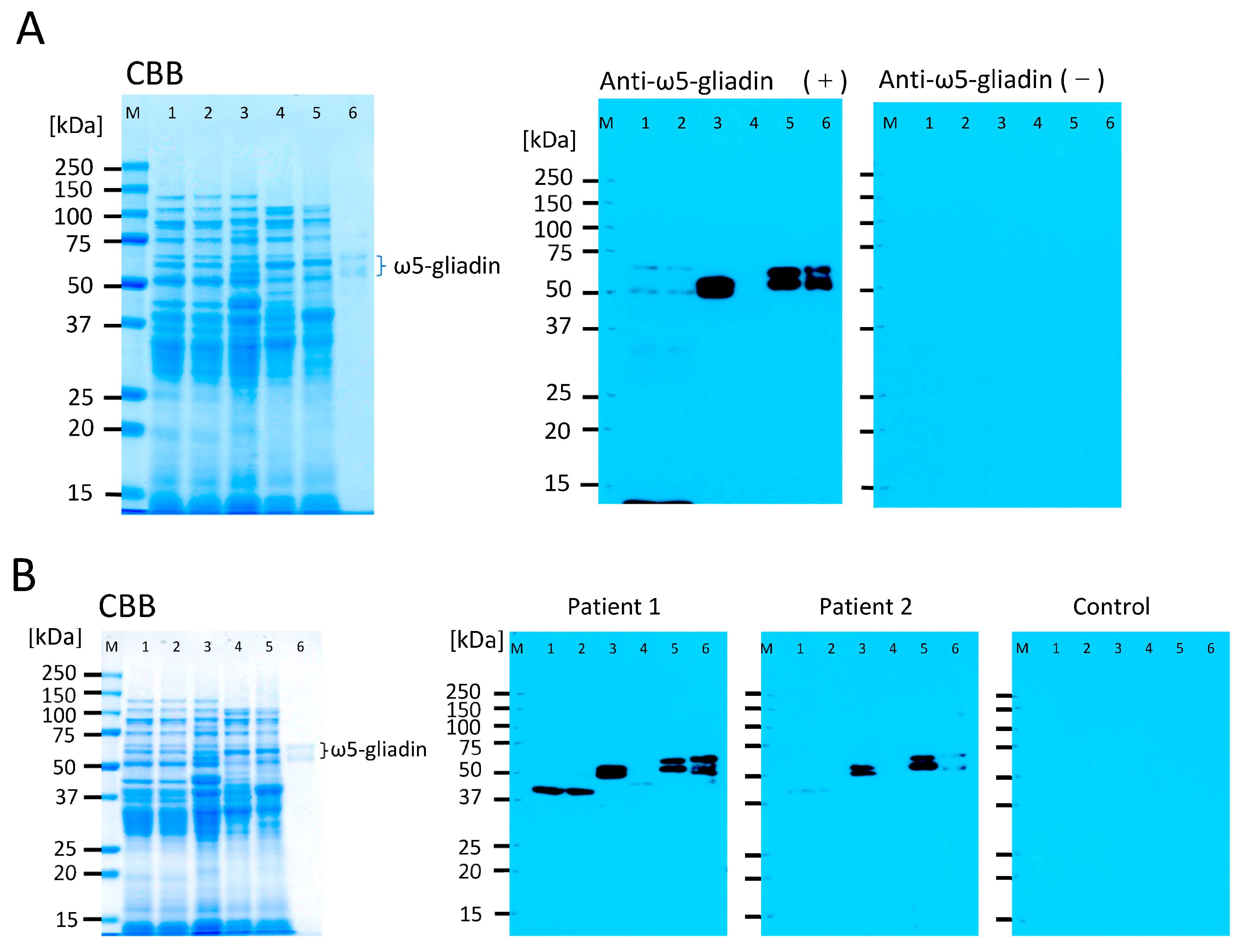{kind=link}
| Allergens | Common Name | MW (kDa) | Route of Allergen Exposure | Ref. No. |
|---|---|---|---|---|
| Tri a 19 | ω5-gliadin | 65 | Food (Ingestion) | [15,16,17,18,19,20,21] |
| Tri a 20 | γ-gliadin | 35–38 | Food (Ingestion), HWP (cutaneous) | [15,22,26] |
| Tri a 21 | α/β-gliadin | 28–35 | Food (Ingestion) | [15,22] |
| Tri a 26 | HMW-glutenin | 67–88 | Food (Ingestion) | [15,20] |
| Tri a 36 | LMW-glutenin | 32–35 | Food (Ingestion) | [15,25] |
| — | ω1,2-gliadin | 40 | Food (Ingestion), HWP (cutaneous) | [15,22,26] |
| — | Peroxidase-1 | 36 | Cross-reactivity to grass pollen allergens | [27] |
| — | β-glucosidase | 60 | Cross-reactivity to grass pollen allergens | [27] |
| Methods | Major Outcomes and Limitations | Ref. No. |
|---|---|---|
| Wheat lines with a reduced allergenicity | ||
| 1BL/1RS translocation (Clement) | Significant reduction of ω5-gliadin protein was presented using SDS-PAGE and immunoblotting with rabbit polyclonal antibody. Low allergenicity was presented with IgE-immunoblotting using small numbers of WDEIA patients’ sera. No oral challenge test was preformed in human. | [36] |
| 1B/1R translocation (Pamier) | Significant reduction of ω5-gliadin protein was determined by RP-HPLC. No significant difference in basophil activation compared with conventional wheat line in WDEIA patients. | [37,38] |
| A genome diploid einkorn lacking B chromosomes | Low allergenicity was presented in negative skin prick test and lack of IgE-immunoreactivity to ω5-gliadin in almost all WDEIA patients sensitized with ω5-gliadin. No oral challenge test was performed in human. | [39] |
| Wheat line lacking all ω-gliadin encoding loci Gli A1, Gli B1 and Gli D1 established using traditional breeding (3xN) | Lack of ω5-gliadin was determined using SDS-PAGE and RP-HPLC. Low reactivity to ω5-gliadin was presented using ELISA with several WDEIA patients’ sera. Significant IgE reactivity was presented to other gluten proteins. No oral challenge test was performed in human. | [40,41] |
| Aneuploid wheat line lacking Gli-B1 locus (1BS-18) | Lack of ω5-gliadin protein was determined using RP-HPLC and immunoblotting with rabbit polyclonal antibody. Low allergenicity was presented using guinea pig and rat wheat challenge models. Oral tolerance to wheat was determined using rat models. No oral challenge test was performed in human. | [42,43,44,45] |
| Wheat line lacking Gli B1 and Glu B3 loci established using double-haploid breeding (DH20) | Low allergenicity was determined using IgE-immunoblotting with 14 WDEIA patients’ sera, immunoblot inhibition assay and ELISA inhibition assay. No oral challenge test was performed in human. | [46,47,48,49] |
| Genetic engineering | ||
| Transgenic wheat lines with RNA interference for ω5-gliadin (Butte 86) | ω5-Gliadin protein was either undetectable or depleted in SDS-PAGE. Greatly reduced IgE-immunoreactivity to ω5-gliadin was presented in most of 11 WDEIA patients using IgE-immunoblotting. Small amount of ω5-gliadin protein derived from 1D chromosome remaind in the line. No oral challenge test was performed in human. | [50,51,52] |
| Enzymic degradation | ||
| Two-step method to produce hypoallergenic wheat using cellulase and actinase | Low allergenicity and immunotolerance effect were confirmed by clinical open studies. Thirteen of 15 children with AD ingested cupcake made of the wheat without allergy symptom. Most children with wheat allergy associated with AD became immunotolerance to normal wheat product after continuous intake of the cupcake. | [53,54,55,56,57] |
| Ion exchanger deamidation | ||
| Deamidation of gliadin using a cation exchange resin | Low allergenicity was presented using IgE-immunoblotting with sera obtained from patients with High Serum IgE, and a mouse model of wheat-gliadin allergy. No clinical study was performed in human. | [58,59] |
| Thioredoxin treatment | ||
| Reduced thioredoxin treatment of wheat allergens | Low allergenicity was presented using ELISA with Serum IgE obtained from patients with wheat allergy, skin prick test and CAP-FEIA inhibition assay. No oral challenge test was performed in human. | [60,61,62,63,64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morita, E.; Matsuo, H.; Kohno, K.; Yokooji, T.; Yano, H.; Endo, T. A Narrative Mini Review on Current Status of Hypoallergenic Wheat Development for IgE-Mediated Wheat Allergy, Wheat-Dependent Exercise-Induced Anaphylaxis. Foods 2023, 12, 954. https://doi.org/10.3390/foods12050954
Morita E, Matsuo H, Kohno K, Yokooji T, Yano H, Endo T. A Narrative Mini Review on Current Status of Hypoallergenic Wheat Development for IgE-Mediated Wheat Allergy, Wheat-Dependent Exercise-Induced Anaphylaxis. Foods. 2023; 12(5):954. https://doi.org/10.3390/foods12050954
Chicago/Turabian StyleMorita, Eishin, Hiroaki Matsuo, Kunie Kohno, Tomoharu Yokooji, Hiroyuki Yano, and Takashi Endo. 2023. "A Narrative Mini Review on Current Status of Hypoallergenic Wheat Development for IgE-Mediated Wheat Allergy, Wheat-Dependent Exercise-Induced Anaphylaxis" Foods 12, no. 5: 954. https://doi.org/10.3390/foods12050954